
Abstract
Neural precursor cells (NPCs) with high proliferative potential are commonly expanded in vitro as neurospheres. As a population, neurosphere cells show long-term self-renewal capacity and multipotentiality in vitro. These features have led to the assumption that neurosphere cells represent an expansion of the endogenous NPCs residing within the embryonic and adult brain. If this is the case, in principle, bona-fide expansion of endogenous NPCs should not significantly affect their capacity to respond to their original niche of differentiation. To address this issue, we generated primary neurospheres from the dopaminergic niche of the ventral mesencephalon and then transplanted these cells to their original niche within mesencephalic explant cultures. Primary neurosphere cells showed poor capacity to generate dopaminergic neurons in the mesencephalic niche of dopaminergic neurogenesis. Instead, most primary neurosphere cells showed glial commitment as they differentiated into astrocytes in an exclusively neurogenic niche. Subculture of primary cells demonstrated that the neurosphere assay does not amplify niche-responsive dopaminergic progenitors. Further, neurospheres cells were largely unable to acquire the endogenous positional identity within the Nkx6.1+, Nkx2.2+, and Pax7+ domains of mesencephalic explants. Finally, we demonstrate that our observations are not specific for embryonic mesencephalic cells, as NPCs in the adult subventricular zone also showed an intrinsic fate switch from neuronal to glial potential upon neurosphere amplification. Our data suggest that neurosphere formation does not expand the endogenous neurogenic NPCs but rather promotes amplification of gliogenic precursors that do not respond to niche-derived signals of cellular specification and differentiation.
Introduction
N
Although neurospheres maintain multi-lineage differentiation potential upon serial passages in culture, only a minor proportion of the cells are neurogenic, whereas the majority of cells give rise to astrocytes [2,7,8]. In most cases, the glial-biased differentiation of neurosphere cells does not significantly change after transplantation to the embryonic or adult brain [9,10]. However, it is not clear as to what extent these observations reflect the glial commitment of donor cells or the presence of gliogenic signals within the host tissue.
NPCs in the ventral mesencephalon (VM) are responsible for generating the largest nucleus of dopaminergic neurons in the mammalian brain [11]. Therefore, neurospheres derived from the VM could provide a suitable source of dopaminergic progenitors in culture. However, few mesencephalic NPCs differentiate into midbrain dopaminergic (mDA) neurons in vitro after neurosphere formation [8,12,13]. Besides, in vitro dopaminergic differentiation of expanded mesencephalic NPCs remains highly inefficient after treatment with mDA-inductive signals such as Shh, Fgf8, and Wnt5a [14 –16]. Likewise, the stromal cell-derived inducing activity of the PA6 line, while promoting dopaminergic specification of embryonic stem cells (ESCs), does not have the same effect on mesencephalic neurospheres [17]. Finally, over-expression of mDA transcriptional determinants such as Lmx1a, Nurr1, Msx1, Ngn2, and Ptx3 does not significantly enhance the yield of mDA neurons from murine and human mesencephalic neurospheres [18].
In contrast to the aforementioned studies, other reports have presented data suggesting dopaminergic potential of neurosphere cells as well as efficient induction of dopaminergic features with exogenous factors such as transforming growth factor β [19 –21]. Thus, the question remains as to whether mesencephalic NPCs change their differentiation potential after neurosphere formation or whether they require specific culture conditions or patterning cues to achieve efficient dopaminergic differentiation.
To contribute to solve this issue, we used a previously reported explant culture system [22] to expose ventral mesencephalic neurospheres to the niche of dopaminergic neurogenesis in the early embryonic midbrain. We found that mesencephalic NPCs expanded as neurospheres showed poor neurogenesis and dopaminergic specification in their original niche of differentiation. Remarkably, after just one passage in vitro, neurosphere cells completely lost their capacity to generate new neurons in the embryonic tissue. Instead, neurosphere cells showed niche-independent differentiation to the astrocytic lineage, indicating an intrinsic developmental restriction to glial fates. Further, neurosphere cells were largely unable to acquire region-specific positional markers at different dorso-ventral levels of mesencephalic explants, suggesting that neurosphere formation significantly abolishes responsiveness to patterning cues. Using the same explant culture approach, we present data supporting a fate switch from neurogenic potential to glial commitment of adult NSCs expanded as neurospheres.
Materials and Methods
Neurospheres culture
Transgenic mice that constitutively produce green fluorescent protein (GFP; gift from Andras Nagy) were crossed with wild-type CD-1 mice to obtain Gfp hemizygous (Gfp +/0) E11.5 embryos. The most ventral region of E11.5 mesencephalons was dissected for cell culture (Fig. 1). VMs were pooled and treated for 20 min at 37°C with 0.1% trypsin (Invitrogen) diluted in Versene (EDTA 0.02%; Invitrogen). Trypsin was then inactivated with neurosphere culture media (see below) containing 10% fetal bovine serum (FBS; Invitrogen). The tissues were mechanically dissociated with a serological pipette until a single-cell suspension was obtained. The cell suspension was centrifuged at 3,000 rpm for 5 min and resuspended in serum-free neurosphere culture media composed of high-glucose Dulbecco's modified Eagle's medium (DMEM) and F12 1:1 v/v supplemented with 25 μg/mL insulin, 100 μg/mL transferring and glutamax 1× (all from Invitrogen), and 20 nM progesterone, 60 μM putrescine, 30 nM selenium, and 2 μg/mL heparin (all from Sigma-Aldrich). At this point, cellular viability was higher than 95% as indicated by trypan blue exclusion. Cells were plated at a density of 5×105 cells/mL in uncoated 12-well culture plates (Corning). Each well was supplemented with 20 ng/mL FGF2 and 20 ng/mL EGF (both from Peprotech). Cultures were maintained at 37°C in humidified 5% CO2-95% atmospheric air incubators for 7 days. Half of the media and growth factors were replaced every 2 days.

