
Abstract
A complex set of extrinsic and intrinsic signals acts in specific temporal and spatial orders to enable neural differentiation during development. These processes have been extensively studied in animal models, but human neural development remains much less understood. This lack of detailed information about human early neurogenesis is a hindrance for the differentiation of pluripotent stem cell lines into specific neuronal phenotypes. Therefore, it is important to strengthen the interspecies comparative approaches. We describe a novel model system in which in vitro differentiation of human and mouse embryonic stem (ES) cells are temporally aligned to each other and compared with mouse telencephalic neurogenesis in vivo. In this comparative model system, we tested the in vitro role of Hedgehog (Hh) signaling for ES cell-derived telencephalic differentiation. In vivo, Hh signaling mediates dorsal/ventral patterning during early stages of telencephalic development. We monitored the effect of pharmacological modulators of the Hh signaling pathway, purmorphamine—an agonist and cyclopamine—an antagonist of the Smoothened receptor (Smo), on the expression of region-specific transcription factors and signaling molecules relevant for telencephalic development in vivo. Purmorphamine strongly upregulated the expression of telencephalic ventral markers Nkx2.1, Nkx6.2, Lhx6, and Lhx8 in mouse and human cells, thus reflecting the in vivo process of the medial ganglionic eminence patterning and specification. Cyclopamine upregulated the expression of telencephalic dorsal markers, but at lower levels in human compared with mouse cells. Modulation of Smo in vitro differentially affected, in mouse and human cells, the expression of molecules of the Hh pathway, especially the Gli1 and Gli3 effectors, Sonic Hh ligand and Ptch receptors. These results provide evidence for the different default differentiation of mouse and human ES cells and prove the utility of the comparative system for optimizing the directed differentiation of human pluripotent stem cells.
Introduction
T
Previous studies have reported successful differentiation of mouse embryonic stem (mES) cells [3 –6] and human embryonic stem (hES) cells [7 –9] into specific neuronal types in response to patterning factors for the anterior/posterior (A/P) and dorsal/ventral (D/V) axes of the central nervous system (CNS). These studies demonstrated that the signaling pathways specifying the major regions of the CNS in general are evolutionary conserved. However, differences in timing and response to homologous signaling mechanism between species exist. Therefore, the generation of homogeneous populations of specific neuronal subtypes, especially from hES cells, remains a difficult problem, which requires differentiation under defined and optimized in vitro conditions. Particularly challenging is the task of generating telencephalic neurons including glutamatergic projection neurons, different subtypes of GABAergic or cholinergic interneurons and projection neurons.
Recent research on early A/P patterning, D/V patterning and neuronal specification has unraveled key developmental mechanisms contributing to the generation of specific telencephalic cell types in vivo and demonstrated that distinct subclasses of neurons have distinct spatial and temporal origins [10 –16].
The secreted proteins of the Hedgehog (Hh) family play key roles during multiple phases of telencephalic development and act in temporally and spatially distinct patterns [17 –21]. At the earliest stages, Hh signaling mediates the D/V patterning. The effect of Hh loss of function was studied in the telencephalon of mutant mice by inactivation of Smo alleles in a conditional model, where Cre expression was driven by the FoxG1 promoter (Smoc/–;FoxG1Cre ) [18]. Smo is a mandatory cell-autonomous mediator of Hh signaling [22]. Smo inactivation at embryonic day 9 (E9) led to a complete loss of ventral telencephalic patterning whereas Smo inactivation at E12 did not affect telencephalic patterning [18]. Sonic hedgehog (Shh), the best characterized Smo ligand, is expressed in the mouse telencephalon starting with E8.5 [18,23]. These findings indicate that precisely timed Hh signaling between E8.5–9 and E12 is essential for patterning of ventral regions and the generation of cell types arising from there, in particular telencephalic GABAergic and cholinergic neurons.
To provide further information about neurogenesis in human cultures and to assess whether the molecular mechanisms that govern mouse neural development in vivo play similar roles in cultured human cells, we developed a novel, parallel cell culture system. Using this system, neuronal differentiation of mES cells and hES cells toward specific telencephalic phenotypes was studied in a direct comparative fashion, under defined culture conditions. We tested the specific hypothesis that the modulation of Hh signaling, in accordance with its role in D/V patterning in vivo, affects in particular the generation of telencephalic committed progenitors and specified neurons. To this end we treated the cultures with small-molecule modulators of Hh pathway at a specific time-frame corresponding to in vivo D/V patterning of mouse telencephalon [24 –27]. Hh signaling was either stimulated or inhibited by the addition of the Smo agonist purmorphamine [24,27] or antagonist cyclopamine [25,26] at concentrations established by previous experiments [5,8,28]. Gene expression analysis showed that in vitro modulation of Hh signaling elicited marked changes in transcription factor (TF) expression patterns, in both mouse and human cell culture models. We also showed how recently identified aspects of Shh dynamics and interneuron specification in the mouse embryonic telencephalon in vivo are reflected in vitro, in our ES cell-derived models.
Materials and Methods
ES cell cultures
The mES cell lines used were ES-E14TG2a (passages 9–15) [29] and B6G-2GFP (passages 16–25) [30]. The hES cell lines used were HS207 (passages 29–35) and HS401 (passages 48–54), both derived in Karolinska Institutet, Sweden [31]. The Ethics Board of Karolinska Institutet and the Regional Ethics Board in Stockholm, Sweden, gave approval for the derivation, expansion, and differentiation of hES cell lines; the Ethics Committee of the Carol Davila University of Medicine and Pharmacy, Bucharest, Romania gave approval for the expansion and in vitro differentiation of hES cell lines.
Undifferentiated ES cells were cultured as previously described [30,31], with some modifications, on a feeder layer of either mouse embryonic fibroblasts for mES cells, or human foreskin fibroblasts (ATCC) for hES cells; both feeders were inactivated with 10 μg/mL mitomycin C for 2.5 h. ES cell culture medium (ESCM) consisted of Knockout Dulbecco's modified Eagle's medium (DMEM) with Knockout serum replacement (15% for mES cells and 20% for hES cells, respectively), 2 mM Glutamax, 0.1 mM MEM nonessential amino acids, 0.5 mM 2-mercaptoethanol (all from Gibco Invitrogen Corporation) supplemented with 1,000 U/mL leukemia inhibitory factor (LIF) (Chemicon) for mES cell, and 8 ng/mL recombinant human basic fibroblast growth factor (FGF2) (R&D Systems) for hES cells. Cell cultures were maintained in a humidified atmosphere with 5% CO2, at 37°C. Medium was changed every day. mES cell colonies were passaged with 0.25% Trypsin-EDTA (Gibco) at a 1:3–1:5 split ratio every other day and hES cell colonies were passaged either with 6 U/mL of dispase (Gibco), or mechanically, at a 1:3–1:5 split ratio every 4–5 days.
Neural differentiation cultures
Neural differentiation of ES cells occurred in discrete steps, which included neural induction, neural patterning, and neuronal specification, modified from previously published protocols [6,8,32 –34]. The steps and time—frames for ES cell differentiation are presented in Fig. 1A.

Temporal alignment of mouse embryonic stem (mES) and human embryonic stem (hES) cell-derived in vitro models with in vivo telencephalic development.
Neural induction medium
Neural induction medium (NIM) consisted of DMEM/F12 with Glutamax (Gibco) containing 25 μg/mL insulin, 100 μg/mL transferrin, 5 ng/mL sodium selenite, 2.5 μg/mL fibronectin (all from Sigma-Aldrich), and 0.1 mM MEM nonessential amino acids solution. The ROCK inhibitor Y-27632 (10 μM; Sigma) was added for 8 days in NIM for hES cells, in addition to 1 h pretreatment of hES cells before starting neural induction [9].
Neural patterning medium
Neural patterning medium (NPM) consisted of DMEM/F12 with Glutamax, 1% N2 supplement, and 0.1 mM MEM nonessential amino acids, supplemented with cyclopamine (5 μM; Sigma) or purmorphamine (1–3 μM; Calbiochem).
Neuronal differentiation medium
Neuronal differentiation medium (NDM) medium consisted of Neurobasal medium with 2% (vol/vol) B27 supplement without vitamin A (Gibco), 2 mM glutamax, 1 μM cAMP, 200 μM ascorbic acid, and 20 μg/mL laminin (all from Sigma). Media were changed every 48 h.
ES cells, at differentiation day 0 (D0), were separated from feeder cells, dissociated with Accutase (Sigma), and cultured as cell suspension (2×105 cells/mL) in ESCM without LIF or FGF2 in nonadhesive bacterial-grade dishes. Cells that became floating cellular aggregates were cultured in the same medium for 2 days. The adherent colony culture was started by plating cellular aggregates on plastic or glass surface coated with polyornithine (Sigma; 15 μg/mL for plastic and 50 μg/mL for glass, 1 h RT) and laminin (20 μg/mL, ON 4°C), and further cultured in NIM. Attached aggregates flattened over 1–2 days and columnar primitive neuroepithelial (NE) cells developed and formed neural rosettes at approximately D6 for mouse and D10 for human cells. The neural tube-like rosettes formed during neural induction were used as a selection criterion; neural colonies were mechanically separated, collected, and dissociated using Accutase and then replated onto polyornithine/laminin coated plates using high density droplets (104 cells/μL). At these time-points (D6 and D10), we started the neural patterning treatment for mouse and human cells, respectively. Neural patterning was modulated with purmorphamine or cyclopamine in NPM. The treatment lasted 4 days for mouse and 8 days for human cells. For neuronal differentiation, neural progenitor clusters were either cultured in NDM, or dissociated with Accutase and plated onto polyornithine/laminin-coated coverslips, followed by culturing in NDM.
