
Abstract
The mitochondrial contribution to the maintenance of human embryonic stem cell (hESC) pluripotency and culture homeostasis remains poorly understood. Here, we sought to determine whether hESC adaptation to different feeder-free culture conditions is linked to a mitochondrial adaptation. The expression of ESC pluripotency factors and parameters of mitochondrial contribution including mitochondrial membrane potential, mtDNA content, and the expression of master mitochondrial genes implicated in replication, transcription, and biogenesis were determined in 8 hESC lines maintained in 2 distinct human feeders-conditioned media (CM): human foreskin fibroblast-CM (HFF-CM) and mesenchymal stem cell-CM (MSC-CM). We show a robust parallel trend between the expression of ESC pluripotency factors and the mitochondrial contribution depending on the culture conditions employed to maintain the hESCs, with those in MSC-CM consistently displaying increased levels of pluripotency markers associated to an enhanced mitochondrial contribution. The differences in the mitochondrial status between hESCs maintained in MSC-CM versus HFF-CM respond to coordinated changes in mitochondrial gene expression and biogenesis. Importantly, the culture conditions determine the mitochondrial distribution within the stage-specific embryonic antigen 3 positive (SSEA3+) and negative (SSEA3−) isolated cell subsets. hESC colonies in MSC-CM display an “intrinsic” high mitochondrial status which may suffice to support undifferentiated growth, whereas hESC colonies maintained in HFF-CM show low mitochondrial status, possibly relying on the production of autologous niche with higher mitochondrial status to support pluripotency and culture homeostasis. Pluripotency markers and mitochondrial status are concomitantly reverted on changing the culture conditions, supporting an unrecognized role of the mitochondria in response to hESC culture adaptation. We provide the first evidence supporting that hESCs adaptation to different feeder-free culture systems relies on a mitochondrial response.
Introduction
A
In this context, it is important to address how, and to what extent, changes in the energy metabolism of hESCs contribute to their adaptation to different culture conditions. It is plausible that continuous dynamic variations in the microenvironment, mediated by either cell-cell interactions or soluble extrinsic factors, contribute to maintaining the balance between self-renewal and differentiation of hESCs through a mitochondrial-metabolic adaptation process. Accordingly, although recent work has begun to shed light on the mitochondrial response during hESC differentiation and the regulation of the cell antioxidant defense mechanisms [3 –6], the mitochondrial contribution to the maintenance of hESC pluripotency and overall hESC culture homeostasis still remains very poorly understood.
Very recently, it has been shown that human-induced pluripotent stem cells (hiPSCs) show a mitochondrial status, antioxidant machinery, and senescence-related mitochondrial pathways very similar to those of hESCs [4,7], suggesting that during the reprogramming process of somatic adult cells to an ESC-like status, the mitochondria seems to undergo a parallel and coordinated reprogramming process, therefore highlighting the potential key role of the mitochondrial contribution and biogenesis in pluripotent stem cell biology. In the present work, we sought to determine whether the adaptation of different hESCs lines to distinct feeder-free culture conditions involves a mitochondrial response.
Materials and Methods
hESCs maintenance and production of feeders-CM
H1, H9, AND1, AND2, AND3, HS181, ES2, and VAL3 hESC lines were used in the present study [8,9]. hESCs were maintained in a feeder-free culture on matrigel-coated T25 flasks (BD Biosciences) in MSC-CM or HFF-CM [2,10]. hESC cultures were maintained for 23–37 passages with daily media changes and were split (1:2) weekly using collagenase IV (Gibco) [11 –13]. In some experiments, the H9 cell line maintained in HFF-CM was cultured for 6–8 passages in the commercial defined media mTesR1 (Stem Cell Technologies) [14].
The MSC-CM and HFF-CM were prepared and collected as described in detail for MEFs CM [2,15,16]. The basal media used to prepare CM consisted of 80% knock out (KO)-Dulbecco's modified Eagle's medium (DMEM) supplemented with 20% KO Serum Replacement, 1% nonessential amino acids, 1 mM L-glutamine, 0.1 mM β-mercaptoethanol, and 8 ng/mL of bFGF (all from Invitrogen). The HFFs were purchased from ATCC (SCD-1112SK). Human MSCs were obtained from postnatal adipose tissue from healthy donors on informed consent as previously described [17,18]. For routine maintenance, HFFs and MSCs were grown in IMDM and advanced-DMEM, respectively, supplemented with 10% FCS and 2 mM L-glutamine. Cells were split 1:2 when they reached 80%–90% confluence. Human MSCs were fully characterized and showed typical fibroblast-like morphology, MSC immunophenotype (CD44+CD90+CD73+CD105+CD45-CD34-CD14-HLA-DR-), and in vitro differentiation capacity into osteoblasts, adipocytes, and chondrocytes [10,19 –21].
