
Abstract
The efficient differentiation of retinal cells from human pluripotent stem cells remains a major challenge for the development of successful and cost-effective cellular therapies for various forms of blindness. Current differentiation strategies rely on exposing pluripotent stem cells to soluble growth factors that play key roles during early development (such as DKK-1, Noggin, and IGF-1) at 20% oxygen (O2). This O2 tension is, however, considerably higher than O2 levels during organogenesis and may impair the differentiation process. In this study, we examined the effect of mimicking the physiological O2 tension (2%) on the generation of retinal progenitor cells (RPCs) from human induced pluripotent stem cells (iPSCs) and human embryonic stem cells (hESCs). Both cell types were induced to differentiate into RPCs at 20% and 2% O2. After 3 days in suspension culture as embryoid bodies (EBs), 2% O2 caused the activation of hypoxia inducible factor responsive genes VEGF and LDHA and was accompanied by elevated expression levels of the early eye field genes Six3 and Lhx2. Twenty-one days after plating EBs in an adherent culture, we observed more RPCs co-expressing Pax6 and Chx10 at 2% O2. Quantitative polymerase chain reaction analysis confirmed that lowering O2 tension had caused a rise in the expression of both genes compared with 20% O2. Our results indicate that mimicking physiological O2 is a favorable condition for the efficient generation of RPCs from both hiPSCs and hESCs.
Introduction
H
A number of studies have also mimicked O2 tensions present in adult organs during the culture of adult stem cells. Low O2 tension in the brain is conserved among mammalian species. In the rat brain, O2 tension varies at different locations (white cortex 0.8%–2.1%, hypothalamus 1.4%–2.1%, and hippocampus 2.6%) [14] as with human brain (3.2% under and 4.4% above the dura) [15]. The subventricular zone (SVZ) is the main neurogenic niche in the human brain where neural stem cells (NSCs) are responsible for adult neurogenesis [16]. The O2 tension of the human SVZ has never been directly measured; however, it is widely accepted that NSCs reside in areas of relatively low O2 tension. This is further supported by studies showing that human NSCs showed improved survival, proliferation, and maintenance of an undifferentiated phenotype when cultured at 5% O2 tension [17]. Low O2 tensions also exist in other adult stem cell niches such as bone marrow, a source of hematopoietic and mesenchymal stem cells (MSCs) [18]. Indeed, there is evidence that lower O2 tension promotes the maintenance of an undifferentiated state in cultured bone marrow-derived MSCs [19,20]. Taken together, these studies have revealed that mimicking in vivo O2 tensions can be used to enhance the undifferentiated expansion and directed differentiation of both pluripotent and adult stem cells.
In this study, we sought to optimize the differentiation of human pluripotent stem cells into retinal progenitor cells (RPCs) by lowering O2 tension. RPCs are NSCs capable of differentiating into all 6 types of retinal neurons and muller glia cells, and have been generated from human pluripotent stem cells [21 –27]. During early development, Wnt and BMP signaling pathways are antagonized in the development of the forebrain [28,29]. Mimicking this phenomenon using soluble growth factors DKK-1 (Wnt pathway antagonist), Noggin (BMP pathway antagonist), Lefty A (nodal pathway antagonist), and insulin-like growth factors (IGF-1) were successful in generating RPCs from human pluripotent stem cells [21,22]. Lamba et al. used a combination of DKK-1, Noggin, and IGF-1 to induce retinal differentiation, and we used this protocol to examine the effect of lowering O2 tension on early neural retinal differentiation of induced pluripotent stem cells (iPSCs) and hESCs [21]. We previously showed that 2% O2 tension increases neuronal differentiation efficiency of mESCs [12]. Under the same condition, we observed a significant increase in the percentage of RPCs from both iPSC and hESC. In conclusion, we have demonstrated the importance of the O2 during in vitro differentiation and propose that O2 tension is a crucial variable in overcoming low retinal differentiation efficiency.
Materials and Methods
Human pluripotent stem cell culture
Undifferentiated human pluripotent cells (MSUH001 iPSCs and Shef3 hESCs were obtained from the Spanish and the UK Stem Cell Bank, respectively) were grown on Mitomycin-C (1 mg/mL; Sigma-Aldrich) inactivated mouse embryo fibroblasts (MEF) in Knockout DMEM supplemented with 20% (v/v) Knockout serum replacement, 1 mM L-glutamine, 1% (v/v) nonessential amino acids, 100 mM β-mercaptoethanol, and 4 ng/mL basic fibroblast growth factor (all Invitrogen). They were cultured in a Sanyo IncuSafe incubator (Sanyo, MCO-18AIC) at 37°C and 5% (v/v) CO2, and 5 mL of fresh growth medium was exchanged every 24 h. iPSCs were transferred to newly Mitomycin-C inactivated MEF every 3–4 days by dissecting colonies into smaller clumps with Fine Tip Mini Pastette (Alpha Laboratories). hESCs colonies were dissected in the same way after incubating with 0.025 mg/mL collagenase (Invitrogen) for 3 min at 37°C.
Early neural retinal differentiation of human pluripotent stem cells
Human pluripotent stem cells were induced to differentiate by using a previously published protocol [21]. Briefly, undifferentiated pluripotent stem cell colonies were dissected into small clumps with Fine Tip Mini Pastette (Alpha Laboratories) and cultured as embryoid bodies (EBs) in suspension in 30 mm nonadherent bacterial grade culture dishes (Sterilin) containing 3 mL retinal induction medium consisting of DMEM/F12 supplemented with 10% (v/v) Knockout Serum Replacement (both Invitrogen), 1 ng/mL human recombinant DKK-1, 1 ng/mL human recombinant noggin, 5 ng/mL human recombinant IGF-1 (all R&D Systems), and 1% (v/v) N-2 Supplement (PAA Laboratories Ltd) for 3 days (Supplementary Fig. S1D; Supplementary Data are available online at
Oxygen tension control
The hypoxic chambers used in these studies had previously been fabricated to maintain 2% O2 during the neuronal differentiation mESCs (full description of construction and operation in Ref. [14]). Briefly, undifferentiated pluripotent stem cell colonies were dissected into small clumps as just described before placing the cell culture dish into a hypoxic chamber. To maintain humidity within the chamber, a 30 mm petri dish (Sterilin) containing 3 mL of distilled water was also placed in the chamber. The chamber was then gassed from a premixed gas cylinder composed of 2% (v/v) O2, 5% (v/v) CO2, and 93% (v/v) N2 (BOC) for 5 min before placing it in an incubator at 37°C (Supplementary Fig. S1B). After 3 days, the culture dishes were removed from the chamber and plated into 6-well plates coated with Matrigel as just described before being returned to the chamber, gassed, and placed back into the incubator (Supplementary Fig. S1C). The medium was changed every 2 days during which the chamber was re-gassed.