Colonies and cellular morphologies were monitored by phase contrast microscopy (Fig. 1B). ES cell or neural colonies were mechanically detached or split under a stereomicroscope. Three independent experiments were performed for each cell line. Samples for mRNA extraction or immunocytochemistry were taken at the following time-points: D0, D6, D10, and D14 for mouse cells and D0, D10, D18, and D26 for human cells.
Dissection of the mouse embryonic telencephalon
Mouse embryonic brains obtained at gestational day (E) 10.5–11 and E12.5–13 from the mouse strain C57BL/6-TgN(act-EGFP)OsbC14-Y01-FM131 (RIKEN Bio research centre) were dissected [34]. Briefly, for the ganglionic eminence (GE) dissection, a longitudinal cut was made through the medial cortex to expose the GE primordium in the floor of the lateral ventricle and the whole GE was snipped off by a superficial longitudinal cut. In E12.5-E13 brains, the medial GE (MGE) and lateral GE (LGE) were separately dissected. The cortex was separated from the rest of the telencephalon by cutting along to the lateral margin of the GE. The tissue pieces were collected for RNA extraction.
Quantitative real-time polymerase chain reaction
Messenger RNA was isolated using Dynabeads® Oligo (dT)25 (Invitrogen Corporation) following the manufacturer's protocol. mRNA was immediately used for cDNA synthesis according to High-Capacity cDNA Reverse Transcription Kit protocol (Applied Biosystems).The levels of cDNA for mouse cells were assessed by quantitative real-time polymerase chain reaction (qPCR) using Fast SYBR®Master Mix (Applied Biosystems, Life Technologies Corporation). Standard curves and melting curves were determined for each set of primers to confirm that a single amplicon was generated. Taqman assays and Taqman master mix (Applied Biosystems, Life Technologies Corporation) were used for the human cells. Detailed information about mouse primers and Taqman assays are provided in Appendix Table A1; All results from 3 technical replicates were normalized to glyceraldehyde-3-phosphate dehydrogenase and expressed as ΔCt values (low ΔCt levels indicate high expression). Relative expression ratios were calculated by the ΔΔCt method [35].
Immunocytochemistry
Adherent cells on glass coverslips were fixed with 4% buffered paraformaldehyde in phosphate-buffered saline (PBS; 0.01 M, pH 7.4), for 20 min at RT and washed twice with PBS. Permeabilization and blocking was carried out using 0.3% Triton-X-100 in PBS (PBST) with 10% goat normal serum and 1% bovine serum albumin (BSA) for 1 h at RT. Cells were incubated overnight at 4°C with primary antibodies diluted in 1% BSA in PBST, against: Pax6, Nestin (1:200, mouse IgG; Santa Cruz), Sox1 (1:200, goat IgG; Santa Cruz), FoxG1 (1:200, rabbit IgG; Santa Cruz), N-cadherin (Ncad, 1:200, mouse IgG; Millipore), brain lipid binding protein (Blbp 1:500, rabbit IgG; Millipore), Nkx2.1 (1:50 mouse IgG; Abcam), Tau (1:200, mouse IgG; Santa Cruz or 1:1,000 rabbit IgG; DAKO), GABA (1:500; mouse IgG; Sigma), vesicular choline transferase (Vacht,1:1,000; rabbit IgG; Sigma), and Shh (1:100; rat IgG; Abcam). After several rinses in PBST, cells were incubated for 1 h with AlexaFluor secondary antibodies (Alexa 488-conjugated goat anti-rabbit or goat anti-rat, Alexa 555-conjugated goat anti-mouse or goat anti-rabbit, 1:2,000; Invitrogen) and washed 3 times in PBS. The coverslips were incubated for 2 min with 4′, 6-diamidino-2-phenylindole (DAPI, 300 nM in PBS) for nuclear staining, rinsed in PBS and deionized water, and mounted on slides with Mowiol (Sigma). Specificity of the antibodies was tested on mouse embryonic brain samples. Negative controls, without primary antibody were performed in all experiments to monitor the nonspecific staining. Pictures were taken with an ApoTome Imaging System based on Axio Observer Z1 (Zeiss) using AxioVision software. The populations of Pax6, Sox1, Foxg1, and Nkx2.1 expressing cells, from the total cell number (as stained with DAPI) for 3 coverslips in each experimental group and at least 5 fields per coverslip were counted, using Metamorph software (Molecular Devices).
Statistics
For statistical analyses, the Statview (version 5.0.1) software was used (SAS Institute). Three independent biological samples for each time point contributed to the data set. Data are presented as mean±standard error of the mean. Student's t-test was applied to compare between 2 groups. When multiple groups were compared across time or treatments, repeated measures analysis of variance was employed, followed by Dunnett's or Student-Newman-Keuls post-hoc test, respectively. Differences were considered significant when P<0.05.
Results
mES and hES cells convert to neural precursors and neurons expressing telencephalic markers
Neural differentiation of 2 lines of mES cells and 2 lines of hES cells was studied under identical, defined conditions, but following different time-schedules for mouse and human cell cultures. In vitro time schedules were based on data from in vivo development as a reference and the stages of neural induction, neural patterning, and neuronal specification were analyzed (Fig. 1A). Time-dependent morphological changes from ES cells to NE cells, neural progenitor cells, and neurons, were analyzed in parallel in both in vitro models (Fig. 1B). Gene expression was analyzed at each step.
Neural induction
Consistent with previous results [32,33,36 –38], neural induction under chemically defined conditions in mES cells and hES cells led to differentiation into NE columnar cells organized in rosettes (Fig. 1B). NE cells were characterized at D6, for mES cells, and at D10 for hES cells. NE identity of cells was confirmed by their characteristic morphology, loss of pluripotent marker gene (Oct4 and Nanog) expression, and upregulation of NE marker genes (Sox2, Otx2, Nestin, and Ncad) (Appendix Fig. A1A). Pax6 and Sox1 expression was not detected at D0 but appeared after neural induction. Expression of Pax6 was detected in mouse NE cells at D6 and in human cells at D10. Sox1 expression was high in mouse at D6 but very low in human NE cells at D10 (Appendix Table A2). This finding was confirmed by immunostaining demonstrating that PAX6 protein was uniformly expressed in human NE cells (89.1%±5.3%), while Sox1 protein was uniformly expressed in mouse NE cells (75.2%±8.3%) (Appendix Fig. A1B).
In both mES and hES cell-derived cultures markers characteristic for endoderm (Gata4 and Afp), mesoderm (Brachyury), or trophectoderm (Tbr2) were expressed at very low levels or not detectable by real-time PCR following neural induction (Appendix Table A2).
These results were consistently obtained in independent experiments using 2 mES cell lines and 2 hES cell lines, at different passages. Thus, we conclude that, under chemically defined conditions of our experiments, neural induction occurs efficiently, uniformly and reproducibly in mouse and human cells but at different time-points in culture.
A/P early patterning
In the absence of exogenously added growth factors or morphogens, NE cells generated from mES cells and hES cells expressed forebrain markers, consistent with previous reports [3,9,33,36,39,40].
In particular, the telencephalic marker FoxG1, which is abundantly expressed in early telencephalic primordium and subsequently in the embryonic telencephalon [41 –43], was not detected in mES and hES cells, but was highly expressed in mouse (D6, ΔCt=5.9±0.4) and human NE cells (D10, ΔCt=3.9±0.4). During the next steps of neural differentiation, FoxG1 expression was further upregulated in the mouse culture at D10 and D14, but did not reach the same high level of expression as in the mouse embryonic telencephalon (Appendix Fig. A2A and Appendix Table A3). In human cells, FOXG1 expression was also slightly increased at D18 and D26 compared with D10 (Appendix Fig. A2A). FoxG1 expression was confirmed by immunostaining, demonstrating a large proportion of both mES and hES cell-derivatives positive for FoxG1 protein (55.2%±7.3% at D10 for mouse and 85.2%±9.1% at D18 for human) (Appendix Fig. A2B). Early anterior patterning was further confirmed by the strong expression of the forebrain-related markers Lhx2, Six3, Otx1, and Otx2 [33,44 –46], while markers specific for the midbrain (En1/2) and hindbrain/spinal cord (HoxB1 and HB9), were expressed at low levels or were not detected (Appendix Table A2).
To further document the in vitro neural induction and early anterior patterning in chemically defined conditions, without added morphogens, we analyzed the endogenous expression of signaling molecules implicated in early steps of telencephalic neural induction and anterior patterning [11,47,48], which were also included in several previously published protocols for neural differentiation of ES cells [3,40,49,50]. Members of the FGF family like Fgf8 and Fgf15 are early and abundantly expressed in the telencephalic neuroepithelium [47,48]. Dickkopf1 (Dkk1), an inhibitor of the WNT signaling pathway, and Noggin (Nog), an inhibitor of the BMP signaling pathway, are expressed in an early peak time-point in mouse telencephalon [51].