To achieve high standards of reproducibility for the mitochondrial study, all the cell lines were harvested and, all the measurements performed during the exponential growth phase (between day 3 and 4 after passage). The timing for daily media change, culture passage frequency, density of hESC plating, and confluence of the hESC cultures at the harvesting time were strictly controlled and maintained constant for the 8 hESC lines studied. The CO2 (5%), O2 (20%), and humidity (95%) levels were constant throughout the study for the different hESC lines.
Quantification of SSEA-3 expression by flow cytometry and cell sorting
The hESC cultures were dissociated with trypsin-EDTA, and the single cell suspension was stained at a concentration of 2–5×105 cells/mL with anti-SSEA-3-PE (BD Biosciences). Then, cells were washed and stained with 7-AAD (BD Biosciences) for 15 min at room temperature for viable cells analysis. Live cells identified by 7-AAD exclusion were analyzed for expression of stage-specific embryonic antigen 3 (SSEA-3) using a FACSCanto-II flow cytometer equipped with the FACSDiva software (BD Biosciences). Supplementary Figure S1 (Supplementary Data are available online at
The hESC lines AND1 and H1 were dissociated and stained for SSEA-3 as detailed above. Live cells identified by 7-AAD exclusion were sorted by fluorescence-activated cell sorting (FACS) based on expression of SSEA-3 using a FACSAria (BD Biosciences). Sorted SSEA-3+ and SSEA-3− cells were collected in phosphate-buffered saline+10% fetal bovine serum, pelleted by centrifugation, and directly processed for DNA/RNA extraction or stored at −80°C. Purity was consistently >90%.
Measurement of the mitochondrial membrane potential
Mitochondrial membrane potential (MMP) was measured in trypsin-dissociated hESCs as the mean fluorescence intensity (MFI) of FL2 channel after staining the cells with 1 μg/mL of JC-1 (5,5-,6,6-tetrachloro-1,1-,3,3-tetraethylbenzimidazolylcarbocyanine iodide; Molecular Probes) for 30 min at 37°C. As shown in Supplementary Fig. S2, JC-1 probe exhibits potential-dependent accumulation in mitochondria as revealed by a fluorescence emission shift from green (FL1 channel ∼525 nm) to red (FL2 channel ∼590 nm) [7]. The JC-1 probe exists as a monomer at low MMP (green fluorescence emission); whereas at higher MMP, it forms red J aggregates that exhibit red fluorescence emission. Therefore, the ratio of red/green fluorescence is an indicator of MMP. To establish the red fluorescence baseline, cells were incubated in the presence of 50 μM of the mitochondrial uncoupler CCCP+ (carbonyl cyanide 3-chlorophenylhydrazone) as detailed in Supplementary Fig. S2. The consequent mitochondrial depolarization is indicated by the decrease in the red/green fluorescence intensity.
Mitochondrial labeling by fluorescence microscopy
The hESC cultures labeled with JC-1 were analyzed by fluorescence microscopy to assess “in situ” the mitochondrial status (MMP). The hESCs were plated in matrigel-coated plates and maintained in HFF-CM or MSC-CM. In each independent experiment, all hESC lines were plated the same day at similar density, and the JC-1 labeling was performed when the cells were at the exponential growth phase 4 days after plating. The cells were washed twice with KO-DMEM and then incubated in their corresponding growth media containing 2 μM JC-1 or 2 μM JC-1+50 μM CCCP for 30 min at 37°C. Then, the cells were washed twice with KO-DMEM and maintained in HFF-CM or MSC-CM until fluorescence microscopy analysis. Pictures were captured with an Axiocam MRM digital camera (Zeiss) attached to an AxioImager A1 fluorescence microscope (Zeiss).