Immunocytochemistry
Cells were fixed with 2 mL 4% (w/v) paraformaldehyde (PFA) for 20 min before being permeabilized with 0.25% (v/v) Triton X-100 in Dulbecco's modified phosphate-buffered saline (PBS) for 10 min at room temperature (all from Sigma-Aldrich). The samples were washed with 2 mL DPBS and incubated in 2 mL blocking solution (2% (v/v) goat serum, 0.25% (v/v) Triton X-100 in PBS, all from Sigma-Aldrich), for 30 min. For the detection of cell surface marker, the permeabilization step was omitted, and cells were incubated straight into the blocking solution for 30 min. The samples were then incubated with 600 μL of primary antibodies diluted in blocking solution overnight at 4°C.
The primary antibodies used were as follows: monoclonal mouse anti-Oct4 IgG (Invitrogen 1:400), monoclonal mouse anti-Tra-1-60 IgM (1:200), polyclonal mouse anti-Nestin IgG (1:400), monoclonal mouse anti-Pax6 IgG (1:300), polyclonal rabbit anti-Otx2 (1:1,000), monoclonal mouse-Neurofilament-M IgG (1:400), polyclonal rabbit anti-S-opsin (1:200), polyclonal rabbit anti-Nrl (1:200, all from Millipore), polyclonal Rabbit anti-Chx10 IgG (Sigma-Aldrich; 1:300), monoclonal mouse anti-rhodopsin (1:200), monoclonal mouse anti-Prox1 (1:200), polyclonal goat anti-IRBP (1:200), polyclonal goat anti-Six3 (1:200), polyclonal goat anti-Lhx2 (1:200, all from Santa Cruz), monoclonal mouse anti-Crx (Abnova), and monoclonal rabbit-anti-PKCα IgG (1:400; Abcam). The cells were then washed twice with 2 mL PBS and incubated with 600 μl of Alexa Flour 488 goat anti-mouse, Alexa Flour 488 goat anti-rat IgM, Alexa Flour 555 goat anti-mouse (all Invitrogen; 1:400), and Cy3 conjugated donkey anti-rabbit IgG (Millipore; 1:300) in blocking solution for 1 h at room temperature (details of primary and secondary antibody combinations can be found in Supplementary Table S1). The same concentration of the corresponding isotype controls were used as a negative control [murine IgM isotype control (Sigma-Aldrich), mouse and rabbit IgG isotype control (both from Abcam)].The cells were washed twice with 2 mL PBS and incubated with 4,6-diamidino-2-phenylindole (DAPI; Invitrogen, 1:1,000) in PBS for 5 min. Fluorescence images were acquired by using a fluorescence microscope (Nikon, Eclipse TE2000-U) and analyzed with NIS-element software. Confocal images were taken with Perkin Elmer Ultraview Spinning Disk confocal microscope.
Detection of pimonidazole hydrochloride at 2% O2
The hypoxia marker pimonidazole hydrochloride (Hypoxyprobe-1 kit, HPI Inc.) was used to demonstrate the exposure of cells to lowered O2 tension in the hypoxic chamber. iPSCs and hESCs were cultured at both 20% and 2% O2 for 2 days. 100 μM pimonidazole was added 2 h before fixation with 4% PFA. The fixed samples were blocked as previously described and incubated with anti-pimonidazole mouse monoclonal antibody IgG1 (HPI Inc., 1:30 in PBS) or mouse IgG isotype control (Abcam) overnight at 4°C. The samples were washed with 1 mL PBS and incubated with 600 μL of Alexa Flour 488 goat anti-mouse IgG (Invitrogen; 1:400) for 2 h at room temperature. The samples were washed with 1 mL PBS and incubated with DAPI (Invitrogen; 1:1,000) for 2 min in the dark before being analyzed by confocal microscopy.
Flow cytometry
Cell surface markers
Samples were washed with 2 mL DPBS and incubated with 2 mL Trypsin (Invitrogen) for 10 min at 37°C. The cultures were quenched with DMEM containing 10% (v/v) FBS (Invitrogen) and physically dissociated by pipetting 10 times with a Pasteur pipette (Fisher Scientific). The dissociated cells were re-suspended in DMEM containing 10% (v/v) FBS and centrifuged for 3 min at 1,200 rpm. The supernatant was removed, and the pellet was re-suspended in 2 mL of PBS. After centrifuging at the same conditions, the supernatant was removed, and the cells were fixed with 0.5 mL 4% (w/v) PFA for 20 min on ice. The cells were centrifuged at 1,200 rpm for 3 min and re-suspended in 0.5 mL blocking solution (2% (v/v) goat serum in DPBS) for 30 min on ice. The cells were again centrifuged at the same condition and incubated with the blocking solution containing monoclonal mouse anti-Tra-1-60 IgM (1:200) and murine IgM isotope control (Sigma-Aldrich, 1:200) overnight at 4°C. The cells were washed twice with 1.5 mL DPBS and incubated in blocking solution containing Alexa Flour 488 goat anti-mouse IgM (Invitrogen) for 2 h at room temperature. The cells were washed twice with 1.5 mL DPBS before incubating with DAPI for 3 min. FACs analysis was then performed by using a flow cytometer (CyAN ADP, 9 color, 3 lazer analyzer; Beckman Coulter). Dead cells and debris were excluded by using forward and side-scatter parameters, and only a single-cell population was selected by using the pulse width parameter. Overall, 99% of samples incubated with the isotype control and a secondary antibody was gated as a negative population. The data were analyzed with Sumit v4.3 software (DAKO).
Intracellular markers
After fixing cells with PFA, the cells were washed with DPBS and incubated with 0.5 mL permeabilizing solution (0.25% Triton in DPBS) on ice for 10 min. After centrifugation, the cells were blocked in 0.5 mL DPBS with 2% (v/v) goat serum, 0.25% (v/v) triton for 30 min on ice before incubating overnight at 4°C with blocking solution containing murine monoclonal IgM isotope control (1:200), monoclonal mouse anti-Oct4 IgG (1:400), monoclonal mouse anti-Pax6 IgG (1:300), and polyclonal Rabbit anti-Chx10 IgG (1:300). The cells were subjected to 3 DPBS washes before incubating with blocking solution containing Alexa Flour 488 goat anti-mouse IgG and Alexa Flour 647 goat anti-rabbit IgG (Invitrogen) for 2 h at room temperature. Isotype controls were used (same concentration as the primary antibodies) as a negative control [mouse and rabbit IgG isotype control (both from Abcam)]. The cells were washed, counter-stained with DAPI, and analyzed as just described.
RNA extraction and cDNA synthesis
Total RNA extraction was achieved by using the Qiagen RNeasy kit (Qiagen). Briefly, cell pellets were harvested by using a cell scraper (TPP Techno Plastic Products AG), re-suspended, and homogenized by aspirating 5 times through an RNase-free syringe with an 18G needle in buffer RLT provided in the kit. The samples were then processed in accordance with the manufacturer's instructions. In the final step, RNA was eluted with 40 μL of RNase-free water by centrifuging for 1 min at 10,000 rpm. The RNA concentration was determined by measuring absorbance at 260 nm using a spectrophotometer (NanoDrop ND-1000; Thermo Scientific). For cDNA synthesis, we used the Ambion 1st strand cDNA synthesis kit (RETROscript®; Ambion) according to manufacturer's instructions with 1 μg of RNA for each reaction in a total reaction volume of 20 μL.