Experimental approach to expose embryonic and adult neurosphere cells to the niche of mesencephalic neurogenesis.
To generate secondary neurospheres, primary neurospheres derived from E11.5 VM were collected under a stereoscopic microscope and treated for 10 min at 37°C with 0.025% trypsin diluted in Versene. Trypsin was inactivated as previously described, and neurospheres were initially dissociated with a serological pipette and then with a fire-polished Pasteur pipette to completely eliminate cell clumps. Cells were centrifuged at 3,000 rpm for 5 min and resuspended in neurosphere culture media. The cell suspension was seeded at a density of 1×105 cells/mL in uncoated 12-well culture plates. Cultures were supplemented with FGF2 and EGF (20 ng/mL each) and maintained at 37°C in humidified 5% CO2-95% atmospheric air incubators for 7 days. For in vitro differentiation of embryonic neurospheres, single-cell suspensions were resuspended in FGF2/EGF-free neurosphere culture media containing 10% FBS. Cells were seeded at a density of 1×105 cells/mL in 12-well culture plates previously treated with 4 μg/mL poly-D-lysine (Sigma-Aldrich), and 5 μg/mL laminin (Invitrogen) and maintained for 7 days at 37°C in humidified 5% CO2-95% atmospheric air incubators.
To generate neurospheres from the adult mouse brain, the anterior subventricular zone (SVZ) from both hemispheres (Fig. 1) were dissected in DMEM/F12 and minced in ∼1 mm pieces. The tissues were dissociated at 37°C for 1.5 h in 3 mL of a solution containing 124 mM NaCl, 5 mM KCl, 3.2 mM MgCl2, 0.1 mM CaCl2, 26 mM NaHCO3, 10 mM D-glucose, 0.2 mg/mL kynurenic acid, 0.67 mg/mL type IV hyaluronidase (all from Sigma-Aldrich), and 1.33 mg/mL trypsin (Invitrogen). To prevent adhesion among tissues, the solution was mixed every 20 min during incubation. The solution was centrifuged at 1,500 rpm for 5 min, and 3 mL of neurosphere culture media containing 1 mg/mL trypsin inhibitor (Sigma-Aldrich) were added. The tissue fragments were mechanically dissociated with a serological pipette and centrifuged at 1,500 rpm for 5 min. The cells were resuspended with neurosphere culture media containing FGF2 and EGF (20 ng/mL each), and the suspension was filtered through a 70 μm nylon membrane (BD Falcon). Cells were seeded at a density of 1×104 cells/mL in uncoated 6-well culture plates (Corning), and cultures were maintained at 37°C in humidified 5% CO2-95% atmospheric air incubators for 7–10 days. In vitro differentiation of adult neurospheres was done as previously described [Rietze, 2001 #406].
Collagen explant culture
Mesencephalons were obtained from E10.5 CD-1 mice embryos and cultivated in a collagen matrix as previously described [22]. Explant media (heretofore referred as Optimix) was prepared by mixing Optimem with Glutamax (72% v/v; Invitrogen), DMEM-F12 (25% v/v; Invitrogen), 2 M glucose (2% v/v; Sigma-Aldrich), and GPS 1× (Invitrogen). Explants were maintained at 37°C in humidified 5% CO2-95% atmospheric air incubators for 7 days.
Cell transplantation
This procedure was essentially as previously described [22]. Neurospheres were treated for 10 min at 37°C with 0.1% trypsin diluted in Versene. Trypsin was then inactivated with Optimix containing 10% FBS. Cell aggregates were mechanically dissociated, and the resulting cell suspension was concentrated in 5–20 μL of Optimix. For transplantation, a flame-stretched Pasteur pipette coupled to a hose with a mouthpiece was used to deposit donor cells over the ventricular surface of collagen explants. Transplanted cells were allowed to attach to the tissue for 3 h at 37°C before adding the media.
Purification of NSCs from the adult SVZ
Purification of adult NSCs was based on cell diameter and low affinity to peanut agglutinin (PNA) [23]. The anterior SVZ of adult GFP+ mice was essentially removed as previously described [24]. Single-cell suspensions of SVZ were passed through a 70 μm cell-strainer (Falcon) to remove debris. At this point, donor cells were either directly transplanted or incubated for 20 min at 4°C with Phycoeritrine-conjugated PNA to proceed with purification. After incubation, the cell suspension was rinsed twice with DMEM/F12 (Invitrogen) and finally in DMEM/F12+1% Bovine Serum Albumin (Sigma-Aldrich) before sorting in an FACS II (Becton-Dickinson) flow cytometer. Sorting was based on forward scatter and selection of phycoeritrine-positive and negative groups with a 5-channel gap between groups. Adult NSCs were collected directly in NS-A basal serum-free media (Euroclone) containing 0.6% glucose, 3 mM NaHCO3, 5 mM HEPES, 2 mM L-glutamine, 0.1 mg/mL apo-transferrin, 25 μg/mL insulin, 60 mM putrescine, 30 mM selenium, and 20 nM progesterone.