Quantification of mtDNA content
Total DNA/RNA from each hESC line studied was obtained using All Prep DNA/RNA kit (Qiagen). The relative content of the mitochondrial genes ND1 (NADH dehydrogenase, subunit 1), COX2 (cytochrome oxidase, subunit 2), and the noncoding 7S DNA was analyzed by quantitative polymerase chain reaction (qPCR) using TaqMan Gene Expression Assays (Applied Biosystems). Values were normalized to the nuclear 18S DNA. The expression of COX2 and ND1 was analyzed by quantitative reverse transcriptase-PCR (qRT-PCR) in triplicate for 3 independent experiments using TaqMan Gene Expression Assays. The PCR conditions were 95°C for 10 min followed by 40 cycles of 95°C for 15 s and 60°C for 60 s. Company nomenclature and code number for these TaqMan gene expression assays are shown in Supplementary Table S1.
Gene expression analysis by qRT-PCR
RNA was extracted using the All Prep DNA/RNA kit (Qiagen). RNA samples were digested with Turbo DNase (Ambion). cDNA was synthesized by using ImProm-II RT system (Promega), and the expression of pluripotency-associated genes and mitochondrial biogenesis-specific transcription factors were analyzed by qRT-PCR using TaqMan Gene Expression Assays (Applied Biosystems). Values were normalized to 18S rRNA. The expression of each gene was analyzed in triplicate for 3 independent experiments. PCR conditions were 95°C for 10 min followed by 40 cycles of 95°C for 15 s and 60°C for 60 s. Company nomenclature and code number for these TaqMan Gene Expression Assays are shown in Supplementary Table S1.
Results
Maintenance of hESCs in MSC-CM induces increased levels of pluripotency markers associated to an enhanced mitochondrial contribution
hESCs can be successfully maintained feeder free in either MEF-CM or HFF-CM or MSC-CM [2,15,16,22 –25]. However, each of these culture conditions provides dynamic variations mediated by distinct requirements of specific soluble extrinsic factors such as bFGF, TGF-β, and IGF-II [1,2,26]. We first sought to answer whether hESC adaptation to different feeder-free culture conditions may be somehow linked to a mitochondrial adaptation. To address this question, 3 hESC lines maintained in MSC-CM (AND1, AND2, and AND3) and 5 hESC lines maintained in HFF-CM (H1, H9, ES2, HS181, and VAL3) were studied. We initially explored the expression of ESC-associated markers and transcription factors in all hESC lines. The expression of the surface marker SSEA-3 is significantly higher (∼4-fold) in all hESC lines maintained in MSC-CM as compared with those hESC lines grown in HFF-CM (Fig. 1A, E). Similarly, the expression of Oct-4, Nanog and Sox-2 is consistently higher (∼2.5 to 4-fold) in hESC lines growing in MSC-CM than in HFF-CM (Fig. 1B, F).