In our models, Fgf8, Fgf15 (or the human ortholog FGF19), Nog and Dkk1 were expressed at low levels in mES and hES cells, but at high levels in both mouse and human NE cells (Appendix Table A2). In mouse a culture, Fgf15 mRNA was significantly (∼4-fold) upregulated at D10 compared with D6, while at later time-points (D14) the expressions of both Fgf8 and Fgf15 decreased. In the human cultures, FGF8 expression was significantly reduced over time while FGF19 expression did not change significantly compared with D10 (Appendix Table A2). We found high expression levels for Fgf15 and Nog and low expressions for Fgf8 and Dkk1 in mouse embryonic telencephalon (Appendix Table A3).
Taken together, these results show that in our models ES cells differentiation towards telencephalic phenotype occurs without addition of morphogens in chemically defined conditions, in agreement with previous reports [3,9,33,36,39,40]. This is consistent with endogenous expression of telencephalic signaling molecules at proper stages to sustain both neural induction and anterior patterning.
Neural differentiation
We analyzed the temporal expression of various pan-neuronal and glial markers [Ncad, Nestin and Blbp for neural progenitors, Doublecortin (Dcx) and Tau for neurons; Oligodendrocyte specific protein (Osp) and Glial fibrillary acidic protein (Gfap) for glial cells] in both mouse and human cultures by qPCR. The levels were compared with the endogenous expression levels of the same markers in mouse embryonic telencephalon.
Ncad mRNA was highly expressed in both mouse (ΔCt=3.9±0.6 at D6) and human NE cells (ΔCt=2.6±0.2 at D10) and its expression level did not significantly change during neural differentiation. The level of expression in the mouse cultures was not significantly different from the levels expressed in the mouse telencephalon (Fig. 2A). Similar results were obtained for Nestin (Appendix Table A2).

The neural progenitor and neuronal markers are temporally expressed in mES and hES cell-derived models.
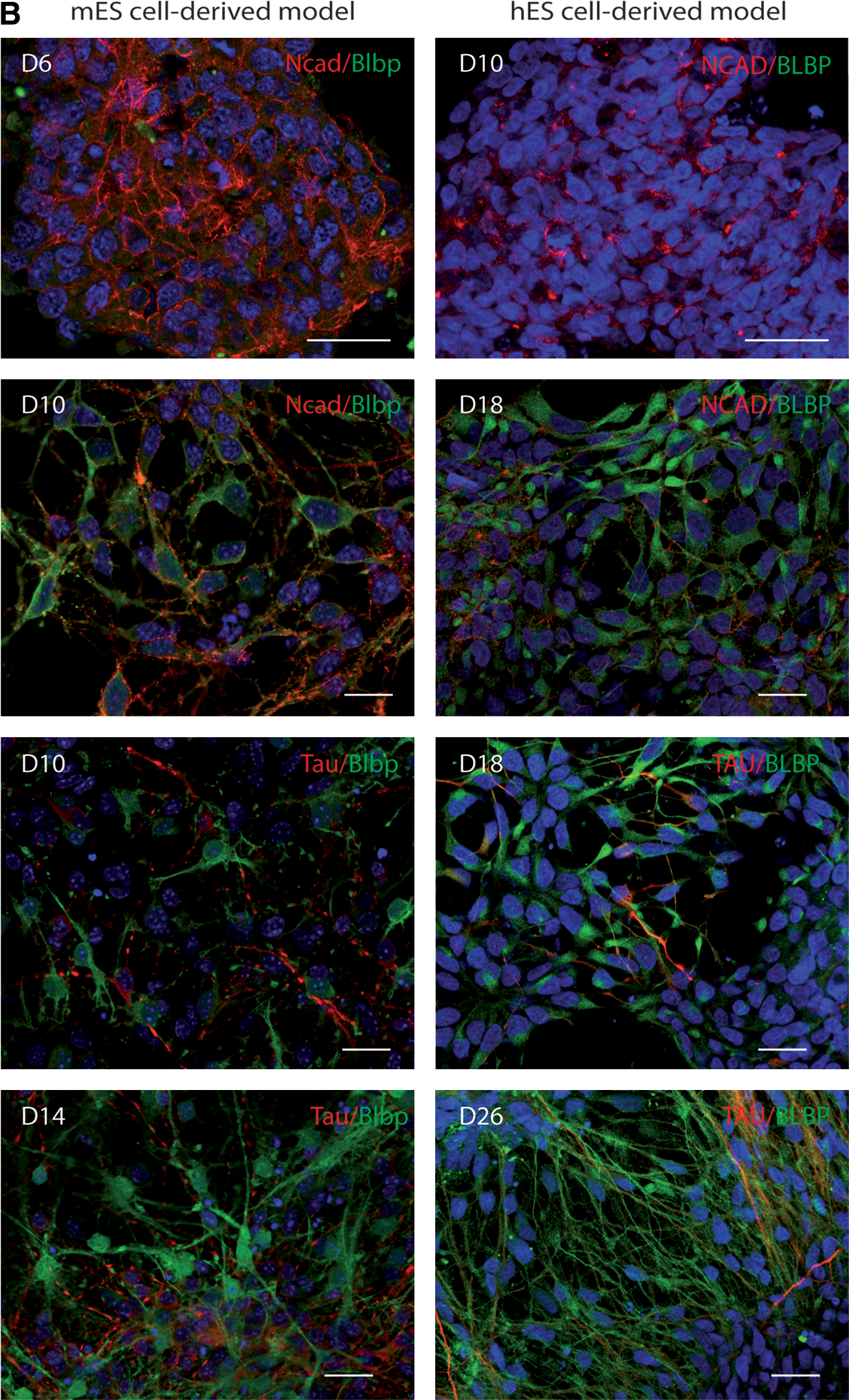
Blbp mRNA levels in both mouse and human NE cells were significantly upregulated during spontaneous differentiation. The level of expression in mouse cultures at D10 was not significantly different from the levels in the mouse E13 ventral telencephalon (Fig. 2A). The expression levels of the early neuronal marker Dcx were similar in E10.5 telencephalon and in mouse culture at D10; both levels were elevated as compared with D6. The expression level of Dcx increased at later time-points (D10 and D14 in mouse cultures and at D18 and D26 in human cultures), compared with the early stages (D6 and D10, respectively) (Fig. 2A). The pan-neuronal marker Tau was expressed at similar levels in the E10.5 ventral telencephalon and in mouse culture at D10, but lower values were obtained from the dorsal telencephalon. The expression level of Tau significantly increased subsequently (D14 in mouse culture and at D26 in human culture) (Fig. 2A).
Glial markers Osp and Gfap were not expressed in mouse or human NE cells in vitro. The level of expression at D10 in mouse culture was low similar to the level in the mouse embryonic telencephalon at E13 (Appendix Tables A2 and A3). Both markers were highly expressed at D14 in mouse cultures (Appendix Table A2). Therefore, in mouse culture, the neurogenic time-window, as aligned with in vivo development, occurs mainly between D6 and D10. After this stage, neurogenesis occurs in parallel with gliogenesis. In the human cultures, OSP and GFAP expression was very low or not detected before and at D26 but increased afterward (Appendix Table A2), further supporting the notion of a neurogenic time-window in human cultures. The expression dynamics of Ncad/Blbp in progenitors and Ncad/Tau in neurons was confirmed by immunocytochemistry (Fig. 2B).
These results demonstrate that neural differentiation takes place in mES cell-derived cultures resulting in the generation of neural progenitors and neurons in a time-frame which mirrors telencephalic neurogenesis in vivo. The expression levels of these markers are comparable between in vivo and in vitro differentiated populations. Neural differentiation in human cells can be temporally aligned with mouse cells in the proposed neurogenic time-windows.
Thus, our temporally aligned, comparative cell culture model offers a novel platform for analyzing the effect of signaling molecules on the generation of specific telencephalic populations in mouse and human cells.
Hh modulation differentially influences the expression of dorsal versus ventral telencephalic markers
To test the effect of Hh signaling pathway modulation on the D/V patterning at the telencephalic level in vitro, the pharmacological modulators purmorphamine or cyclopamine were applied between D6-D10 to mouse cultures and between D10-D18 to human cultures (Fig. 1A). As a baseline for these experiments, we determined the expression levels of Hh receptor Smo, the direct target for purmorphamine and cyclopamine, in our cell culture models and compared them to the expression levels in mouse embryonic telencephalon. Smo was expressed at high level in both mouse and human NE cells similar to mouse embryonic telencephalon. Since Smo expression remained high during the subsequent differentiation steps, we conclude that both mouse and human cell culture models are responsive to Hh modulation (Appendix Tables A2 and A3).
Next we determined the expression of TFs known to regulate patterning and specification of telencephalic progenitors and neurons in vivo: Pax6, Emx1, Emx2, Lhx2, Tbr1, and Tbr2 (dorsal markers) and Nkx2.1, Nkx6.2, Gsh2, Olig2, Dlx2, Lhx6, Lhx8, and Isl1 (ventral markers) in spontaneously differentiating cultures. The mRNA expression levels of these markers were compared with the levels in mouse embryonic telencephalon (Fig. 3A and 4A). Finally the effects of cyclopamine and purmorphamine on the D/V patterning were investigated both in mES and in hES cell-derived cultures (Fig. 3B and 4B).