Tissue processing and immunofluorescence
Collagen explants were fixed and processed for cryo-sectioning as previously described [22]. Primary antibodies were used to detect Nestin (1:500; mouse), β-III Tubulin (1:100; mouse), GFAP (1:250; rabbit), NeuN (1:400; mouse), BLBP (1:1,000; rabbit), and TH (1:500; rabbit) from Chemicon; β-III Tubulin (1:2,000; rabbit) from Covance; and Nkx6.1, Nkx2.2, and Pax7 (supernatants 1:2; mouse) from Developmental Studies Hybridoma Bank. Primary antibodies were detected with the following secondary antibodies: anti-mouse IgGs (1:1,000; goat) coupled to Alexa-Fluor 594 and 647 and anti-rabbit IgGs (1:1,000; goat) coupled to Alexa-Fluor 594 and 647 from Molecular Probes. Images were captured with a lazer scanning microscope (LSM) 510 Meta and analyzed with the LSM 5 image examiner software, both from Zeiss.
Data analysis
The data on embryonic and adult neurospheres are representative of at least 3 independent transplantation experiments. Data are presented as the average percentage±standard deviation; “n” indicates the number of GFP+ cells counted. Confocal z-dimensional montages were obtained for every cell marker, and single optical planes were examined independently to unambiguously identify double-labeled cells. Detection of a specific marker in donor cells was always confirmed by analyzing orthogonal planes within z-stacks. The pinhole aperture of all channels was maintained close to value 1.0 for high confocality, and only clearly double-labeled cells were counted.
Results
Dopaminergic differentiation depends on complex combinations of signaling factors within the niche of the VM [25]. Here, to provide mesencephalic neurosphere cells with the proper signaling networks for neurogenesis and dopaminergic differentiation, we used explant cultures of the embryonic midbrain (Fig. 1). Mesencephalic explants derived from E10.5 mouse embryos efficiently instruct dopaminergic differentiation of ESC-derived precursors and early telencephalic NPCs [22,26]. In addition, E10.5 explants have the advantage of being a robust neurogenic niche without significant glial differentiation during the culture period [22]. In our experimental approach, neurospheres were derived from the most ventral domain of E11.5 mesencephalon (Fig. 1), as this region is endowed with neurogenic NPCs that differentiate into dopaminergic neurons [27]. Mesencephalic neurospheres expanded from E11.5 GFP+ mice were primarily composed of Nestin+ cells that gave rise to neuronal and glial cells upon in vitro differentiation (data not shown). Seven days after transplantation of single-cell suspensions derived from primary neurospheres, donor cells were completely integrated into the host tissue (Fig. 2A). However, a few donor cells were β-III Tubulin+ neurons (Fig. 2A, Table 1). Accordingly, few donor cells transplanted around the ventral midline of mesencephalic explants (i.e., the niche of dopaminergic neurogenesis; Fig. 1) were TH+ dopaminergic neurons (Fig. 2B, Table 1). Consistent with these data, a large proportion of GFAP+ astrocytes and Nestin+ NPCs were observed within the donor population (Fig. 2C, Table 1 and data not shown). Interestingly, after transplantation of whole neurospheres (i.e., without cell dissociation) many host-derived GFAP+ cells appeared at the integration site (Fig. 2D), suggesting that the niche within neurospheres induces glial differentiation in the neurogenic mesencephalic tissue. Altogether, these data indicate that neurosphere formation does not amplify mesencephalic dopaminergic progenitors but committed glial precursors. It is important to remark that the low yield of neurospheres from E10.5 mouse embryos, also reported by others [28], precluded a more extensive analysis of neurospheres derived at that developmental stage. However, our preliminary data with E10.5 neurospheres indicated essentially the same conclusions just described for E11.5 neurospheres.

Primary mesencephalic neurospheres show poor neurogenic potential after transplantation to mesencephalic explants.
Data are the average percentage±standard deviation of positive donor cells for a given marker. “n” indicates the number of GFP+ cells counted.
NPC, neural precursor cell; GFP, green fluorescent protein; PNA, peanut agglutinin; SVZ, subventricular zone; ND, not determined.
Next, we asked whether those few dopaminergic neurons observed after transplantation of primary neurospheres cells resulted from amplification of NSCs or differentiation of committed neuronal progenitors. Since the latter population has limited self-renewal capacity, we decided to subculture primary neurospheres to generate secondary neurospheres (Fig. 1). This strategy promotes a significant enrichment of NSCs within the cell aggregates [2]. Thus, progenitor cells within secondary neurospheres are more likely the product of NSCs amplification in vitro. In contrast with the data on primary neurospheres, after transplantation of secondary neurosphere cells, we found no evidence of donor-derived β-III Tubulin+ (n=295), NeuN+ (n=143), or NF-M+ (n=36) neurons (Fig. 3A–C). In addition, no GFP+/TH+ cell (n=160) was observed at the site of dopaminergic neurogenesis within mesencephalic explants (Fig. 3D).