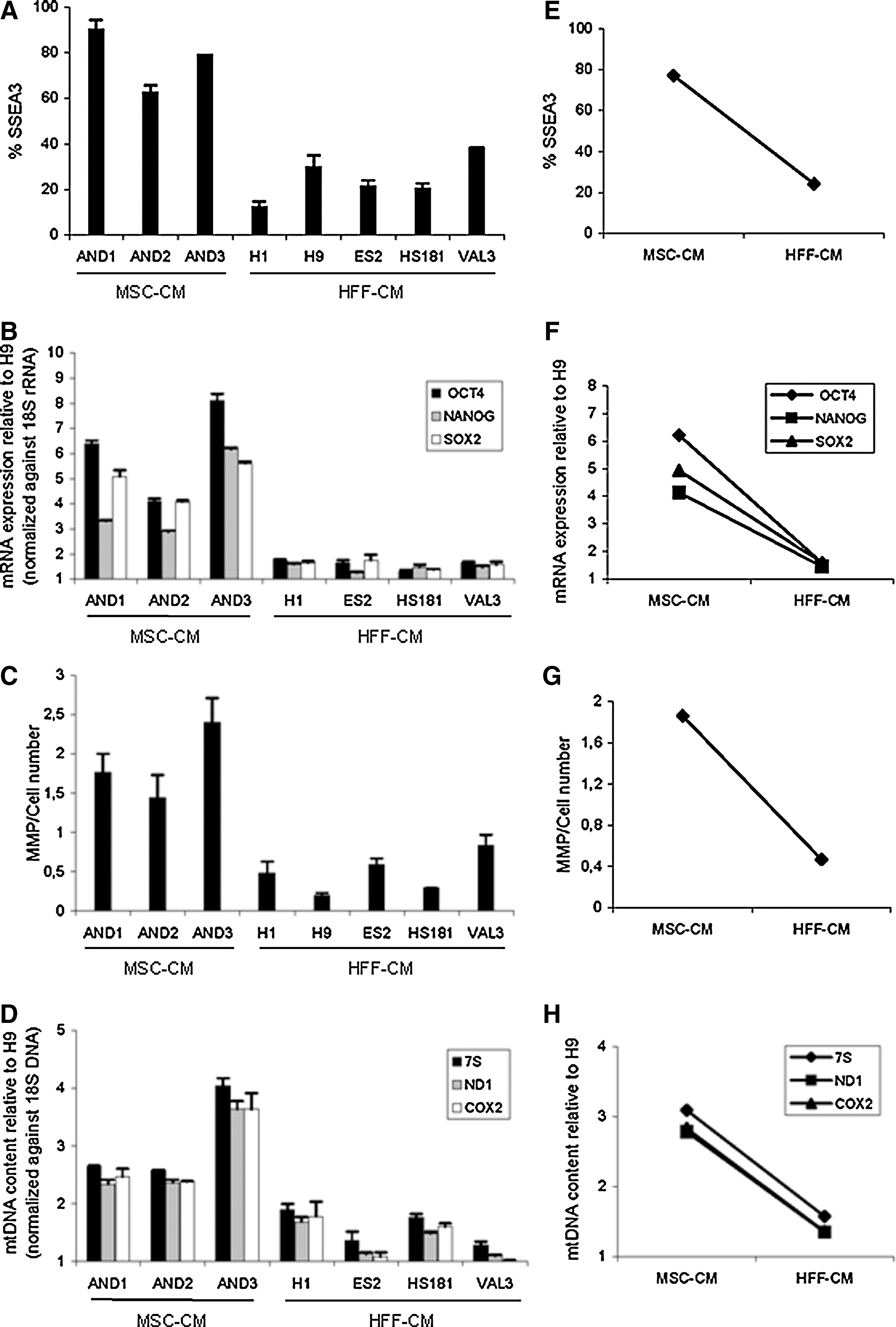
Human ESCs maintained in MSC-CM express increased levels of pluripotency markers associated to an enhanced mitochondrial status. The proportion of stage-specific embryonic antigen 3 positive (SSEA3+) cells
We next analyzed the MMP, which provides a relative measure of the mitochondrial energy-coupling and function [27 –29] and the mtDNA content for each hESC line. hESC lines growing in HFF-CM consistently displayed lower MMP (Fig. 1C, G) and a robust ∼2.5-fold decrease in mtDNA content (Fig. 1D, H) than those maintained in MSC-CM. To highlight the differential profiles for ESC pluripotency factors and mitochondrial status (MMP and mtDNA content), the data were also represented mean grouped according to the culture system used to maintain the hESC lines (Fig. 1E–H). The data identifies a parallel trend between the expression of ESC pluripotency factors and the mitochondrial status in relation to the culture conditions in which the cells were maintained, suggesting that the increased levels of pluripotency markers observed in hESCs maintained in MSC-CM are associated to an enhanced mitochondrial status.
Differences in the mitochondrial status between hESCs maintained in MSC-CM versus HFF-CM respond to coordinated changes in mitochondrial gene expression and biogenesis
Based on the differences observed in the mitochondrial status between hESC lines maintained in MSC-CM and HFF-CM, we next sought to determine whether the higher mitochondrial DNA content present in hESC lines maintained in MSC-CM responded to coordinated mechanisms of mitochondrial adaptation. It is expected that under physiological metabolic conditions, higher mitochondrial status and mitochondrial genetic load would require an increased expression of the mitochondrial molecular machinery and regulatory factors involved in the mitochondrial biogenesis. We, therefore, analyzed the expression levels of master mitochondrial genes implicated in mtDNA replication and transcription [30] including the POLG (mitochondrial DNA polymerase gamma), POLR (mitochondrial RNA polymerase), and TFAM (transcription factor A mitochondrial) as well as 2 transcription coactivators involved in the regulation of the mitochondrial biogenesis: PGC1A and PGC1B (peroxisome proliferator-activated receptor gamma, coactivator 1 alpha, and coactivator 1 beta) [30,31].