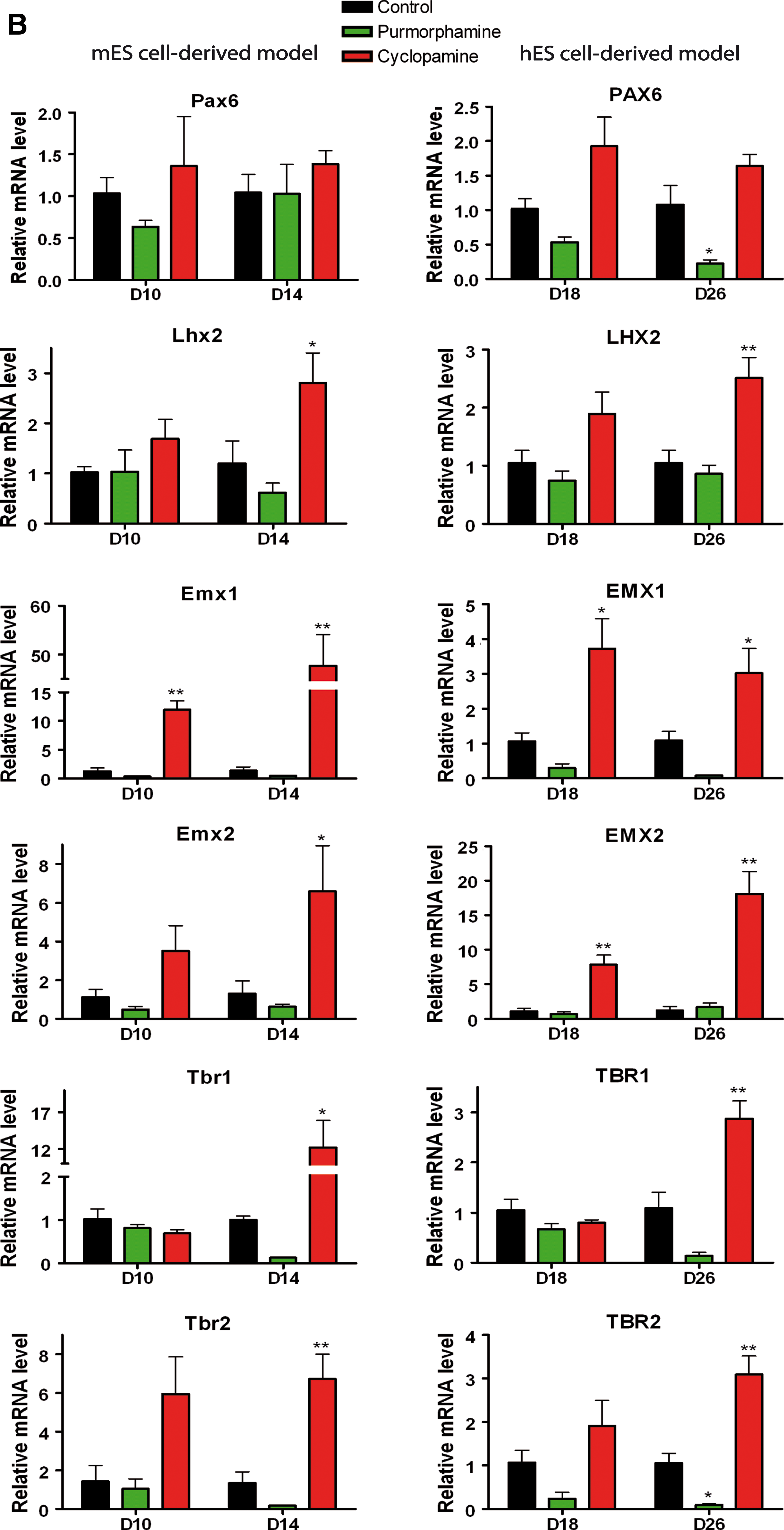
Pharmacological modulation of the Hh pathway differentially affects the expression of dorsal telencephalic transcription factors in mES and hES cell-derived telencephalic models.

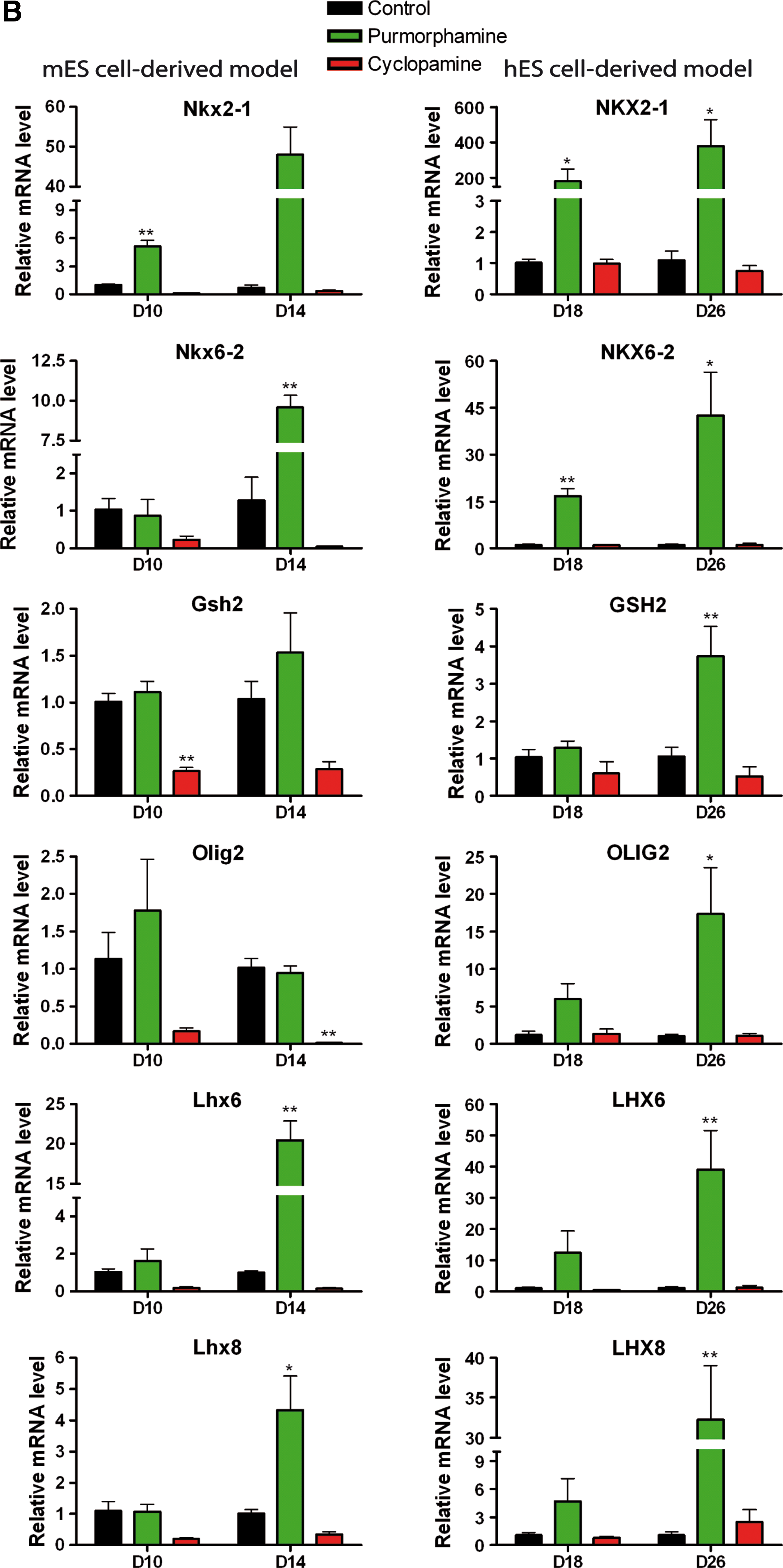
Pharmacological modulation of the Hh pathway differentially affects the expression of ventral telencephalic transcription factors in mES and hES cell derived telencephalic models.

Dorsal telencephalic markers
In the mouse dorsal embryonic telencephalon, Pax6, Emx1, Emx2, and Lhx2 are specifically expressed in progenitor domains, like the ventricular zone (VZ) and the subventricular zone (SVZ). Pax6 is mainly expressed in the neocortex, while Emx1 and Emx2 are medially expressed in the archicortex (later hippocampus) [38,43,52 –54]. Lhx2 is expressed in both regions [46]. Tbr2 and Tbr1 are expressed in the SVZ and mantle zone (MZ) in mouse embryonic cortex, corresponding to basal progenitors and further cortical plate cells, respectively [53,55].
In comparison with mouse cultures analyzed at D10 under control (untreated) conditions, the expression levels of Pax6, Lhx2, Emx1, Emx2, Lhx2, Tbr1, and Tbr2 in the dorsal embryonic telencephalon (cortex) were significantly higher while the levels in the ventral telencephalon were not significantly different (Fig. 3A). This confirms previous results indicating a default ventral telencephalic patterning in mES cell-derivatives [5,39]. In human control cultures, the dorsal markers PAX6, LHX2, EMX1, and EMX2 were expressed at high level in NE cells and their expression did not change over time (at D18 and D26), while TBR1 and TBR2 were expressed at later time-points and their expression increased over time (Appendix Table A2). This provides further evidence for a default dorsal telencephalic patterning in hES cell-derived neural cultures [40].