Real-time quantitative polymerase chain reaction
Quantitative polymerase chain reactions (qPCRs )were performed by using the MESA BLUE qPCR MasterMix Plus for SYBR® Assay (Eurogentec) following the manufacturer's instructions. All primer pairs were acquired from Qiagen, which are listed in Supplementary Table S2. Each reaction contained 1 μL of 1st strand cDNA in a total reaction volume of 20 μL, and all measurements were taken in triplicate. The PCR conditions used were as follows; 2 min at 50°C, 10 min at 95°C, followed by 40 cycles at 95°C for 15 s, 55°C for 30 s, and 60°C for 30 s. The final 3 steps were 95°C for 15 s, 60°C for 15 s, and 95°C for 15 s. All samples were normalized to levels of β-actin (an endogenous housekeeping gene), and relative quantification was carried out as previously described [30].
Statistical analysis
Triplicate samples were analyzed in all experiments. The values are expressed as means±standard error of the mean. To determine statistical significance, paired Student's t-tests were performed, and P≤0.05 was considered significant.
Results
Detection of hypoxia in cells exposed to 2% O2
Pimonidazole hydrochloride is a widely used hypoxic marker for detection of hypoxia both in vitro and in vivo due to its chemical stability and solubility in water [31]. In the cells exposed to low O2 tension, pimonidazole hydrochloride forms adduct with thiol groups in proteins whose immunoreactive side chain can be immunochemically analyzed by using a monoclonal antibody raised against the adducts [32]. Two human pluripotent stem cell lines were cultured at 2% O2 for 2 days in the hypoxic chamber stained positively for hypoxyprobe-1, whereas those cultured at 20% O2 were negative (Supplementary Fig. S2). This provides supporting evidence that cells cultured in our chambers at 2% O2 [12] were exposed to hypoxic conditions.
Activation of hypoxia inducible factor responsive genes and retinal specification in EBs derived from iPSCs (iPSCs-EBs)
Clinical application of ESCs-based cell therapy is often perceived as unethical due to the use of embryos and has drawbacks such as immune rejection. iPSC-based cell therapies would circumvent the use of embryos and may minimize the risk of immune rejection through treating patients with their own cells [33]. Therefore, we examined whether lowering O2 tension had a beneficial impact on the early neural retinal differentiation of iPSCs. For these experiments, we used MSUH001, which had previously been generated in Jose Cibelli's lab (Michigan State University) by expressing a combination of 4 factors in human somatic fibroblasts [34]. These cells morphologically resembled hESCs (Fig. 1A and Supplementary Fig. S5A), and immunocytochemistry revealed that they express pluripotency markers Oct4 and Tra-1-60 (Fig. 1C). Flow cytometry showed tha a high percentage of the population expressed Oct4 (92%±1.56%) and Tra-1-60 (84%±2.05%) (Fig. 1D, E).

Characterization of MSUH001-iPSCs.
EB formation is a widely used technique in the initial stage of pluripotent stem cell differentiation [1]. iPSCs colonies were dissected into small clumps and cultured in suspension for 3 days to form EBs at 20% and 2% O2 in the presence of eye field inducing factors (Supplementary Fig. S1B, D). The expression level of pluripotency genes in EBs formed under both O2 tensions was comparable to that of pluripotent iPSCs (Fig. 2A). Although qPCR analysis of Oct4 and Nanog revealed that their expression was lower at 2% compared with 20% O2, these results were not significant. Immunocytochemistry of D3 EBs revealed that many cells formed in both conditions remained Oct4-positive (Fig. 2B), thus indicting that 2% O2 did not affect the down-regulation of pluripotency markers in EBs differentiated in the presence of DKK-1, Noggin, and IGF-1.
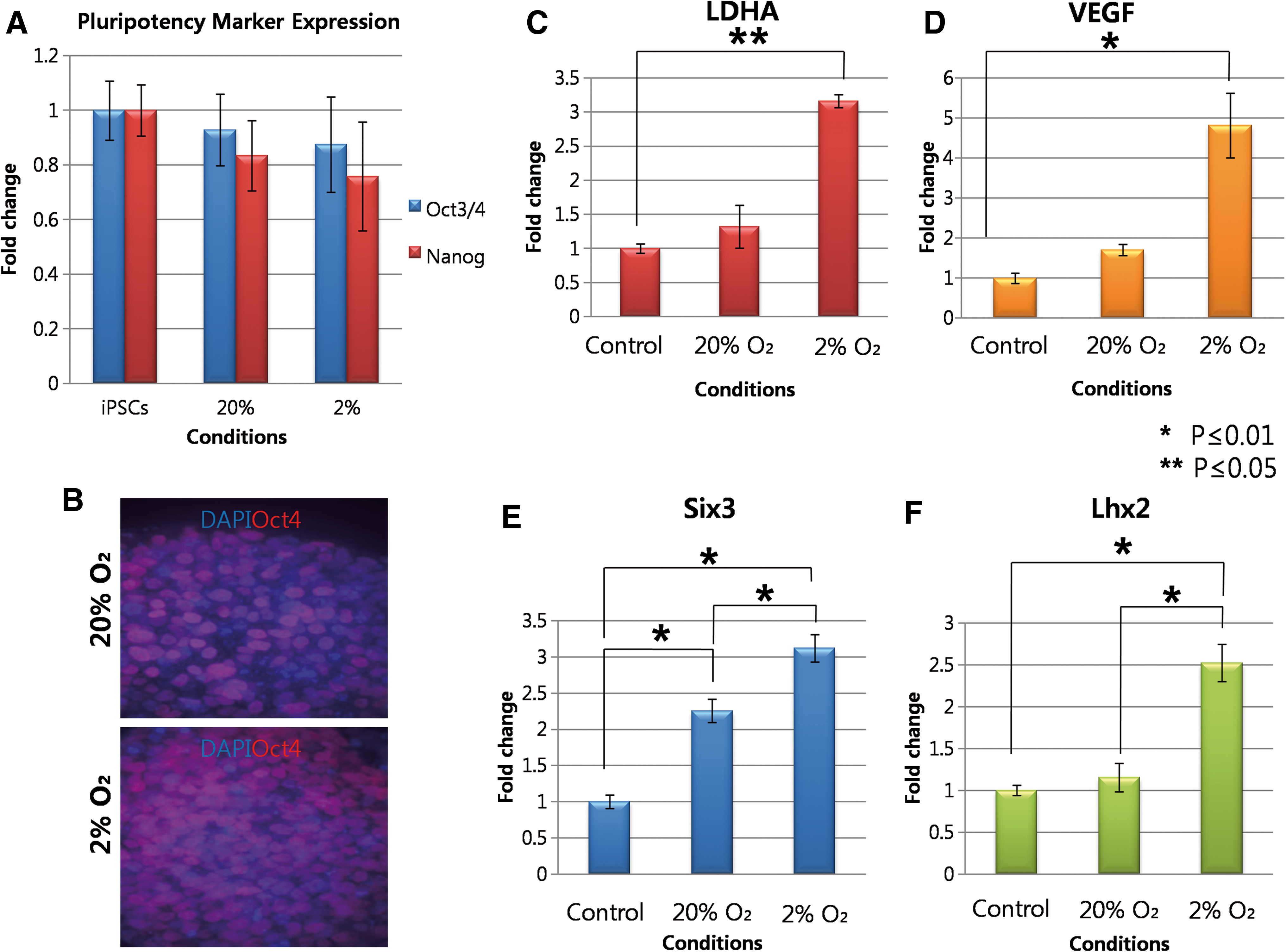
Expression of pluripotency, early eye field, and HIF responsive genes in D3 iPSCs-EBs cultured in both O2 conditions.