The expression of the mitochondrial genes COX2 and ND1 was ∼2.5-fold higher in hESC lines maintained in MSC-CM than in those lines grown in HFF-CM (Fig. 2A–D). Similarly, all the hESC lines maintained in MSC-CM displayed consistent higher expression levels (∼3-fold) of POLR and TFAM (Fig. 2B–E) and slightly increased expression levels (1.3-fold) of POLG (Fig. 2B–E), indicating that hESCs maintained in MSC-CM have a more active mitochondrial molecular machinery than those maintained in HFF-CM. Further, hESC lines maintained in MSC-CM also displayed a much higher expression of the mitochondrial biogenesis genes PGC1B (∼18-fold) and PGC1A (∼80-fold) than those lines in HFF-CM (Fig. 2C–F), which is consistent with a coordinated nuclear-mitochondrial regulation necessary to trigger mitochondria stimulation. These data indicate that the differences in mitochondrial DNA content and MMP found between hESC lines maintained under different feeder-free culture conditions respond to coordinated changes in the regulation of mitochondrial biogenesis and gene expression.
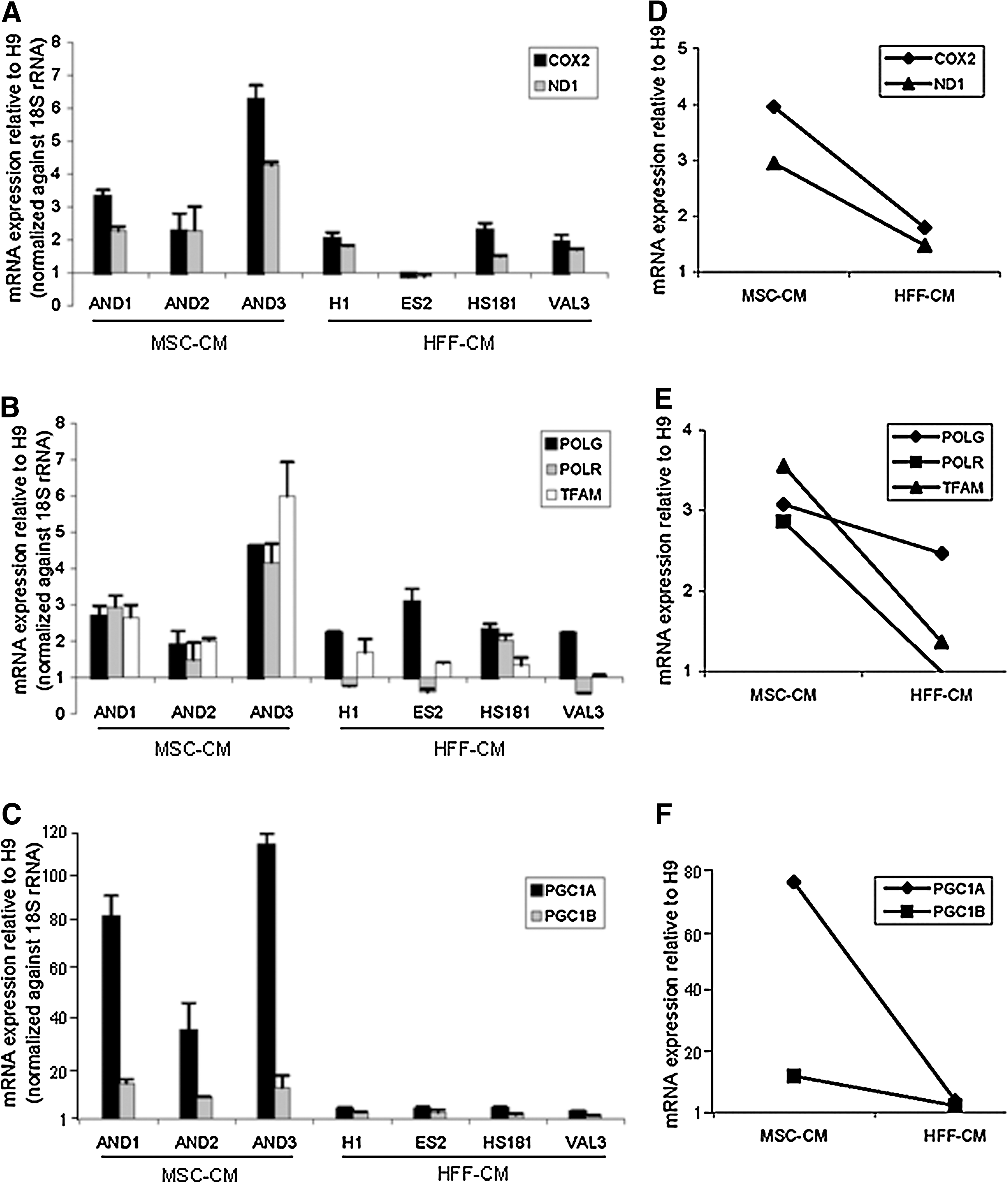
Differences in the mitochondrial status between hESC lines maintained in MSC-CM versus HFF-CM respond to coordinated changes in mitochondrial gene expression and biogenesis.