Significant increases in mRNA expression were observed for Emx1 at D10 and D14 (∼50-fold), and for Lhx2, Emx2, Tbr1, and Tbr1 at D14 in cyclopamine treated mouse cultures (Fig. 3B). These results support previous findings that inhibition of the Hh pathway is critically required for the generation of dorsal telencephalic progenitors from mES cells [5].
For human cells, the effect of cyclopamine treatment reached significance for PAX6 at D18, EMX1/2 at D18 and D26, and LHX2, TBR1, and TBR2 at D26, although the levels of upregulation were generally low (only ∼2–3-fold), with the exception of EMX2 (∼20-fold) (Fig. 3B). Therefore, cyclopamine caused a more prominent increase in the expression of dorsal patterning markers in mouse progenitors compared to human progenitors. Our results also indicate a significant effect particularly for prospective hippocampal progenitors expressing Emx1 and Emx2.
For the dorsal markers in mouse cultures, differences in expression after purmorphamine treatment did not reach statistical significance. Purmorphamine treatment in the human cultures selectively reduced PAX6 and TBR2 expression at D26 compared with controls (Fig. 3B).
Ventral telencephalic markers
The ventral marker Nkx2.1 is expressed in the mouse embryonic ventral telencephalon in the progenitor domains throughout the MGE [43,56,57] whereas Gsh2 is expressed in the LGE and caudal ganglionic eminence (CGE) [43,56]. Nkx6.2 is expressed along the MGE/LGE sulcus and at high levels in the dorsal MGE (dMGE) [20,58]. Dlx2, Lhx6, Lhx8, and Isl1 are mainly expressed in the SVZ and MZ of the mouse embryonic ventral telencephalon, corresponding to the location of specified neurons [43,43,59 –63]. Lhx6 and Lhx8 have very similar expression patterns from the earliest stages of MGE development [16,40,59,64]). Additionally, Lhx6 is expressed in the dorsal telencephalon, corresponding to cortical interneurons originating from ventral telencephalon [61].
Ventral markers were highly expressed in mES cell-derived neural culture but their level of expression was in general lower than the expression in mouse ventral embryonic telencephalon (Fig. 4A). However, the abundant expression of these genes confirmed the default ventral telencephalic patterning in mES cell-derived cultures [5,39]. In human cultures ventral markers were expressed at very low levels and their expression did not change during spontaneous neural differentiation, consistent with a default dorsal telencephalic patterning in neural derivatives of hES cells in vitro (Appendix Table A2).
Compared with untreated cultures, cyclopamine caused a trend toward downregulation of ventral markers in mouse cultures, but significance was reached only for Gsh2 at D10 and Olig2 at D14 (Fig. 4B).
Purmorphamine had a dramatic effect on the ventral markers. In preliminary experiments, purmorphamine was used at the concentration of 1, 2, and 3 μM (Appendix Table A4) and in all the other experiments (Figs. 3 –6) at 2 μM. Nkx2.1 expression was significantly upregulated at D10 (∼8-fold) and D14 (∼50-fold). Nkx6.2 was also significantly upregulated by purmorphamine at D14 (∼10-fold) whereas Lxh6 and Lhx8 were upregulated at D14 (∼50-fold and 5-fold, respectively), subsequent to Nkx2.1 upregulation (Fig. 4B). In human cultures, purmorphamine significantly and massively upregulated the expression of NKX2.1 (∼200-fold at D18 and ∼400-fold at D26), NKX6.2 (∼20-fold at D18 and ∼40-fold at D26), OLIG2 (∼18-fold at D26), LHX6 and LHX8 (∼40-fold at D26). The expression of GSH2, DLX2 and ISL1 was also increased (∼4-fold ∼5-fold, and ∼7-fold, respectively, at D26) (Fig. 4B).

Pharmacological modulation of the Hh pathway differentially affects the expression of neuronal specification markers in the mES and hES cell-derived telencephalic models.
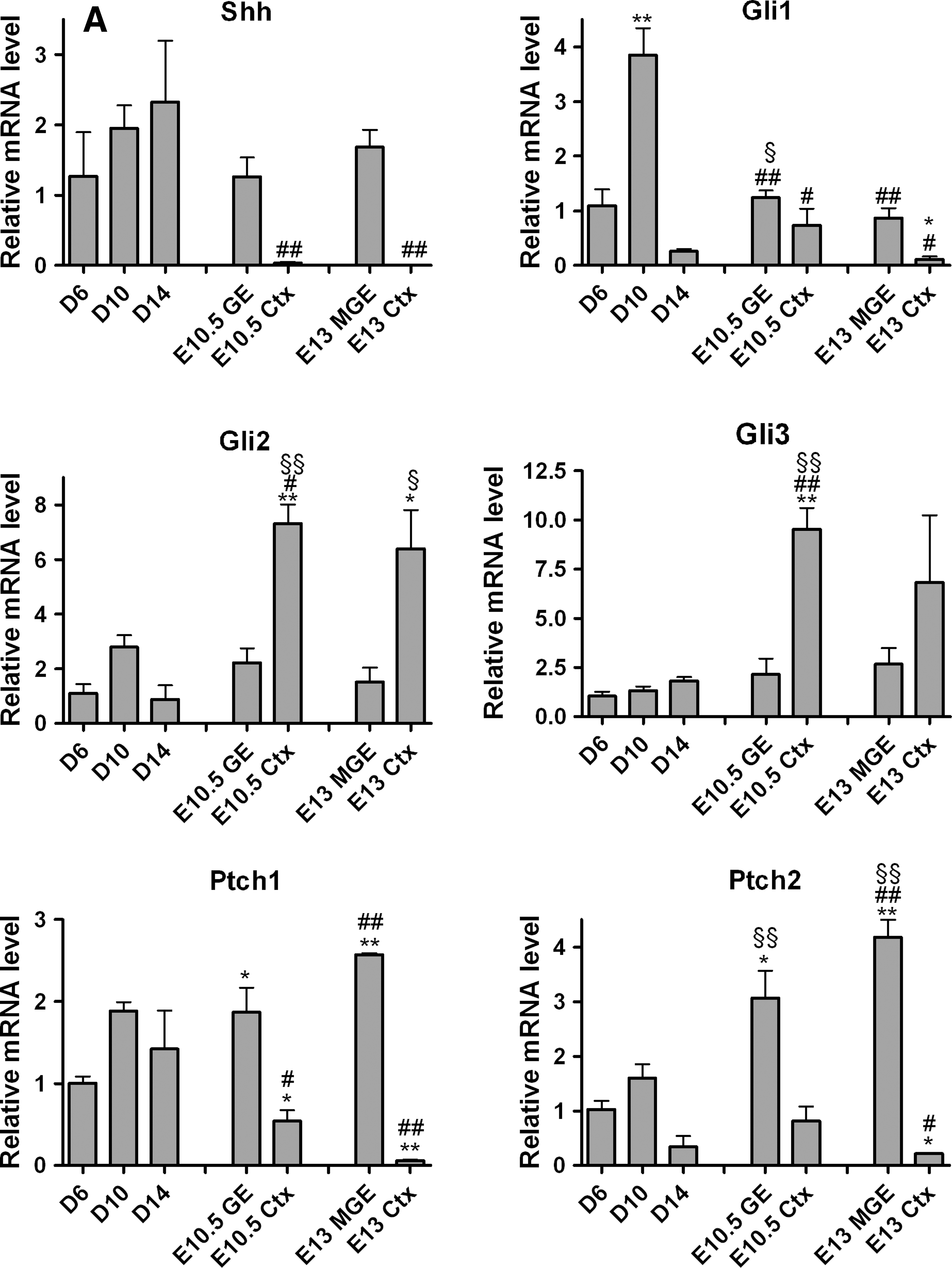
Pharmacological modulation of the Hh pathway via Smo differentially affects the expression of Gli effectors, Shh ligand, and Ptch receptors in mES and hES cell-derived telencephalic models.


Therefore, purmorphamine mainly had an effect on ventral patterning markers. The strongest effect was observed for the expression of MGE-related marker NKX2.1 in human telencephalic progenitors, which was confirmed by immunocytochemistry (Appendix Fig. A4). Nkx2.1 was expressed in 61.3%±2.3% of purmorphamine treated mouse cells versus 22.6%±6.3% in spontaneously differentiated cells and was not detected in cyclopamine treated cells at D10. NKX2.1 was expressed in 84.3%±7.3% of purmorphamine treated human cells and not detected in spontaneously differentiated cells or cyclopamine treated cells at D18. Therefore, human NE cells treated with purmorphamine were efficiently patterned into progenitors, the majority of which expressed NKX2-1.
Hence, during a time-window which enables telencephalic D/V patterning to occur, in vitro induction of Hh signaling by its agonist purmorphamine induced at first a MGE-like cell identity (expression of Nkx2.1, Lhx6, Lhx8) [57] and in addition specified MGE subgroups (such as the dMGE with high expression of Nkx6.2), in both mouse and human models, but more predominant in the latter.