The exposure of human cells to O2 tensions below 21% O2 leads to the stabilization of hypoxia inducible factor (HIF)-1a and the activation of over 250 downstream genes including vascular endothelial growth factor (VEGF) and lactate dehydrogenase (LDHA) [35]. We next investigated whether 2% O2 had led to the expression of HIF-1a responsive genes VEGF and LDHA in EBs exposed to 2% O2 for 3 days. qPCR analysis showed a significant 3.2-fold increase in VEGF expression and a 4.8-fold increase in LDHA expression in EBs formed at 2% O2 compared with 20% O2 (P≤0.05 in both cases, Fig. 2C, D). The results confirm that EBs cultured at 2% O2 for 3 days in the presence DKK-1, Noggin, and IGF led to the up-regulation of HIF-1a responsive genes, most probably through O2 dependent stabilization of HIF-1a.
Next, to determine whether lowered O2 tension acts in synergy with the growth factors to induce the retinal specification of pluripotent cells, we performed qPCR on D3 EBs to determine the expression level of Six3 and Lhx2, which are 2 genes required for early eye field formation [36,37]. The analysis revealed that there was a 2.2-fold increase (P≤0.05) in Six3 expression when EBs were differentiated at 20% O2 in the presence of DKK-1, Noggin, and IGF-1 compared with spontaneously differentiating EBs controls at 20% O2 in the absence of any growth factors (Fig. 2E). However, we observed a bigger margin of increase in the expression level of Six3 when EBs differentiated with all growth factors at 2% O2 compared with spontaneously differentiating EBs at 20% O2 (3.1-fold increase, P≤0.05, Fig. 2E). We observed a similar phenomenon with Lhx2 expression level. The expression level of Lhx2 in EBs differentiated with growth factors at 20% O2 was 1.2-fold higher than that in the spontaneously differentiating EBs (not significant, P>0.05, Fig. 2F). However, Lhx2 expression level significantly increased when EBs differentiated in the presence of all growth factors at 2% O2 compared with spontaneously differentiating EBs at 20% O2 (2.5-fold increase, P≤0.05, Fig. 2F). At the same time point, the expression level of Otx2 was also compared between the conditions; however, no significant differences were observed between the conditions (Supplementary Fig. S3A).
Effect of lowered O2 tension on generation of RPCs derived from iPSCs (iPSCs-RPCs)
Two days after plating EBs, cells migrated as tightly packed neuroepithelial cells from the site at which the EBs had attached under both conditions in a similar pattern as previously observed (Fig. 3) [26]. Eight days after plating EBs, densely packed populations of cells arranged in a radial manner, termed as neural rosettes, due to their resemblance to early neural tube development [38] formed in close proximity to the original site of attachment at 2% O2. At 20% O2, neural rosettes did not form until 10 days after plating. The morphology of cells under both conditions became multilayered and indistinguishable 15 days after plating EBs.
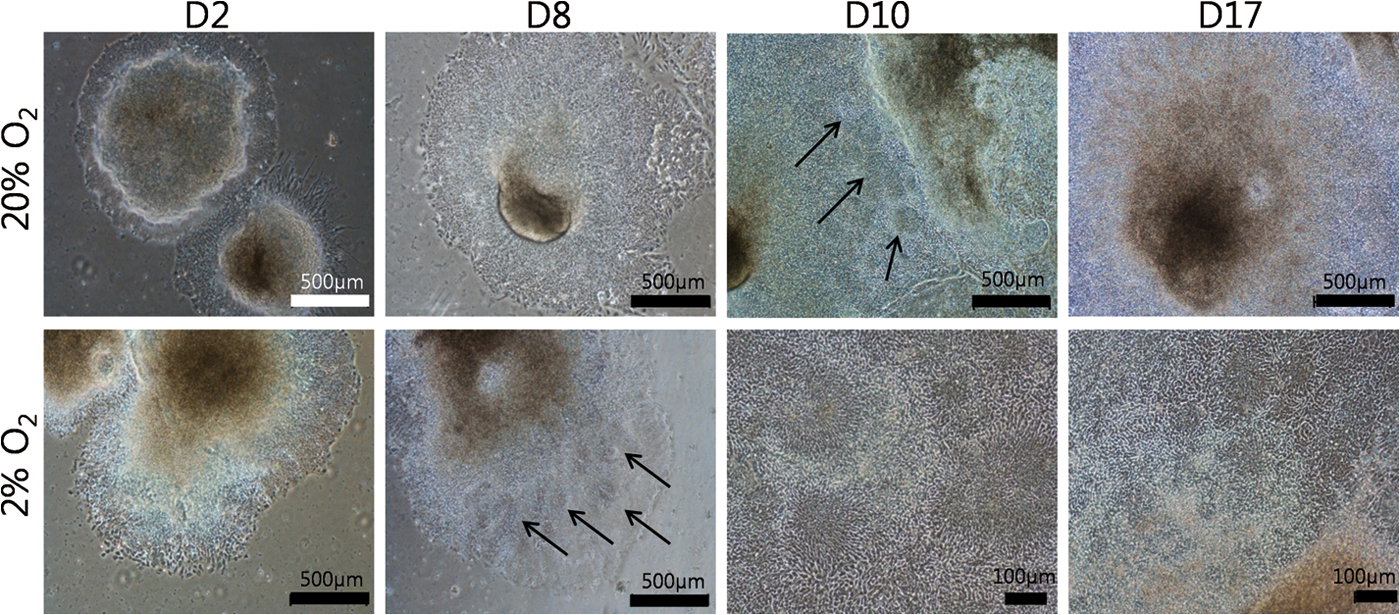
Morphological analysis retinal differentiation of iPSCs at both O2 tensions. Representative images were taken on 2, 8, 10, and 17 days after plating iPSCs-EBs. Arrows indicate neural rosette formation. Color images available online at
Pax6 and Chx10 are key transcription factors required for RPC multipotency and proliferation [39,40], and it has been shown that cells co-expressing these markers can be generated from human pluripotent stem cells 21 days after plating EBs in the presence of DKK-1, Noggin, and IGF-1 [21]. Using immunocytochemistry, we found that cells co-expressing RPC markers Pax6 and Chx10 were visually more abundant at 2% O2 relative to 20% O2 21 days after plating EBs (Fig. 4A, B). At 20% O2, the areas of Pax6 positive cells were abundant throughout the culture, but only a few colonies co-expressed Chx10 (Fig. 4A). Pax6 positive cells were also abundant in the culture at 2% O2 (Fig. 4B), but more colonies co-expressing Pax6 and Chx10 were observed at 2% O2 as compared with 20% O2 (Fig. 4B). At the same time point, cells expressing other retinal markers such as Otx2, Lhx2, and Six3 were also observed by immunocytochemistry, but no visible differences were found between the 2 conditions (Fig. 5).