hESC culture conditions determine the mitochondrial response and cellular distribution within the SSEA-3+ and SSEA-3− cell subsets
We regularly observed that hESCs maintained feeder free in HFF-CM produce a larger amount of autologous niche (hdFs); whereas when maintained in MSC-CM, they barely produce supporting niche (Fig. 3A, B). These hdFs are mainly SSEA-3−, thus supporting the higher expression of SSEA-3 in all hESC cultures maintained in MSC-CM as compared with those in HFF-CM (Fig. 1A). This observation suggests that niche dependency contributes to the adaptation response to particular culture conditions.

The culture conditions in which hESCs are maintained determine the mitochondrial response and cellular distribution within the SSEA-3+ and SSEA-3− cell subsets. Representative phase contrast morphology
We, thus, determined whether the mitochondrial cellular distribution within the hESC cultures depends on the culture system used. We analyzed by fluorescence microscopy the cellular distribution of the mitochondria by measuring the MMP (JC-1 labelling), as a bona fide indicator of the mitochondrial polarization, in AND1 and H1 hESC cultures maintained in MSC-CM and HFF-CM, respectively. Interestingly, a strikingly different mitochondrial distribution was found between AND1 and H1 hESC cultures (Fig. 3C, D). The AND1 cultures that are composed by undifferentiated hESC colonies with limited autologous niche showed a homogenous distribution, restricted to the undifferentiated hESC colonies, of high mitochondrial status as depicted by high MMP identified by robust JC-1 red fluorescence (Fig. 3C). In contrast, H1 cultures that comprise undifferentiated hESC colonies tightly surrounded by extensive autologous niche (Fig. 3B) showed a heterogeneous mitochondrial distribution characterized by low mitochondrial (MMP) status within the undifferentiated colonies as displayed by JC-1 green fluorescence and higher mitochondrial status specifically associated to the surrounding niche (hdFs; Fig. 3D). Importantly, the average mitochondrial status was much higher in AND1 than in H1 hESC cultures (Figs. 3C, D and 1C, G). Since AND1 cultures have higher expression levels of pluripotency-associated factors than H1 (Fig. 1A, B), it is likely that a more efficiently coupled mitochondrial function is associated with higher expression levels of pluripotency factors in hESC cultures maintained in MSC-CM.
To confirm the cellular distribution of the mitochondrial status in hESCs maintained in MSC-CM and HFF-CM, SSEA-3+ and SSEA-3− cell subsets were prospectively isolated (Fig. 3E) from both AND1 and H1 hESC cultures by flow cytometry; and the mitochondrial genetic load, gene expression, and biogenesis were assessed in these purified SSEA-3+ and SSEA-3− cell subsets (Fig. 3F, G). In line with the JC-1 labeling data, AND1 SSEA-3+ cells showed a ∼4-fold higher mtDNA content (Fig. 3F) and ∼2.5- to 6-fold higher expression levels of the mitochondrial mRNAs, COX2, ND1, TFAM, and PGC1A (Fig. 3G) than AND1 SSEA-3− cells. In contrast, H1 SSEA-3+ cells displayed lower mitochondrial genetic load and mitochondrial-specific biogenesis factors than the H1 SSEA-3− cells (Fig. 3F, G). These data indicate that hESC colonies in MSC-CM display an “intrinsic” high mitochondrial status which suffices to support undifferentiated growth and culture homeostasis, whereas hESC colonies maintained in HFF-CM show a lower mitochondrial status, possibly relying on the production of autologous hdFs with higher mitochondrial status to support pluripotency and culture homeostasis.