Hh modulation differentially affects the expression of neurotransmitter and neurochemical phenotypic markers
We next analyzed the effects of the Hh modulation on neuronal differentiation and specification. Expression levels of the pan-neuronal markers Tau and Synaptophysin1 (Syn1), the neurotransmitter phenotype specific markers including the glutamatergic markers Vglut1 and Vglut2, the GABAergic markers Gad1, the cholinergic marker Vacht, and the GABAergic neurochemical markers somatostatin (Sst), parvalbumin (Pv), and calretinin (Calr) were analyzed in ES cell-derived cultures. Their expression was compared to the mouse embryonic telencephalon.
Modulation of the Hh pathway had no significant influence on the expression of the pan-neuronal marker Tau, indicating similar levels of neurogenesis in treated and control cultures (Fig. 5B).
Vglut1 was expressed at very low levels in mouse cultures at D10 and in the ventral telencephalon, but was significantly higher in the cortex (Fig. 5A). In human cultures mRNA for VGLUT1 was detected at D26 (Appendix Table A2). The treatment with cyclopamine upregulated the expression of Vglut1 in both mouse cells (∼250-fold) and human cells (∼3-fold), while purmorphamine treatment decreased expression in human cells at D26. Vglut2 expression did not significantly change in mouse cultures, but was upregulated by cyclopamine in human cultures at D26 (Fig. 5B).
For mouse cells, the temporal expression of Gad1 significantly increased between D10 and D14, reaching levels similar to those in mouse GE, while the level of Gad1 in the cortex was very low at the embryonic stages analyzed (Fig. 5A). Purmorphamine increased Gad1 expression (∼4-fold) at D14 in mouse and at D26 in human cells, whereas no effect was observed with cyclopamine (Fig. 5B).
Since most GABAergic neurons in the mouse telencephalon express Sst, Pv, or Calr, we analyzed expression of these neurochemical markers at the latest stage of our cell cultures and compared them with the values obtained from mouse embryonic telencephalon. Pv+ and Sst+ subgroups originate from the MGE, whereas the Calr+ subgroup mainly originates from the CGE [4,65,66]. Within the MGE, there is a bias toward Sst+ interneurons, derived from the dMGE, and Pv+ interneurons, from the ventral MGE (vMGE) [16,57,67]. Calr and Pv were expressed at low or undetectable levels in our cultures and in the mouse embryonic telencephalon at the developmental stages analyzed here (Appendix Tables A2 and A3). Sst was differentially expressed in the dorsal and ventral telencephalon (Fig. 5A). Its expression was also significantly modulated by treatment with purmorphamine in both mouse cultures (∼7-fold) at D14 and human cultures (∼3-fold) at D26 (Fig. 5B).
Similar low levels of Vacht were detected in mouse cultures at D10 and D14 and in the mouse embryonic telencephalon (Appendix Table A2). Expression of Vacht after purmorphamine and cyclopamine treatment was quantified at D14 for mouse cells and D26 for human cells. In mouse cells purmorphamine significantly upregulated Vacht expression (∼5-fold) compared with control conditions. For human cells, a similar trend was found, (∼4-fold), but the effect did not reach statistical significance (Fig. 5B).
Immunocytochemistry confirmed the presence of GABA+ and Vacht+ neurons after purmorphamine treatment in mES cell-derived cultures at D14, while such neurons were eventually detected in later time-points in hES cell-derived cultures (Appendix Fig. A5).
We therefore conclude that, although overall neuronal differentiation (determined by Tau expression) was not influenced by Hh modulation in our model, the neurotransmitter marker expression was. In particular, the expression of the GABAergic marker Gad1 was upregulated by purmorphamine treatment, together with the cholinergic marker Vacht, in both mouse and human cells. The ventral progenitors generated both GABAergic and cholinergic neurons, consistent with in vivo development. The highest effect of purmorphamine treatment was determined for the generation of a GABAergic subpopulation expressing Sst, corresponding to Sst+ cortical interneurons derived from dMGE in vivo.
Hh modulation differentially affects the expression of Hh signaling pathway
In an attempt to explain the species differences in the default D/V telencephalic patterning and also the differential reactivity to Hh modulation via Smo observed in mouse and human cultures, we determined expression levels of molecules belonging to the Hh signaling pathway and compared to their expression levels in the mouse embryonic telencephalon.
The central mechanism that determines neural progenitor competence to Hh signaling is the activity of the Gli family of transcriptional regulators–Gli1, Gli2, and Gli3 [68,69]. Gli1 is expressed at high levels in the progenitor domain of the dMGE [58,68] whereas Gli2 is highly expressed in the progenitor domain of the dorsal telencephalon (cortex), with a lower expression in the LGE. Gli3 is also highly expressed dorsally, in the progenitor domain, with lower expression in the LGE and MGE [43,68,70]. Finally, Shh is expressed within the ventral telencephalon, in the VZ of the preoptic region and in the MZ of the MGE [58,68].
Gli1 was expressed in the mES cell-derived NE cells (D6) at similar level as in the mouse MGE (Appendix Tables A2 and A3). Further, Gli1 was significantly upregulated in the mouse model at D10 and downregulated at D14 (Fig. 6A). Gli1 was expressed at high levels in the mouse embryonic ventral telencephalon and was less abundant in the dorsal telencephalon (Appendix Table A3). By contrast, GLI1 expression was not detected in human NE cells and was expressed at very low levels during their spontaneous differentiation (Appendix Table A2).
We found a low mRNA expression level for Gli2 in mouse NE cells and human NE cells. Similar findings were obtained for Gli3 in mouse NE cells, but we detected high levels in human NE cells (Appendix Table A2). These levels of expression were maintained in both mouse and human cells during spontaneous differentiation (Fig. 6A). Expression in mouse cultures was not significantly different from levels in the mouse ventral telencephalon, but was significantly lower than the levels detected in the dorsal telencephalon (∼8-fold) (Fig. 6A). Gli2 and Gli3 were expressed at higher levels in the dorsal than ventral embryonic telencephalon (Appendix Table A3).
We found a high mRNA expression level for ligand Shh, and for the receptor Ptch1 in mouse NE cells in vitro, and at later stages during spontaneous differentiation, while the expression level for the receptor Ptch2 was lower (Fig. 6A and Appendix Table A2). High levels of expression were determined for Shh, Ptch1, and Ptch2 in mouse GE, whereas the level of expression in cortex was very low (Fig. 6A and Appendix Table A3). By contrast, SHH mRNA expression was not detected in human NE cells and remained at very low levels during the subsequent differentiation steps; PTCH1 and PTCH2 were expressed at low and very low levels, respectively (Appendix Table A2). SHH was not detected by immunocytochemistry in hES cells and hES cell-derived neural populations during spontaneous differentiation, but was readily detected in mouse neural derivatives (Fig. 6C).These results confirm previous reports of different levels of endogenous Shh in mES and hES cell-derived telencephalic precursors [40].
We then analyzed the effect of the Smo activity modulation on molecules of the Hh pathway and on FGF family members. Compared with control cultures, Gli1 expression was significantly upregulated by purmorphamine in mouse cultures at D14 and GLI1 in human cultures at D18. Gli3 expression was upregulated by cyclopamine and downregulated by purmorphamine at D10 in mouse cultures. In human cells, GLI3 expression showed a trend toward downregulation by purmorphamine (Fig. 6B).
Compared with control cultures, Fgf8 expression was not significantly altered by purmorphamine or cyclopamine treatment in mouse and human models, but Fgf15 expression was upregulated (∼5-fold) by purmorphamine at D14 in mouse and FGF19 in human cells at D18 (∼5-fold) and at D26 (∼7-fold) (Appendix Fig. A3B).
Finally, we analyzed the differential expression of Shh itself. Compared with control cultures, mouse cells showed no significant change in Shh expression by purmorphamine treatment and only a trend of downregulation by cyclopamine (at D10). In contrast, purmorphamine caused a steep upregulation in human cells (∼15-fold at D18 and ∼40-fold at D26) (Fig. 6B). The Hh receptors Ptch1 and Ptch2 followed the same expression pattern as the Shh in both mouse and human cells. Purmorphamine upregulated the expression of Ptch1 (∼4-fold) and Ptch2 (∼15-fold) at D14, while cyclopamine downregulated their expression at D10 in mouse cells. In human cells, purmorphamine highly upregulated both PTCH1 and PTCH2 (∼15-fold at D18 and D26) (Fig. 6B).
The finding of a differential Shh expression after purmorphamine treatment in mouse and human models was confirmed by immunocytochemistry, where Shh expression was associated with both progenitor and neuronal phenotypes (see co-staining for Shh, Nestin, and Tau), in mES and hES cell-derived cultures (Fig. 6C).
We conclude that both mES and hES cell-derived telencephalic cells responded to Hh modulation by the small molecules cyclopamine and purmorphamine via Smo receptor. The observed changes are consistent with documented processes occurring during in vivo development. Marker gene expression levels reached similar levels in related structures in vitro and in vivo.
Discussion
Understanding the development of specific neural subtypes provides an essential framework for the design of rational approaches regarding the in vitro differentiation of pluripotent stem cells for in vitro studies and cell replacement therapies. Of particular interest, albeit a methodological challenge, is to generate specifically the types of neurons that build-up the telencephalon. In the absence of functional evidence regarding the in vivo human neurogenesis, it is necessary to extrapolate findings obtained in mouse experiments in vivo and in vitro to human cell cultures. We have recently proposed a comparative model of mES and hES cell differentiation following discrete steps of early telencephalic development: neural induction, neural patterning, and neuronal specification. Further, we discussed specific milestones between these steps that should be achieved before the next step of the protocol to be initiated [71]. In the present study we have described in detail a cell culture system which is based on this developmental approach and outlined how we established the alignment of developmental stages for cells from the 2 species in culture. In addition, we developed a specific profile of marker genes, which was derived from in vivo studies. The progress of human and mouse cells through the 3 differentiation steps was referenced to a parallel ex vivo analysis of mouse embryonic telencephalon. To exemplify the value of this approach we have analyzed in greater detail a single process, the step of D/V telencephalic patterning.