Characterization of iPSCs-RPCs 21 days after plating EBs.
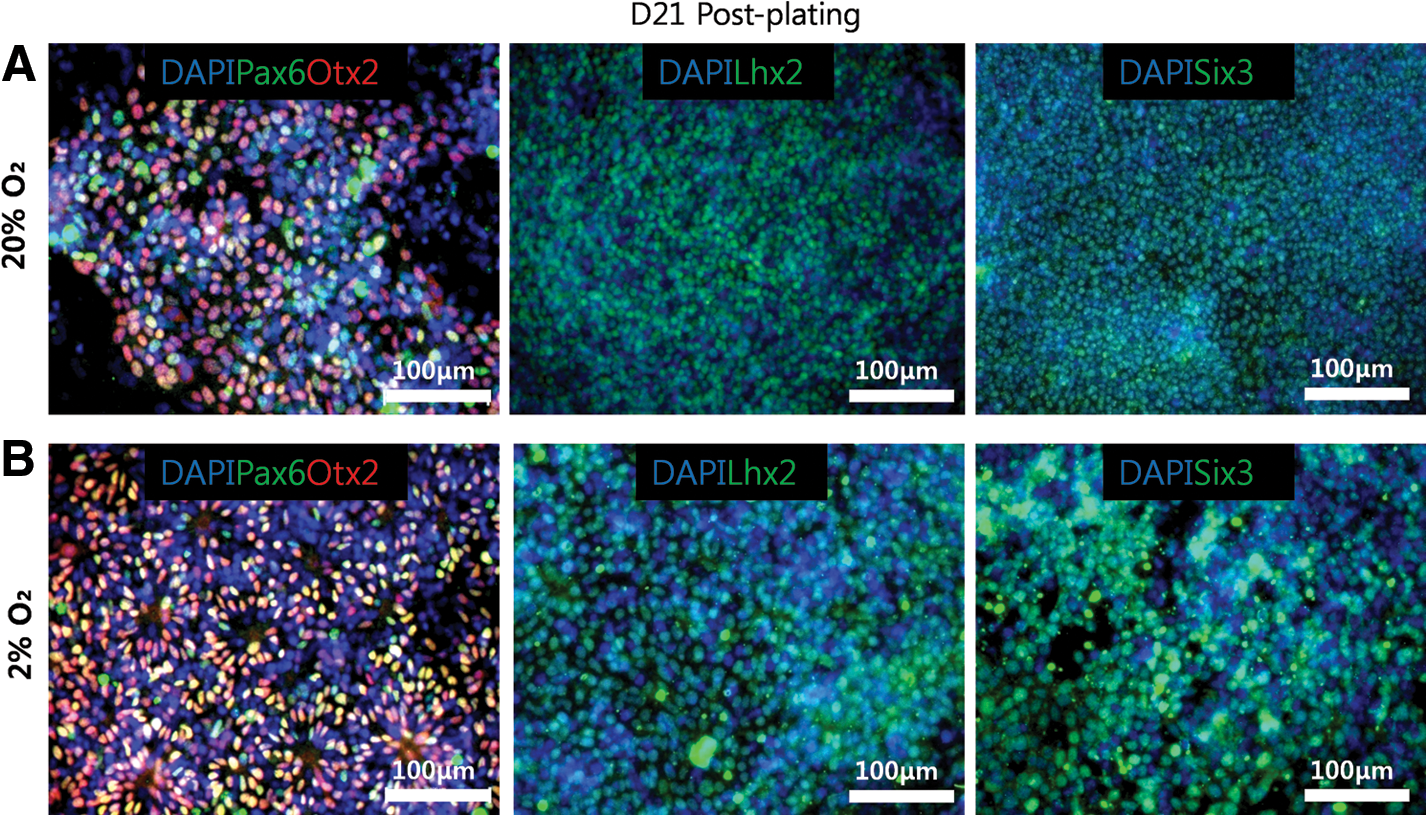
Immunocytochemistry for other retinal markers.
Quantification of iPSCs-RPCs using flow cytometry revealed that at 20% O2, 74%±4.6% were Pax6 positive, and 31%±2.8% expressed Chx10 (Fig. 6A). The majority of Chx10 positive cells also co-expressed Pax6 (30.9%±0.2%) 21 days after plating EBs, representing a true population of RPCs (Supplementary Fig. S4B, C). At 2% O2, there was a slight increase in the percentage of cells expressing Pax6 (80%±3.8%), although this rise was not found to be significant (P>0.05). Culture at 2% O2 was associated with significant rises in the percentage of cells expressing Chx10 (55%±3.8%), which co-expressed Pax6 (54%±1.3%, both P≤0.05) (Fig. 6B and Supplementary Fig. S4A). These results confirm our earlier observation using immunocytochemistry and quantify the improvements in RPC formation that can be achieved at 2% O2.