The expression of ESC pluripotency markers and mitochondrial status is fully reverted on changing the culture conditions
We have so far shown that the adaptation of individual hESC lines to distinct culture systems seems to rely on a coordinated mitochondrial response. However, whether the mitochondrial-metabolic response is intrinsic to inherent cellular and physiologic hESC features or, in contrast, hESC lines may adapt to distinct culture conditions by reverting their mitochondrial status still needs to be addressed. In an attempt to answer this question, the H9 hESC line maintained in HFF-CM for more than 30 passages was transferred to MSC-CM or mTeSR1 (Fig. 4A), a chemically defined media formulated for feeder-free culture. After 8 passages of parallel culture in HFF-CM, MSC-CM, and mTeSR1, the original H9 hESC cultures maintained in HFF-CM displayed quite abundant autologously produced hdFs (Fig. 4A), whereas those H9 hESC cultures maintained in MSC-CM or mTeSR1 barely showed autologous niche, the culture being mainly comprised by undifferentiated colonies (Fig. 4A). In addition, hESCs transferred from HFF-CM to MSC-CM or mTeSR1 showed ∼3- to 7-fold and ∼5- to 10-fold higher, respectively, expression levels of Oct4, Nanog and Sox2 as compared to HFF-CM (Fig. 4B).

The expression of ESC pluripotency markers and factors associated to mitochondrial contribution/status is fully reverted on changing the culture conditions.
We finally wanted to confirm whether this reversion in culture morphology and expression of ESC-associated factors on changing the culture conditions is accompanied by a parallel reversible adaptation in the mitochondrial status. In fact, H9 cultures transferred from HFF-CM to MSC-CM or mTeSR1 revealed a ∼2.5- and ∼4-fold, respectively, higher mtDNA content (Fig. 4C). Similarly, on transfer to MSC-CM or mTeSR1, H9 hESCs displayed significantly higher (∼5 to10-fold) expression levels of the mitochondrial genes COX2 and ND1, the transcription factor TFAM, and especially higher levels (over 50-fold increase) of the biogenesis-associated gene PGC1A. Of note, a concomitant/parallel reversion was found for both the expression of factors associated with pluripotency and mitochondrial response.
Discussion
Despite recent efforts to develop hESC feeder-free and/or serum-free culture systems, very little is known about the underlying mechanisms and microenvironment factors maintaining pluripotency and genomic stability [12,22 –24]. We have recently shown that hESCs maintained feeder free in MEF-CM displayed distinct soluble factors required for the successful maintenance of hESC culture homeostasis and pluripotency than hESCs maintained in human-feeders CM [1,2,8,32]. Additionally, the environmental conditions provided by distinct human feeders (HFFs vs. MSCs) may also differ; and, therefore, the supporting mechanisms when using HFF-CM for undifferentiated growth of hESCs might also be different from those of MSC-CM [26].
hESCs are thought to be a product resulting from cell culture adaptation, because the properties of the inner cell mass are transient in nature, whereas cultured hESCs divide indefinitely without losing pluripotency. The study of the energetic and oxidative metabolism of hESCs has barely been studied, and the role played by the mitochondria in hESC biology remains to be elucidated. How mitochondrial adaptation as a response to different culture systems may affect the pluripotency of the hESC lines still needs to be addressed [33 –36]. In the past few years, several reports have begun to answer some of these open questions, particularly focused on the mitochondrial biogenesis progression through hESCs differentiation [3,5,6,37]. Human iPSCs were very recently shown to have a mitochondrial status and an antioxidant machinery very similar to those of hESCs [4,7], suggesting that during the reprogramming process of somatic adult cells to an ESC-like status the mitochondria undergoes a parallel and coordinated “metabolic reprogramming process,” further highlighting the need to study the potential role of the mitochondrial contribution and biogenesis in pluripotent hESC biology.