Alignment of mouse and human cell cultures
PAX6 is uniformly expressed in the single-layered NE cells of human embryos at around E18, during transition from the neural plate to the neural tube. At this point PAX6 determines the loss of pluripotency in the NE cells by differentially targeting pluripotency and NE genes [38]. Sox1 is the earliest marker for the single-layered NE cells in mouse embryo, being expressed at around E8 when the neural plate begins to form [38]. mES cell-derived NE cells uniformly express Sox1 around D6 in our cultures, whereas hES cell-derived NE cells uniformly express PAX6 around D10, in line with previous similar protocols [5,9,40,50,72 –74]. Based on the earliest and uniform expression in the NE cells in vivo and in vitro of Sox1 in mouse and PAX6 in human, we consider that the D6 condition for our mouse model corresponds to E8-8.5 of mouse development, whereas D10 in the human model is comparable to E18 in human embryos.
A similar strategy was applied to align the D/V patterning stages between human and mouse cells. In the human embryonic telencephalon, the restricted expression of PAX6 to the dorsal region at E26 [40] can be taken as an indication for the timing of D/V patterning. In mouse telencephalon, the timing of D/V patterning is well documented. In our culture we defined a neurogenic time-window between D6-D10 in mouse and D10-D18 in human model. The expression level of markers for progenitor and neuronal cells was not substantially different at E10.5–11 in mouse telencephalon compared to D10 in mouse, and D18 in human cultures. Importantly, glial markers were absent at these time-points.
This alignment based on days in vitro turned out to be a useful indicator in our cultures. However, we aim to replace a fixed scheme of days in vitro with a more flexible scheme, considering the differences between individual lines of pluripotent stem cells and the variability of cell behavior in culture. Future treatment decisions will be taken on the basis of a quantitative profile, consisting of a complex battery of developmental markers, at first described in the present study. Importantly, expression levels of these markers in our cultures have been compared to in vivo expression levels. For example, one milestone that has to be reached after neural induction and before the D/V patterning treatment to be started consists of defined threshold expression values not only for PAX6 or Sox1, but in addition for Ncad, Nestin, and Sox2. At the same time, other markers should be expressed below certain levels, including markers for endoderm and mesoderm and markers for pluripotency or for more differentiated stages of neural development (neuronal or glial).
Comparative gene expression analysis in vivo and in vitro
The present format of our model system, which is based on days in vitro, complemented with an analysis of in vivo mouse telencephalon development, permitted a careful comparison of the expression of TFs involved in A/P and D/V patterning and for signaling molecules involved in telencephalic neurogenesis.
Early A/P patterning
Consistent with previously published protocols for either mES or hES cell neural differentiation and studies of in vivo neural development, NE cells developed in our cultures exhibit an anterior phenotype, expressing telencephalic (FoxG1) and forebrain specific TFs (Otx1, Otx2, Six3, and Lhx2), while more posterior TF are lacking [3,5,9,36,40,50,74,75]. Our study complements the previously published data by showing that this default early telencephalic phenotype, which develops in chemically defined conditions and without addition of morphogens, is also characterized by the endogenous expression of specific signaling molecules known to be expressed in the early telencephalon in vivo. In particular, we detected a high endogenous expression of Fgf8 and Fgf15, and of Dkk1 and noggin. This finding is relevant since Dkk1 and noggin have been previously used as additives in ES cell derived telencephalic cell cultures. Since Dkk1 and noggin are endogenously expressed in an early peak in mouse and human NE cells in vivo and in vitro [51], exogenously added Dkk1 and noggin appear to be dispensable for early stages of telencephalic differentiation in vitro.
D/V patterning
The Hh pathway is of essential importance for D/V patterning in many regions of the CNS, including the telencephalon [10,11,18]. A large number of previously published ES cell differentiation protocols employed Hh pathway modulation via addition of Shh ligand or alternatively of its agonist/antagonists [3,5,9,40,50,74,76,77]. Gaspard et al. showed that the majority of neural progenitors derived from mES cells developed a ventral phenotype and expressed endogenous Shh [5]. hES cell-derived telencephalic progenitors, when cultured without morphogens, displayed dorsal telencephalic traits, which was correlated with a low expression of endogenous SHH [40]. Our study confirmed these findings in a direct comparison between the species.
Blocking Hh signaling with the Smo antagonist cyclopamine efficiently converted mouse ventral telencephalic precursors to dorsal progenitors, consistent with previous results [5,40,50]. This effect of cyclopamine was correlated with a strong and significant increase in the expression of dorsal markers in the mouse cultures, while the same treatment had little effect on human cultures.
Danjo et al. recently reported that, in mouse cultures, moderate concentrations of Shh or Smo agonist caused generation of LGE-like progenitors, whereas at higher concentrations also MGE-like progenitors develop [74], while Maroof et al. reported the generation of a uniform population of Nkx2.1+, MGE-like mouse progenitors that further differentiated into Lhx6+ neurons [77].
With regard to human ventral progenitors, Aubry et al. reported the generation of LGE-like derived progenitors and neurons from hES cells treated with a combination of SHH, Dkk1, and the neurotrophin BDNF [73]. A recent study, in which hES cells were differentiated with a protocol similar to the one described here, showed that activation of SHH and/or inhibition of WNT permits specification of ventral telencephalic progenitors [40]. Shh, when added at high doses, almost completely eliminated the dorsal marker expressions, while NKX2.1 expression was increased. Supplying Shh at a lower concentration resulted in the differentiation of both LGE and MGE-like progenitors. WNT inhibition by Dkk1 or a higher concentration of Shh caused the generation of a ventral population of NKX2.1-expressing MGE progenitors [40]. This is in concordance with our results. MGE-like progenitors, expressing Nkx2.1, developed in both mouse and human cultures after purmorphamine treatment, while LGE markers and Olig2 marker were only moderately increased. The dramatic upregulation of Nkx6.2, a subregional marker of MGE, expressed in its dorsal part, is in agreement with recently published findings [20].
Neuronal specification
In our models we found evidence for specification of dorsal phenotypes, since the glutamatergic markers Vglut1 and Vglut2 had been expressed together with the early cortical specification markers Tbr1 and Tbr2 [55]. We complemented the temporal expression analysis of these markers in vitro with quantification of their expression in the developing mouse embryonic telencephalon in vivo. Cyclopamine upregulated the expression of Vglut1, Tbr1, and Tbr2 in both mouse and human cultures, and VGLUT2 was upregulated in human cultures; however, the effect of cyclopamine in human cultures was modest.
Our direct comparative analysis of dorsal telencephalic neuronal specification in mouse and human cultures confirmed previously reported data on the default dorsal specification in hES cell-derived cultures [40]. Further, a different efficiency of cyclopamine treatment was observed in the 2 species. This small molecule efficiently induced patterning of the dorsal progenitors in mouse, as previously reported [5], while it exerted modest effects in the human model.
In ventral telencephalic neurogenesis in vivo, Nkx2.1+ cells give rise to Lhx6, Lhx8, and Isl1 expressing neurons that subsequently acquire GABAergic and cholinergic neurotransmitter markers [56,59 –62,64,65,78]. The various subclasses of GABAergic neurons are mainly Lhx6+ interneurons, but also Isl1+ GABAergic projection neurons. The telencephalic cholinergic neurons express Lhx8 and Isl1 [79].
Previous in vitro studies demonstrated the efficient generation of neurons with the properties of cortical and striatal GABAergic interneurons from Nkx2.1+ progenitors in vitro [74,77]. Following treatment with purmorphamine for both mouse and human cells, we found a robust increase in expression of MGE-related neuronal genes Lhx6, Lhx8, and Isl1. These genes are direct targets of Nkx2.1 activity [20,79]. Similar high expression of Lhx6 was previously reported in mouse cell cultures treated with Shh [77].The expression of glutamate decarboxylase 65/67 (GAD 65/67 or Gad1) and Vacht expression was confirmed by qPCR and in later time-points by immunocytochemistry.
In vivo, the MGE generates early projection neurons from its most ventral aspect, while more dorsal domains, at later time-points, give rise to subpopulations of interneurons [20].
During neurogenesis, continued Shh signaling is required to maintain Nkx2.1 expression in MGE progenitors, and Nkx2.1 is required for the specification of Sst+ and Pv+ interneurons [20,65,68,80]. Lhx6 is also required for the specification of Sst+ and Pv+ subgroups [61,80]. Exposing vMGE progenitors to exogenous Shh in slice culture produces a shift toward Sst+ fate, while suppressing the generation of Pv+ interneurons [20].
This set of in vivo/ex vivo data is in agreement with our results obtained in vitro. Purmorphamine, at least in the time-window of differentiation in our cultures, predominantly caused the generation of a GABAergic population expressing Sst. Hence, interneuron diversity appeared to be influenced by Hh signaling within the ventral telencephalic progenitors in the ES cell-derived culture models as well.