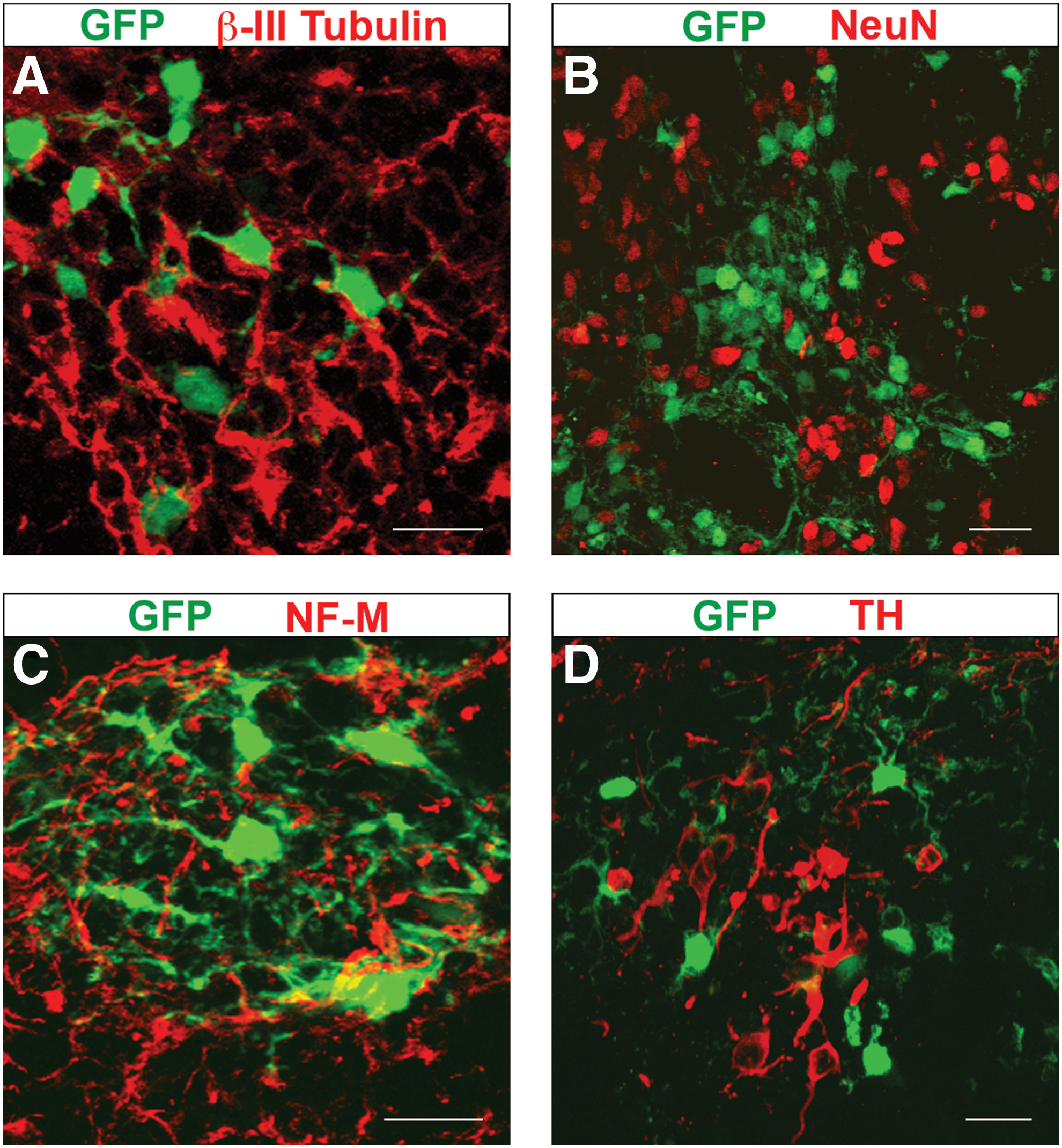
Niche-dependent neuronal differentiation of mesencephalic NPCs is completely abolished after generation of secondary neurospheres.
As expected from the absence of donor-derived neurons after transplantation, the vast majority of secondary neurosphere cells presented glial morphologies after integration into the mesencephalic parenchyma (Fig. 4A) [29]. On the basis of their branching complexity, some GFP+ cells resembled immature glial cells (Fig. 4A, left image); whereas highly branched donor cells had a similar morphology to protoplasmic astrocytes (Fig. 4A, right image) [30,31]. Although we did not find GFP+ cells with a typical morphology of mature oligodendrocytes [32,33], some donor cells could be oligodendrocyte progenitors (Fig. 4A, left image). Further, confirming the astrocytic identity of most donor cells, many GFP+/GFAP+ cells were observed within tissue sections (Fig. 4B, Table 1). In addition, many Nestin+ donor cells were found (Fig. 4B, Table 1), and some of them also produced GFAP (data not shown). Interestingly, most donor cells produced BLBP (Fig. 4C), a marker associated with radial glial cells, and reactive astrocytes [34]. These data indicate that, when analyzing differentiation of neurosphere cells within the context of embryonic explants, neurogenic NPCs are no longer present after subculture while glial-restricted progenitors are further amplified.