In the present work, we show that hESCs grown in HFF-CM display quite abundant autologously produced hdFs while hESCs in MSC-CM or mTeSR1 barely show autologous niche. Besides, hESCs maintained in HFF-CM show significantly lower expression levels of the ESC-associated surface marker SSEA-3 and transcription factors including Oct4, Nanog, and Sox2. We also provide the first evidence supporting that the adaptation of hESCs to different feeder-free culture systems depends on a mitochondrial response. We explored the mitochondrial response to 2 different feeder-free culture systems among an extensive cohort of hESC lines. Our results reveal a parallel and robust trend between the expression of ESC pluripotency factors and several measurements of the mitochondrial status/contribution (MMP, mtDNA content, and the expression of master mitochondrial genes implicated in replication, transcription, and biogenesis) in relation to the culture conditions in which the cells were maintained, indicating that the increased levels of pluripotency markers observed in hESCs maintained in MSC-CM are associated to an enhanced mitochondrial contribution. The differences in MMP and mtDNA content found between hESC lines maintained under different feeder-free culture conditions respond to coordinated changes in the regulation of mitochondrial biogenesis and gene expression. Importantly, hESC culture conditions determine the mitochondrial response and cellular distribution within the SSEA-3+ and SSEA-3− cell subsets. Thus, hESC colonies in MSC-CM display an “intrinsic” high mitochondrial status that suffices to support undifferentiated growth and culture homeostasis; whereas hESC olonies maintained in HFF-CM show a lower mitochondrial status, possibly relying on the production of autologous hdFs with higher mitochondrial status to support pluripotency and culture homeostasis. The expression of ESC pluripotency markers and mitochondrial status is fully and concomitantly reverted on changing the culture conditions, indicating that the mitochondrial-metabolic response seems to be adaptable to culture conditions rather than intrinsic to inherent cellular and physiologic hESC features. Altogether, our data support an unrecognized role of the mitochondria in response to culture adaptation and clonal evolution of hESCs. Consequently, it is plausible that perturbations in the mitochondrial status of hESCs may have detrimental consequences affecting not only hESCs pluripotency but also genomic integrity.
Several recent studies have examined the mitochondrial status in hESCs and hiPSCs maintained in mouse feeders and reported a global repression of mitochondrial activities commonly associated with undifferentiated growth of hESCs/hiPSCs, and that nonoxidative glycolisis appears to be the major source of energy [3,4,7,37]. Interestingly, however, our study based on hESCs maintained feeder free in MSC-CM reveals that higher mitochondrial activities correlate with higher expression of pluripotency markers. This discrepancy is related to the different culture conditions used, thus denoting that mitochondrial-metabolic responses are not intrinsic to inherent cellular and physiologic hESC features but instead reflect how a hESC line may adapt to distinct cultures conditions by reverting the mitochondrial status. Overall, our data indicate that hESCs maintained feeder free in human-feeders CM have the necessary mechanisms in place to rapidly modulate mitochondrial gene expression, biogenesis, and mtDNA content, which are features associated with cells displaying an elevated mitochondrial activity, relying primarily on oxidative phosphorylation to cover the energy demand [38]. Accordingly, in a very recent work, Birket et al. [39] have shown that hESCs display an elevated mitochondrial activity which produces ∼77% of the cellular ATP by oxidative phosphorylation, suggesting that the mechanisms of adaptation to the culture environment may depend on a more complex metabolic functional network than simply soluble factors-mediated signaling pathways such as bFGF, TGF-β, and IGF-II. In line with this observation, a link has been recently demonstrated between the expression of key sensors of the endoplasmic reticulum stress and pluripotency markers in different culture conditions [40].
It is desirable to find out how the maintenance of hESC cultures is affected by possible mitochondrial genome changes, considering that each hESC line was derived from embryos with different mitochondrial genetic haplotypes or polymorphic changes in their mitochondrial genome. Importantly, even genetically identical hESCs show significant cell-to-cell variability in pluripotency, gene expression, genomic stability, and differentiation potential. Identifying the origins of this variation is important, as it would help us understand why genetically identical cells can show a range of distinct responses to the environment [41]. It is envisioned that the generation of hESC lines devoid of their own endogenous mtDNA (so-called ρ0 cells) would represent a formidable “knockout” cellular system in which to precisely address questions about the role of the mitochondria in regulating the tight balance between self-renewal and differentiation as well as during the early phases of human embryogenesis.
Footnotes
Acknowledgments
P.M's Group was funded by the Innovation and Science Department of the Junta de Andalucía to P.M (P08-CTS-3678), the FIS/FEDER to P.M (PI10/00449) and to C.B (CP07/00059), the MICINN to P.M (PLE-2009–0111) and the Spanish Association Against Cancer (CI110023) to P.M. M.M's Group was funded by the Regional Ministry of Health of the Junta de Andalucía to M. M (2008/0317) and the Spanish Ministry of Science and Innovation, to M. M. (RYC-2006–001013). V.R-M was supported by the Marie Curie IIF (PIIF-GA-2009–236430). We thank professor Outi Hovatta (Karolinska Institute, Sweden) for kindly providing the HS181 hESC line.
Author Disclosure Statement
The authors report no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.