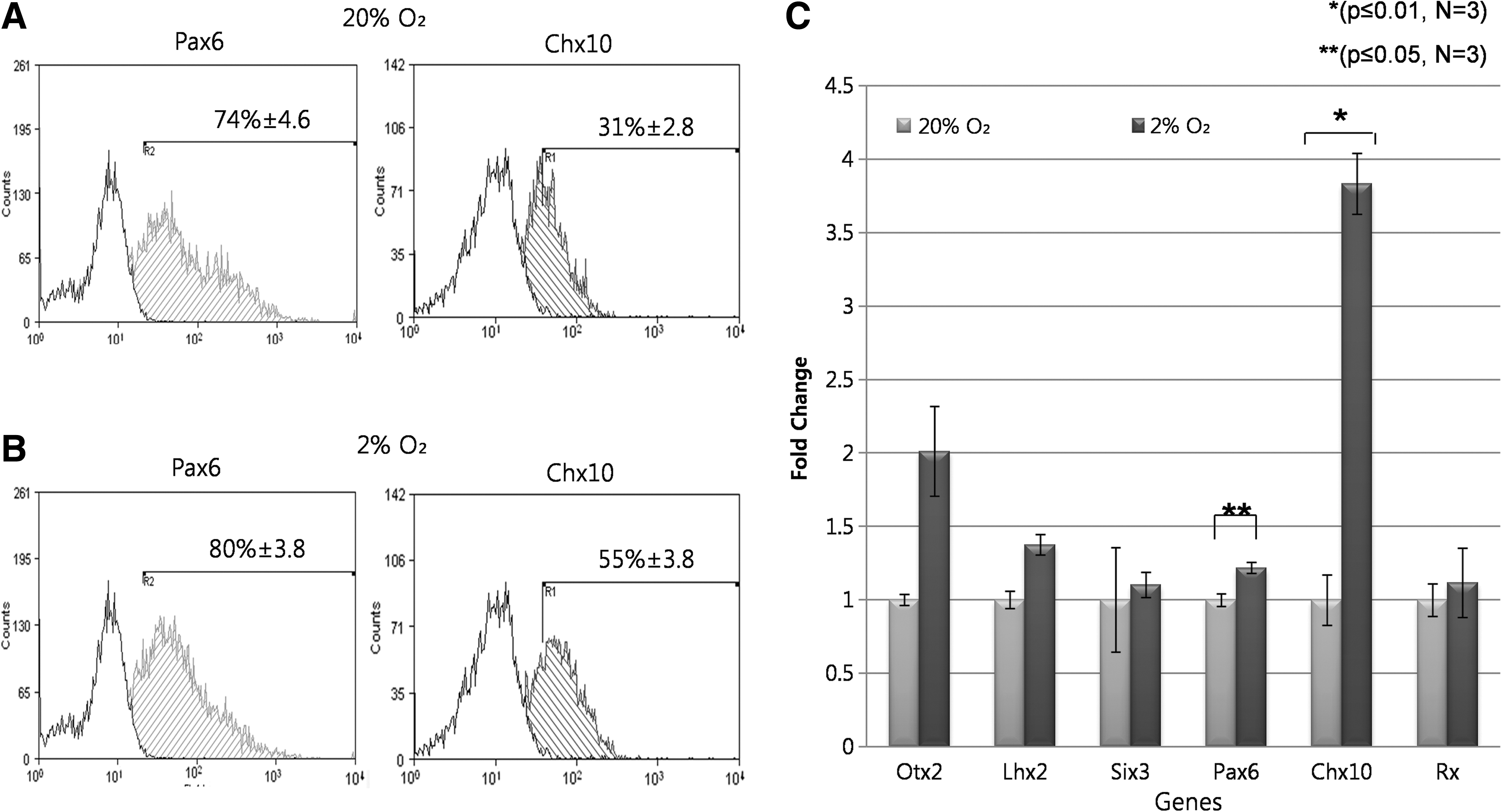
Flow cytometry and qPCR analysis of retinal differentiation cultures at 20% and 2% O2.
Twenty-one days after plating EBs, our qPCR analysis revealed no significant increases in the expression levels of early eye field markers such as Lhx2, Otx2, Six3, and Rx at 2% versus 20% O2 (P>0.05 in all cases). However, a significant 1.2- and 3.8- fold increase was observed in the expression level of Pax6 and Chx10 at 2% O2, respectively, relative to 20% O2 (Fig. 6C). This does not, however, indicate that at 2% O2 culture, many Chx10 positive cells are not co-expressing Pax6. As seen earlier with flow cytometry data (Fig. 6B, C), there is a small difference in the expression level of Pax6 between the 2 conditions and a much larger increase in the expression of Chx10. As we revealed, all the Chx10-positive cells co-expressed Pax6 (Supplementary Fig. S4B, C); so, we can now confidently say that an increase in the expression level of Chx10 corresponds to Pax6/Chx10 co-expressing RPC populations. This was further supported when we detected no immunoactivities to an antibody raised against PKCα 21 days after plating EBs in both conditions, as Chx10 is also a marker for mature bipolar cells (Supplementary Fig. S4D). However, at the same time point, we showed the expression of Neurofilament-M (a marker for ganglion cells) and Prox-1 (a marker for horizontal cells) in both conditions (Fig. 7B). This coincides well with the early retinal development, as RPCs sequentially differentiate into ganglion, cones, and horizontal cells first; then, rods and amacrine cells followed by muller glial and bipolar cells (Fig. 7A). We failed to detect that PKCα and markers for mature photoreceptors (S-opsin and Rhodopsin) may signify that retinal differentiation was prematurely terminated 21 days after plating EBs. However, we detected a small number of cells co-expressing Crx (a cone precursor maker) and IRBP (interphotoreceptor retinoid binding protein) 21 days after plating EBs at 2% O2 (Fig. 7B). These cells were, however, not found in 20% O2 culture, therefore these results suggest that 2% O2 may favour the development of care photoreceptors. The timing of the appearance of Crx/IRBP positive cells also well correlates with the early retinal development, as cones are the first retinal neuron to be generated along with ganglion and horizontal cells from RPCs. The inability of lowered O2 to significantly increase the expression level of these late retinal markers suggests that lowered O2 selectively acts on the early stages of retinal differentiation.

Multilineage differentiation of iPSCs-RPCs at 20% and 2% O2.
Generation of RPCs from hESCs (hESCs-RPCs) under 20% and 2% O2
hESCs colonies were cultured in the presence of MEF feeder layer and formed typical round, tightly packed stem cell colonies (Supplementary Fig. S5A). The colonies also expressed a high level of pluripotency markers Oct4 and Tra-1-60 as measured by imunocytochemistry and FACs (Supplementary Fig. S5C, D). hESCs colonies were removed from the feeder layer and cultured in suspension in the presence of the same growth factors for 3 days in both O2 tensions (Supplementary Fig. S5E). In line with iPSCs-EBs, we observed a significant 2.1-fold increase (P≤0.05) in the expression level of HIF-1a target gene VEGF in EBs cultured at 2% compared with 20% O2 (Supplementary Fig. S5F).
Three days after plating hESCs-EBs at 2% O2, the cultures contained spindle-like cells that formed extensive neurite networks on the periphery of the attached EBs (Supplementary Fig. S6). Although the same spindle cells were visible at 20% O2, there appeared to be fewer neurite processes (Supplementary Fig. S6). Neural rosettes were formed 8 days after plating EBs at 2% O2, which did not appear until 12 days after plating EBs at 20% O2 (Supplementary Fig. S6, arrows). In fact, 15 days after plating EBs, the morphology became increasingly difficult to define by using phase contrast microscopy, as cells became multilayered in both O2 conditions.
To examine the effect of lowered O2 tension on early neural retinal differentiation of hESCs, immunocytochemistry was performed 21 days after plating EBs. At 20% O2, there were large areas of Pax6-positive cells, and only a small population of these cells co-expressed Chx10 confining to densely populated clusters of cells in close proximity to where EBs had attached (Fig. 8A, B). At 2% O2, Pax6 positive cells were also abundant, but noticeably more of these cells co-expressed Chx10 (Fig. 8C–F).