Secondary neurosphere cells show exclusive glial differentiation after transplantation to mesencephalic explants.
Previous reports have shown that NPCs grown as neurospheres acquire the correct positional identity after ectopic transplantation into organotypic explant cultures [35,36]. On the other hand, resembling the mechanisms of neuronal subtype specification at early stages of development [37], astrocytic diversification also depends on positional specification of glial precursors [38]. Consequently, we asked whether neurosphere-derived astroglial cells acquired region-specific positional information upon integration into distinct dorso-ventral domains of the mesencephalon in culture (Fig. 1). In contrast with previous evidence of ectopic specification of neurosphere cells [35,36], we did not find a single GFP+ cell (n=270) acquiring proper positional identity at the Nkx6.1+, Nkx2.2+, and Pax7+ domains of mesencephalic explants (Fig. 5).
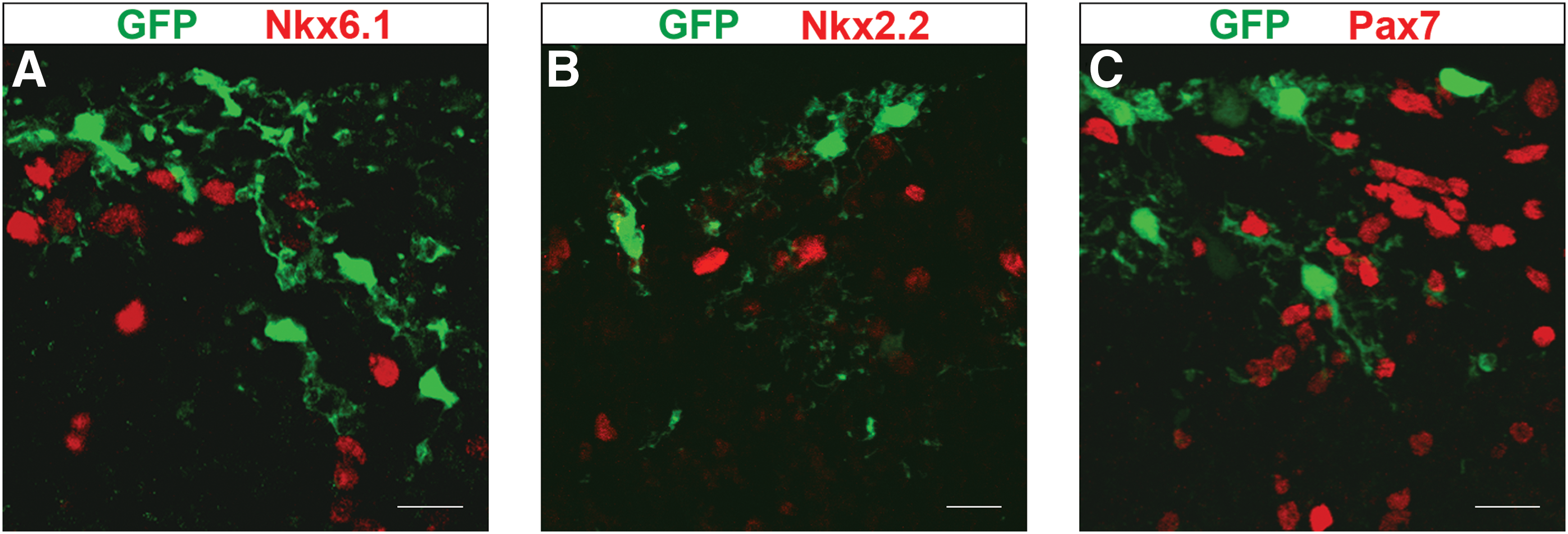
Mesencephalic neurosphere cells do not acquire the endogenous positional identity after transplantation to mesencephalic explants.
To gather more evidence on the effect of neurosphere expansion on the neurogenic potential of NSCs, we decided to extend our analysis to the adult brain. Although neurospheres have been derived from the adult midbrain [39], it is a matter of debate as to whether this region of the mammalian brain maintains NSCs with neurogenic potential during adulthood [40,41]. Therefore, we chose to transplant neurospheres derived from the SVZ of the adult forebrain (Fig. 1), a niche endowed with NSCs that provide new neurons to the murine olfactory bulb [42].
To support any finding with adult neurospheres, transplantation of freshly isolated adult NSCs was carried out (Fig. 1). Dissociated cells from the adult SVZ went through the ventricular surface of mesencephalic explants and developed neuronal and glial morphologies in deeper layers of the tissue (Fig. 6A). This observation was confirmed by detection of β-III Tubulin or GFAP in GFP+ cells (Fig. 6B, C). Interestingly, most GFAP+ donor cells produced Nestin (Fig. 6B), suggesting the presence of adult astrocyte-like NSCs [42,43]. Some transplanted cells neither acquired neural morphology nor expressed any of the evaluated cell markers (data not shown). To confirm ectopic neurogenesis of forebrain cells, FACS-purified NSCs from adult SVZ were directly transplanted into mesencephalic explants. Purification was based on sorting cells bigger than 12 μm with low affinity to peanut agglutinin (>12 μm PNAlo) [23]. This protocol yields a significant enrichment of adult NSCs with about 1 in every 7 cells giving rise to a clonal neurosphere in vitro [23]. Data obtained from 3 independent purification/transplantation experiments demonstrated that a high proportion of adult NSCs differentiated into β-III Tubulin+ neurons after incorporation into the mesencephalon in culture (Fig. 6D, Table 1). In this case, GFAP+ donor cells were not found, whereas some GFP+/β-III Tubulin- cells had a morphology of neural progenitors (data not shown). These data suggest that adult NSCs from the SVZ have the capacity to exclusively differentiate into neurons in response to embryonic neurogenic factors.