In summary, our results demonstrate that human glutamatergic neurons are generated in the absence of morphogens, whereas ventral telencephalic GABAergic and cholinergic neurons are induced by the Hh agonist purmorphamine. Purmorphamine reproducibly and robustly controls MGE-related specific patterning and differentiation of subpopulations during specific time-frames of in vitro differentiation. The efficient generation of NKX2.1+ human progenitors in vitro is a significant step toward the homogeneous generation of specific types of human telencephalic cells.
Expression of Gli TFs correlates with differential responses to Hh modulation in ventral telencephalic progenitors
The 3 Gli genes are expressed in distinct and partially overlapping spatial domains in the ventral telencephalon [43,68]. Consistent with this broad expression of Gli2 and Gli3, gain-of-function studies have shown that most regions of the telencephalon are able to respond to Hh signaling [20,67 –70]. Gli3 is an essential factor for Hh modulation in telencephalon and is the first Gli member to be expressed in telencephalic anlagen [43,70]. A truncated form of Gli3 represses Shh activity [43]. Thus, Hh signaling in the ventral telencephalon might simply counteract Gli3 repressor activity. Detailed analyses of gene expression indicates that the response to Hh signaling during telencephalic development is region specific and that specific targets of Hh signaling, including Gli1 and Nkx6.2, are only expressed in the most dMGE [16,20,67].
We found Gli TFs to be differentially expressed in mouse and human NE cells. Together with the differential level of endogenous Shh expression, these species differences might explain the default dorsal phenotype of human telencephalic precursors and the default ventral phenotype of mouse telencephalic progenitors.
The enriched expression of Gli1 and Nkx6.2 in the dMGE suggests that Shh signaling is enhanced in this region relative to the vMGE [43,81], and that this difference correlates with the specification of the Sst+subgroup of GABAergic neurons. Hh signaling modulation in our cultures from both species also modulates patterning consistent with the intra-MGE patterning in vivo and has an influence on specification of Pv+ versus Sst+ interneurons. Since different interneuron subtypes are generated at different stages of development in vivo [65,68], both the level of Hh signaling and its duration are likely to be critical [68,82,83]. Nkx2.1+ progenitors respond to high levels of Shh signaling primarily by generating Sst+ interneurons [65,68], which is in line with previous results in mES cell-derived cultures [74] and our results. Additionally, this aspect has been shown for the first time here in a hES cell-derived model.
FGF signaling is another important factor patterning and specification of telencephalic neurons. FGF8 promotes proliferation and survival of telencephalic progenitor cells, and specifies anterior telencephalic fate through positively regulating expression of several TFs, including Foxg1 and Nkx2.1 [84,85]. In the embryonic telencephalon, Hh signaling acts both upstream and downstream of FGF signaling via Fgf15 and Fgf8. Fgf8 is also upstream of Shh and regulates Shh induction [85]. Shh itself promotes Fgf15 expression and maintains Fgf8 expression [86]. Fgf8 represses caudal telencephalic fate through reducing the expression of the CGE marker CoupTF1 or the caudo-dorsal marker Emx2, whereas Fgf15 activates the caudal fate. Caudally, Fgf15 is also implicated in the specification of GABAergic neurons [68,84,85].
In our study Fgf15 and Fgf8 genes responded to Hh modulation in vitro, in accordance with their functions in mouse forebrain development. Purmorphamine upregulated Fgf15/FGF19 in our cell cultures, providing novel clues for the FGF signaling in human telencephalic development.
Finally, we have examined how the Shh expression itself is modulated by Smo agonist or antagonist treatment. We report for the first time that Shh expression is regulated in a very dynamic way by Hh pathway modulation, both in mES and hES cell-derived models. In particular, a very robust upregulation of SHH by purmorphamine was observed in the human model, where SHH was not expressed in untreated controls (Fig. 6B). The same expression pattern was for the first time documented here for the receptors Ptch1 and Ptch2 at the telencephalic level in vitro. Shh was expressed in both progenitor cells and neurons in our cultures (Fig. 6C, D). This might be explained by the noncell autonomous mechanisms recently described in the mouse embryonic telencephalon, where both Lhx8 and Lhx6 genes controlled the expression of Shh in the MZ of the MGE, corresponding to early-born neurons [68,87]. Thus, Lhx6 and Lhx8 appear to regulate MGE development by promoting Shh expression in MGE neurons, which, in turn, promotes the developmental program of the dMGE.
Conclusion
Our results show that similar to mES cells, hES cells differentiate into region specific progenitors, following the same developmental principles that have been identified by studying mouse CNS development. Together with previous findings, our data support that human neural progenitors in culture develop a reverse default D/V phenotype compared with mouse. However, early human NE cells can be efficiently differentiated into dorsal and ventral telencephalic progenitors via modulating similar molecular mechanism as described for other vertebrates. The activation of Hh signaling via Smo with purmorphamine converts the primitive dorsal telencephalic precursors to ventral progenitors. These progenitors then differentiate into neuronal subtypes including GABAergic and cholinergic neurons.
Therefore, mES and hES cell-derived models, directly compared in parallel experiments and temporally aligned to in vivo telencephalic development, offer a platform for testing the effect of morphogens, growth factors, and pharmacological substances for the generation of specific neuronal subtypes.
Telencephalic progenitors and neurons generated in vitro from human pluripotent cells provide a unique paradigm to study the human telencephalic development.
Even more importantly, the application of optimized telencephalic differentiation protocols to IPS cell cultures [1,2] derived from patients with neurodegenerative or neurogenetic diseases, will provide unique new opportunities to develop in vitro models of human ailments of the telencephalon such as Alzheimer's disease, Huntington's disease, epilepsy, and neuropsychiatric disorders. These models, based on human neurons in culture, will critically complement existing animal models, which do not fully reflect important features specific of the normal and pathological human brain.
Footnotes
Acknowledgments
This work was supported by CNCSIS, Romania (Project IDEI 626/2007), Innsbruck Medical University, Austria (IFTZ grant 2008), SPIN FWF W1206-B05 doctoral program, Dementia Foundation, Sweden, and Gun och Bertil Stohnes Stiftelse, Sweden. The authors acknowledge Prof. Outi Hovatta for kindly providing the human embryonic stem cells lines; Dr. Alina Codita and Dr. Galina Apostolova for critically reading the article; and Theodor Regalia, Monika Waldner, and Tanja Massimo for technical support.
Author Disclosure Statement
No competing financial interests exist.
Appendix
| D14 mES cell-derived model | |||
|---|---|---|---|
| Purm 1 μM | Purm 2 μM | Purm 3 μM | |
| gene/ΔCt | |||
| Calr | 6.3±0.2 | 5.8±0.4 | 7.2±0.2 |
| Dcx | 2.9±0.1 | 4.0±0.3 | 3.9±0.0 |
| Dkk1 | NE | 11.3±0.2 | NE |
| Dlx2 | 5.1±0.3 | 4.6±0.3 | 6.9±0.1 |
| Emx1 | 12.7±0.5 | NE | NE |
| Emx2 | 9.1±0.2 | 8.8±0.3 | 9.0±0.1 |
| Fgf15 | 7.2±0.1 | 8.0±0.5 | 8.7±0.3 |
| Fgf8 | 11.0±0.2 | 9.8±0.5 | 11.1±0.2 |
| Foxg1 | 4.7±0.1 | 4.5±0.4 | 6.0±0.1 |
| Gad1 | 2.4±0.2 | 2.2±0.3 | 4.8±0.1 |
| Gfap | −1.0±0.4 | −2.0±0.3 | 0.4±0.2 |
| Gli1 | 7.2±0.1 | 5.9±0.1 | 7.6±0.0 |
| Gli2 | 8.1±0.1 | 8.3±0.2 | 8.4±0.0 |
| Gli3 | 12.8±0.2 | 12.3±0.2 | 12.2±0.1 |
| Gsh2 | 10.1±0.1 | 10.8±0.5 | 11.8±0.3 |
| Ncad | 2.4±0.2 | 2.0±0.4 | 3.2±0.1 |
| Nkx2-1 | 7.3±0.3 | 7.0±0.4 | 9.4±0.2 |
| Nkx6-2 | 5.1±0.2 | 5.0±0.2 | 5.3±0.1 |
| Nog | 6.8±0.3 | 6.5±0.2 | 5.7±0.3 |
| Olig2 | 0.8±0.3 | 1.3±0.1 | 1.5±0.2 |
| Osp | 2.0±0.1 | 2.0±0.1 | 2.1±0.2 |
| Shh | 5.3±0.2 | 4.5±0.2 | 5.9±0.4 |
| Sst | 3.9±0.1 | 3.8±0.5 | 5.9±0.3 |
| Tau | 2.8±0.1 | 2.6±0.3 | 4.2±0.2 |
| Tbr1 | 12.6±0.3 | 12.1±0.1 | 11.1±0.2 |
| vGlut1 | 11.5±0.5 | 12.2±0.4 | NE |
| vGlut2 | 6.4±0.4 | 7.1±0.0 | 7.5±0.1 |
Data are expressed as ΔCt±SEM; n=3. P>0.05 for all telencephalic marks gene expression for purmorphamine 1 and 3 μM treatment as compared with purmorphamine 2 μM treatment by Student's t-test.
NE, not expressed.