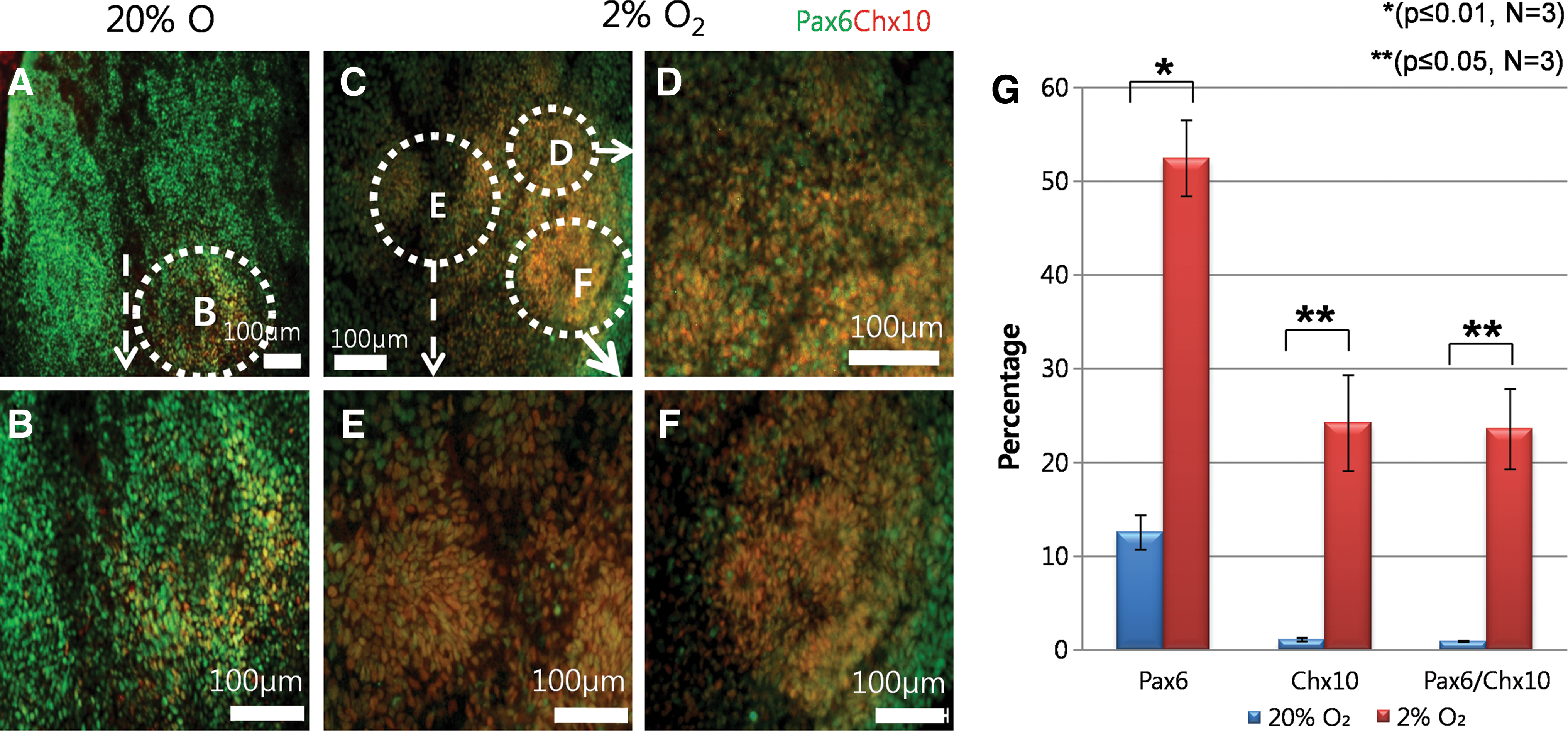
Immunocytochemical and flow cytometry analysis of hESCs-RPCs 21 days after plating EBs.
To quantify Pax6 and Chx10 co-expressing RPCs at both conditions, we used flow cytometry. This analysis revealed that 12%±1.8% of cells differentiated at 20% O2 were Pax6-positive, of which 1%±0.2% co-expressed Chx10 (Fig. 8G and Supplementary Fig. S7A). At 2% O2, significantly more cells expressed Pax6 (52±5.1%, P≤0.05) (Supplementary Fig. S7B) and co-expressing Pax6 and Chx10 (24%±5.1%, P≤0.05) when compared with 20% O2 controls (Fig. 8G). These results confirm an earlier observation using immunocytochemistry and quantify the improvements in RPC formation that can be achieved at 2% O2.
Expression levels of early eye field and RPC markers were observed 14 days after plating hESCs-EBs; the expression level of all the genes increased at 2% O2 in comparison to 20% O2, but only Pax6 and Chx10 were statistically significant (P≤0.01 and P≤0.05 respectively, Supplementary Fig. S7C), thus indicating that lowered O2 tension enhanced transcription of RPC markers.
Comparison between early neural retinal differentiation of iPSCs and hESCs
We observed morphological differences when both pluripotent cell types were induced to differentiate into Pax6 and Chx10 co-expressing RPCs. When hESCs-EBs were plated at both conditions, spindle cells with neurite processes developed on the peripheral region of the attached EBs (Supplementary Fig. S6). However, the development of this neuronal morphology was not observed when iPSCs-EBs were attached. Instead, cells migrated outward from the attached site as tightly packed neuroepithelial colonies similar to those observed in a previous study (Fig. 3) [26]. Flow cytometry revealed that at 20% O2, retinal differentiation of Shef3-hESCs was inefficient, generating just more than 1% of RPCs co-expressing Pax6 and Chx10 (Fig. 8G and Supplementary Fig. S7A). In contrast, iPSCs generated more colonies co-expressing Pax6 and Chx10 (31%±2.1%) at the same condition (Fig. 6A). Despite the differences in the efficiency, the effect of lowering O2 tension in generating RPCs was similar in both pluripotent cell types (Fig. 6B and Supplementary Fig. S7B). qPCR analysis revealed that 2% O2 caused an up-regulation of Pax6 and Chx10 mRNA expression level 14 days after plating iPSCs-EBs compared with 20% O2 culture (Fig. 6C). However, the up-regulation in the genes expressions was observed 21 days after plating hESCs-EBs (Supplementary Fig. S7C). These results indicate that lowering O2 tension enhances the differentiation of both cell types into RPCs, although there are subtle differences between MSUH001 iPSCs and Shef3 hESCs.
Discussion
In ocular genetic disorders, such as retinitis pigmentosa, degeneration of photoreceptors leads to many forms of visual impairment from progressive vision loss to, in most severe cases, complete blindness [41]. To date, these conditions remain untreatable; however, efforts have been made to restore vision by means of transplanting postmitotic photoreceptor precursors [25,27,42]. Human pluripotent stem cells are an ideal source for generating transplantable retinal cells, as they are capable of producing almost all types of cells in the body [1]. Using combinations of forebrain inducing growth factors such as DKK-1 (wnt pathway antagonist) and Noggin (BMP pathway antagonist) as well as a nodal pathway antagonist Lefty A (shown by Takahashi group [22 –24]), it has been possible to generate RPCs from pluripotent stem cells with varying efficiencies [21,22,26]. In particular, Lamba et al. generated ∼80% RPCs co-expressing RPC markers Pax6 and Chx10 21 days after plating hESCs-EBs [21]. In the current study, we have shown that the efficiency of this RPC differentiation protocol is cell line dependent and can be enhanced by lowering O2 tension.