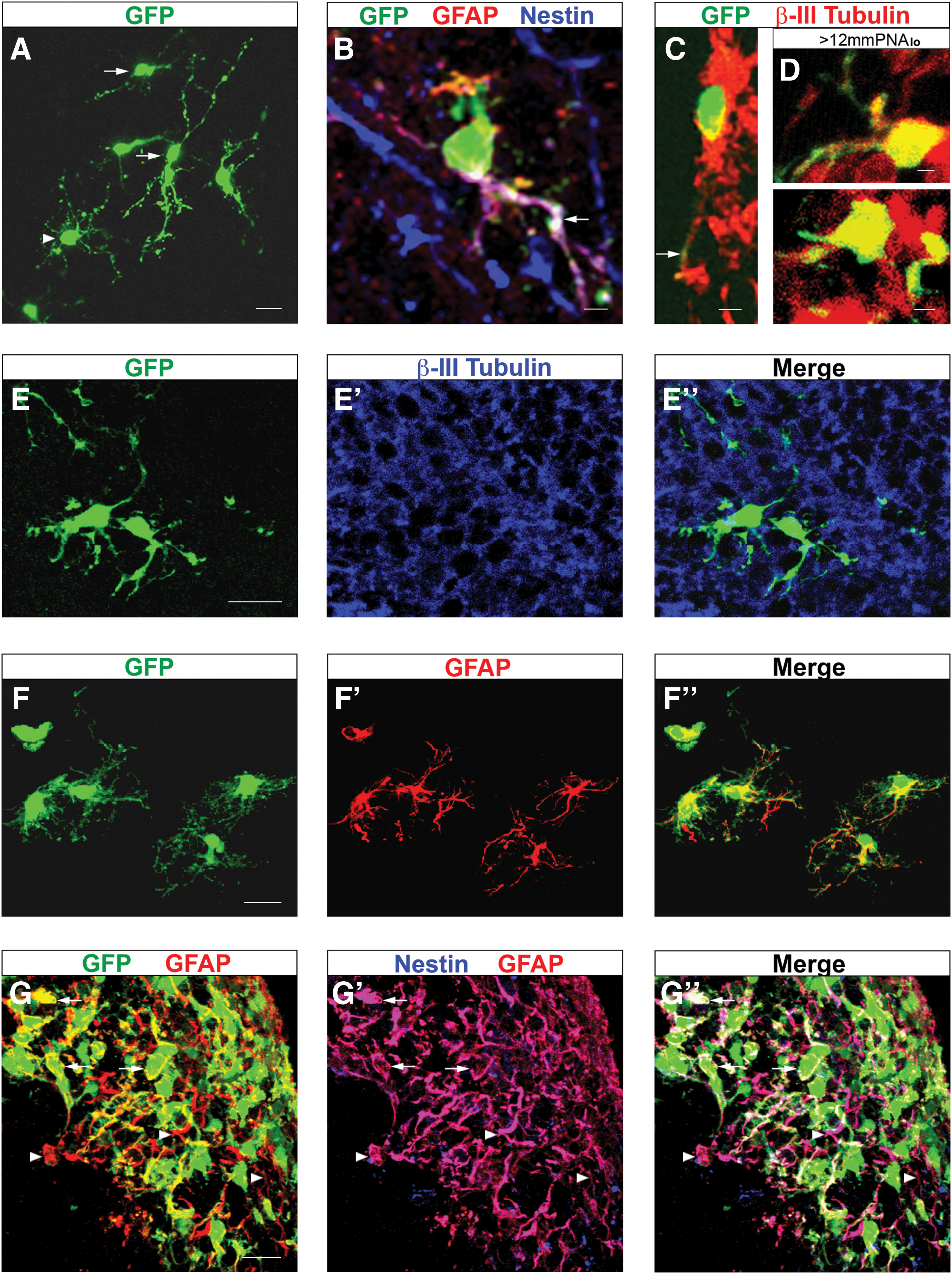
Adult NPCs lose their capacity to produce neurons in the embryonic brain after neurosphere expansion.
Next, we derived and transplanted neurospheres from the adult SVZ (Fig. 1). As previously reported, primary neurospheres from the adult SVZ had self-renewal capacity and always gave rise to neurons, astrocytes, and oligodendrocytes upon in vitro differentiation [23]. In contrast, after transplantation of adult neurosphere cells to mesencephalic explants, no GFP+/β-III Tubulin+ was found (Fig. 6E-E’’). Instead, almost every donor cell produced GFAP and presented mature astroglial morphology (Fig. 6F-F’’, Table 1). Similar to the embryonic neurospheres, adult neurospheres transplanted without cell dissociation induced precocious astrocytic specification at the integration site (Fig. 6G-G’’). However, most GFAP+ cells within the donor and host populations also produced Nestin (Fig. 6G-G’’), suggesting that the niche within adult neurospheres precludes terminal astroglial differentiation. Our data indicate that adult neurogenic NPCs become insensitive to neurogenic cues within explants and restrict their fate to the astrocytic lineage upon neurosphere expansion.
Discussion
The present study provides the following insights regarding the differentiation potential of neurosphere-expanded NPCs: (1) the astroglial-biased differentiation usually observed after neurosphere formation is not reversed by the niche of embryonic neurogenesis, suggesting commitment to glial lineages; (2) the dopaminergic neurons derived from primary mesencephalic neurospheres are not likely the product of amplification of dopaminergic progenitors; (3) neurosphere-expanded NPCs do not respond to the patterning cues that provide positional information during early stages of development; (4) our data with adult SVZ cells suggest that the NPC plasticity changes observed after neurosphere amplification are stage- and region-independent in general.
Neuronal specification and differentiation of NPCs depends on the correct combination of intrinsic factors (i.e., transcription factors, epigenetic modifications, etc) and extrinsic factors (i.e., signals within the niche). Here, we used an explant culture system to obviate the need of providing proper extrinsic signals to NPCs, thereby revealing their intrinsic potential and fate restrictions. Our data demonstrate that mesencephalic NPCs expanded as neurospheres intrinsically lose their capacity to differentiate into neurons in their original niche of differentiation. This conclusion is supported by the fact that mesencephalic explants efficiently promote neuronal specification and differentiation of ESC-derived precursors and early telencephalic precursors [22,26]. Thus, mesencephalic NPCs expanded as neurosphere are no longer likely responsive to neurogenic patterning signals within the embryonic midbrain. This insight contributes to explain the low yield of neurons obtained from neurosphere differentiation in diverse culture conditions or after transplantation [9]. The absence of robust neurogenic potential in neurosphere cells might be the result of NPCs intrinsically changing their potency to the astroglial lineage. Alternatively, the neurosphere assay could promote the expansion of endogenous glial precursors that show some multipotential behavior in vitro. Interestingly, a recent study [28] demonstrated that the initial appearance of neurosphere-forming cells around E10.5 depends on Sox9, a transcription factor involved in determination of the astrocytic lineage [44]. In any case, the robust astrocytic differentiation of neurosphere cells in a neurogenic niche reflects a commitment to that glial lineage, rather than a fate choice.
Certainly, one important feature of the neurosphere assay is the maintenance of NPCs with neurogenic potential over multiple passages in culture [2]. However, some studies have shown that the observed in vitro neurogenic potential of neurosphere cells is no longer evident after transplantation to the neural tube [10,45]. In these cases, the question still remained as to whether the glial-biased differentiation of transplanted neurosphere cells reflected an intrinsic property of donor cells or the absence of strong neurogenic signals within the host tissue. To solve this issue, we derived neurospheres from a neurogenic stage of development (i.e., E11.5) and then transplanted these NPCs to midgestation (i.e., E10.5) mesencephalic explants. The data presented here suggest that multiple rounds of cell divisions in vitro change the identity of early neuroepithelial cells to glial-restricted progenitors that share some features (such as production of BLBP) with reactive astrocytes. Nevertheless, these cells still show multipotentiality in vitro over multiple passages. Interestingly, injured-induced proliferation of reactive astrocytes in the cortex demonstrated that these cells exclusively generate more astrocytes in vivo but behave as multipotent neurosphere-forming cells in vitro [46]. Apparently, this is also the case for progenitor cells in the SVZ, where EGF-infusion ceases neurogenesis and promotes astrocyte differentiation in situ [47]. Surprisingly, however, these astrocytes generate multipotential neurospheres in vitro [47].
Some studies have claimed significant mDA neuron differentiation of neurosphere cells [19 –21]. However, these reports are difficult to compare with our data, as they only show total numbers of neurons or percentages relative to controls [19 –21]. In addition, important differences might arise from the use of primary neurospheres as opposed to sub-cultured neurospheres. Here, we show that primary neurosphere cells do have neurogenic potential (albeit low), and some cells are even specified into mDA neurons after transplantation. However, the lack of neuronal differentiation after transplantation of secondary neurosphere cells makes unlikely that those neurons observed with primary neurospheres were indeed the product of mDA progenitor expansion in vitro. The decrease in neurogenic potential observed in the neurosphere assay might be a general feature that inherently results from expansion of NPCs in a context independent of their niche, regardless of the amplification technique. For example, in vitro proliferation of mesencephalic NPCs in adherent conditions also reduces their capacity to generate mDA neurons [14].
In this study, the absence of neurons derived from secondary neurospheres should be interpreted at the population level, as our data cannot rule out the possibility of rare neurogenic NSCs unsuccessfully homing into the explants. In principle, homing of rare multipotent NSCs would involve integration into the tissue, proliferation, and differentiation into committed neurogenic progenitors. This process could take longer than 7 days and, therefore, it was unnoticed by the time of our analysis. However, it is worth noting that using the same experimental system, non-neuralized ES-derived cells and uncommitted neural precursors from early mid- and fore-brain efficiently engraft into the mesencephalic niche and differentiate according to the integration site [22,26]. The lack of unique markers to identify mesencephalic NSCs prevents us now from directly evaluating the role of homing in grafting efficiency, but future experiments with more homogenous populations of putative NSCs will help solve this issue. The data presented here suggest that at least the bulk population of heterogeneous NPCs transplated to the mesencephalon change their potency from neurogenic to gliogenic during neurosphere expansion.