Organogenesis in mammalian embryos takes place in a low O2 environment (∼2%–4% O2) [4], and we have shown that mimicking this O2 tension has a significant impact in inducing key retinal genes during differentiation. Lhx2 and Six3 are early eye field genes required for retinal specification and optic cup development, respectively [36,37]. It has been reported that Lhx2 defines presumptive retina field and transactivates Six3 expression in the same domain [36]. We have observed a nonsignificant increase in the expression level of Lhx2 in EBs differentiating at 20% O2 in the presence of DKK-1, Noggin, and IGF-1 compared with spontaneously differentiating EBs 20% O2 (P>0.05 Fig. 2F). However, the expression level significantly increased at 2% O2 when EBs differentiated in the presence of the growth factors compared with spontaneously differentiating EBs at 20% O2 (Fig. 2F). The increase in the expression level of Lhx2 coincided with an increase in Six3 expression at 2% compared with 20% O2 in the presence of the growth factors (Fig. 2E), thus indicating that lowering O2 to 2% acts synergistically with DKK-1, Noggin, and IGF-1 to enhance the early retinal specification of iPSCs. To further support this, we differentiated iPSCs in the absence of the growth factors in the base media (DMEM supplemented with 10% KOSR) at 2% O2 to observe whether 2% O2 alone is capable of inducing retinal differentiation. Twenty-one days after plating EBs, qPCR analysis revealed no significant changes in the expression level of the genes in the base media at 2% O2 compared with that at 20% O2 (Supplementary Fig. S3B). This suggests that the lowered O2 tension alone was unable to induce retinal differentiation, and none of the retinal genes were detected by immnuocytochemistry (data not shown).
When iPSCs- and hESCs-EBs were plated for 21 days in both O2 tensions, we observed the formation of neural rosette at an earlier time point at 2% O2 compared with 20% O2 (Fig. 3 and Supplementary Fig. S6). This suggests that 20% O2 may delay the development of neural lineages cells from human pluripotent stem cells. In addition, there was a significant increase in the population of cells co-expressing Pax6 and Chx10 under 2% O2 compared with 20% O2 21 days after plating iPSCs- and hESCs-EBs (Figs. 6A, B and 8A–F). The variation in the differentiation efficiency is unclear, although this was also observed in an earlier retinal differentiation study using 2 different hESCs lines [22]. This may indicate that heterogeneous ESCs contain a subpopulation of cells that are refractory to retinal differentiation. We have hypothesized that differences between Shef3 and MSUH001 may be due to an amount of refractory cells present in each cell type. In addition, it is possible that the reprogramming process may enhance the retinal differentiation of iPSCs. Although the percentage of hESCs-RPCs was lower in comparison to iPSCs-RPCs, lowering O2 tension increased the efficiency of RPC formation from both pluripotent cells types.
Increasing differentiation efficiency of pluripotent stem cells in various lineages of cells under low O2 (∼2%–5%) have been reported [9,11,12]. In the case of neuronal differentiation, the efficiency was analyzed by looking at pan neuronal markers such as BIII tubulin and MAP2 [11,12]. Here, we have shown that lowered O2 tension could also be used to enhance generation of a specific population of progenitor cells in the central nervous system (CNS), which are capable of producing all 6 types of retinal neurons and glia cells [43].
The exact mechanism of hypoxia on retinal differentiation requires further investigation, but it has been reported that hypoxic condition is critical in CNS development [44]. HIF pathway is activated by lower O2 tension, and studies of HIF knockout mice revealed embryonic lethality at E10.5 accompanied with neural tube defect and cell death within the cephalic mesenchyme [45,46]. VEGF is a downstream target of HIF pathway [47,48]. Although it is a well-known growth factor for the formation and maintenance of vascular structure [49,50], it also participates in neurogenesis via VEGFR2/Flk-1 receptor [51] and plays a significant role in developing the normal retina [52,53]. VEGFRS/Flk-1 is expressed in neural retina during mouse embryogenesis, and it has been reported that VEGF mediates mouse RPCs proliferation in vitro through VEGFR [54]. Since low O2 activates key signal pathway in vivo during early development, our results indicate that lowering O2 tension during in vitro differentiation may activate key signaling pathways required for CNS development.
Increasing the efficiency of RPC generation from human pluripotent stem cells has a significant advantage for the therapeutic application of stem cell therapy for genetic ocular disorders. It has been reported that RPCs generated from human pluripotent stem cells are common progenitor cells for Crx and Nrl expressing cells that are capable of forming functional photoreceptors on transplantation [25,27,42] However, the efficiency of producing these cells is very low and has not been optimized, which hinders clinical application for treating blindness [25,27]. Our results demonstrate that lowering O2 tension enhanced the generation of RPCs, which can be used as a source material to produce larger numbers of clinically viable postmitotic precursors. Unfortunately, staining for transcription factors such as Pax6 and Chx10 requires a permeabilization step that kills all the cells. Therefore, it is currently not possible to sort and reculture Pax6/Chx10 co-expressing RPCs to generate a higher number of photoreceptors.
Low O2 can be used in synergy with DKK-1, Noggin, and IGF-1 to enhance RPCs production 21 days after plating EBs. However, our results suggested that a prolonged culture may be required to generate postmitotic or mature photoreceptors. We have observed differentiation of ganglion and horizontal cells at the same time point in both conditions but failed to detect photoreceptors (Fig. 5). Despite this, we demonstrated how lowered O2 could favor development of photoreceptors as a small population of Crx/IRBP expressing cells at 2% O2, which was absent in 20% O2 culture (Fig. 7B). It would be interesting to test whether the formation of these cells from RPCs using soluble factors such as retinoic acid (RA) and taurine [22] can be enhanced by lowering O2 tension.
In conclusion, we have shown that differentiating human pluripotent cells under the atmospheric O2 tension is a suboptimal condition for RPCs formation and that the differentiation can be enhanced by lowering the O2 tension near the physiological level. The neural inducing protocol used in this particular system appears to act synergistically with the hypoxic regime employed to induce a significant increase in RPCs differentiation efficiency. This suggests that, in addition to the identified factors, microenvironmetal O2 also plays a pivotal role in generating RPCs from human pluripotent stem cells.
Footnotes
Acknowledgments
The authors would like to thank Professor Jose B. Cibelli (Michigan State University) for the permission for the MSUH001 iPSCs cell line for this research and the Spanish Stem Cell Bank for providing the cell line. D.B would like to thank the Department of Biochemical Engineering of University College London (UCL) for a studentship. P.M.T would like to thank the Mexican Science and Technology Agency (CONACyT) and UCL for the award of a doctoral fellowship. D.H is in receipt of a Technology Strategy Board grant.
Author Disclosure Statement
The authors disclose no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.