The change in potency of NPCs upon neurosphere formation likely depends on the activity of specific signaling pathways. It is known that the Notch family of transmembrane receptors controls the transition from neurogenic NPCs to astrogliogenic NPCs during development [48,49]. Although the Notch pathway is not necessary for neurosphere formation, it plays an important role in the maintenance of NSCs self-renewal [50]. Further, knocking out the gene coding for the Notch1-ligand Dll1 significantly enhances neuronal differentiation of neurosphere cells at the expense of astrocytes [7]. Interestingly, a short pulse of Notch activity promotes an irreversible fate switch of NPCs to the astroglial lineage [7]. An important goal for the future will be to test whether inhibition of Notch1 activity during neurosphere formation increases responsiveness of NPCs to their original neurogenic niche.
A number of studies have determined the positional identity of NPCs expanded as neurospheres [8,16,35,36,51]. Two of these papers have alleged that murine and human neurosphere cells change their positional identity after being exposed to ectopic niches within organotypic explants [35,36]. In particular, Hitoshi et al. used LacZ under the control of a Dlx enhancer to demonstrate that cortical and midbrain neurospheres specifically activate reporter gene expression after integration into slices of ventral telencephalon [35]. Nevertheless, this article does not provide clear evidence of regional specification at the cellular level, as LacZ+ donor cells are not well defined [35]. On the other hand, Kim et al. employed specific amplification of human transcripts after transplantation of human neurospheres into murine explants [36]. Thus, these data exclusively indicate changes in the transcriptional profile of donor cells, whereas modifications in the respective protein levels were not determined [36]. In contrast, our study involved a detailed confocal analysis of hundreds of individual GFP+ neurosphere cells to look for the presence of specific transcription factors (i.e., Nkx6.1, Nkx2.2, and Pax7). Our data support the absence of regional specification of neurosphere cells integrated into multiple cellular compartments of mesencephalic explants. This phenomenon might be related to the lost of response to patterning cues. In consistency with this assumption, mesencephalic neurosphere cells do not significantly change their transcriptional profile in vitro after exposure to morphogens such as Shh, retinoic acid, and several FGF family members [16].
Our data indicate that adult NSCs of the anterior SVZ exclusively differentiate into neurons in mesencephalic explants. Accordingly, a previous work demonstrated that SVZ cells show neuronal markers after transplantation to the embryonic brain [52]. However, since NSCs were not purified in that study, it remained unclear as to whether NSCs differentiated into neurons within the host environment or rather some neurons were already present before transplantation [52]. Here, through a purification protocol based on the selection of >12 μm PNAlo cells, we provide evidence that NSCs from adult SVZ differentiate into neurons within the embryonic milieu. Ectopic neuronal differentiation could result from interactions of adult NSCs with a permissive niche (i.e., that allows differentiation) or an instructive niche (i.e., that directs differentiation). Notably, NSCs and even committed neuronal progenitors from the SVZ mostly generate glial cells after ectopic incorporation into no neurogenic regions of the adult brain, such as the striatum [53,54]. Hence, although we cannot discern between an instructive and permissive mechanism from our data, the selective neuronal differentiation of adult NSCs suggests an influence of the mesencephalic explants in promoting neurogenesis.
Remarkably, primary neurospheres derived from the adult SVZ exclusively gave rise to astrocytes after transplantation. In light of the strong neurogenic potential of freshly isolated cells from the adult SVZ, it is unlikely that our observations are just the result of exposing adult neurosphere cells to an ectopic niche of embryonic neurogenesis. Interestingly, something similar to our data was observed in vivo after infusion of EGF into the anterior ventricle of the adult murine brain [47]. In that case, NPCs within the adult SVZ no longer produced new neurons for the olfactory bulb and differentiated in situ into astrocytes [47]. The possibility remained, however, that the EGF infusion disrupts the adult niche of neurogenesis, instead of intrinsically changing the potency of adult NPCs. Here, through exposure of NPCs to EGF ex-vivo during neurosphere formation, we show that this phenomenon is likely an intrinsic change in potency of adult NPCs.
Several applications have been proposed for the neurosphere culture paradigm [6]. Neurosphere generation is still the most common in vitro assay to test for the presence of long-term proliferating NSCs in a region of the embryonic and adult brain. However, the evidence presented here indicates that the neurosphere assay could suffer of severe limitations as a model system of neural development or as a source of transplantable neurons. Future research might provide important insights into the molecular mechanisms that restrict the developmental potential of neurosphere cells. In addition, the search of new strategies to expand neurogenic NPCs in vitro remains an important challenge for future therapeutic purposes.
Footnotes
Acknowledgments
We are grateful to Elizabeth Mata, Sergio González, and Marcela Ramírez for assistance in mice care and reproduction. This work was supported by Project Program Grant IMPULSA-UNAM 03 and Consejo Nacional de Ciencia y Tecnología (50956-Q).
Author Disclosure Statement
No competing financial interests exist.