
Abstract
Junctional adhesion molecule-C (JAM-C) is an adhesive cell surface protein expressed in various cell types. JAM-C localizes to the apically localized tight junctions (TJs) between contacting endothelial and epithelial cells, where it contributes to cell–cell adhesions. Just as those epithelial cells, also neural stem cells are highly polarized along their apical–basal axis. The defining feature of all stem cells, including neural stem cells (NSCs) is their ability to self renew. This self-renewal depends on the tight control of symmetric and asymmetric cell divisions. In NSCs, the decision whether a division is symmetric or asymmetric largely depends on the distribution of the apical membrane and cell fate determinants on the basal pole of the cell. In this study we demonstrate that JAM-C is expressed on neural progenitor cells and neural stem cells in the embryonic as well as the adult mouse brain. Furthermore, we demonstrate that in vivo JAM-C shows enrichment at the apical surface and therefore is asymmetrically distributed during cell divisions. These results define JAM-C as a novel surface marker for neural stem cells.
Introduction
T
Interestingly, murine JAM-C is highly expressed on hematopoietic stem cells in the bone marrow [14]. This observation as well as the described function of another JAM-family member (JAM-A) as a marker for adult hematopoietic stem cells [14] and our previous report showing the expression of JAM-A on neural NG2-glia cells [15] prompted us to investigate JAM-C expression in the developing and adult mouse brain. Furthermore, an expression of JAM-C at the mRNA level in the stem cell compartments of the adult brain has been suggested previously [16].
Neural stem cells (NSCs) are self-renewing multipotent cells present in the developing and adult mammalian central nervous system (CNS) [17]. NSCs generate all the neurons and glia cells during embryonic brain development, but they are also responsible for the production of new neurons and glia cells in the adult mammalian brain [17,18]. Both in the embryo and in the adult brain NSCs exist in niches that are important for the maintenance of their stem cell characteristics [19]. In the embryonic brain, the best described neurogenic region is the ventricular zone (VZ) of the lateral ventricles. The neural progenitor cells in this region produce all neurons of the developing cortex [20]. In the adult brain, the main NSC-containing neurogenic region is the subventricular zone (SVZ) of the lateral ventricles. Neurons born in the adult SVZ migrate over a great distance through the rostral migratory stream and become granule neurons and periglomerular neurons in the olfactory bulb. Furthermore, adult neurogenesis also occurs in the dentate gyrus in the hippocampus [18].
The defining feature of all stem cells, including NSCs, is their ability to self renew. This self-renewal can occur by symmetric cell divisions which will generate two daughter cells with the same cell fate, thereby increasing the pool of stem cells. Self-renewal can also occur by asymmetric cell divisions, which generate one daughter cell that is identical to the mother cell and a second daughter cell that undergoes differentiation (eg, into a neuron). In NSCs, the decision whether a division is symmetric or asymmetric largely depends on the distribution of the apical membrane [21] and cell fate determinants on the basal pole of the cell [22, 23].
Here we show that JAM-C is expressed on neural progenitor cells and neural stem cells in the embryonic as well as the adult mouse brain. Furthermore, we demonstrate that in vivo JAM-C shows enrichment at the apical surface and therefore is asymmetrically distributed during cell divisions. Interestingly, this apical polarization is abrogated in in vitro cultures of NSCs. Thus, we provide evidence that JAM-C is a novel surface marker for neural stem cells and could be valuable for further investigations focusing on asymmetric cell divisions.
Materials and Methods
Materials
The following primary antibodies were used: anti-ZO-1 (Invitrogen; species: mouse, dilution: 1:600), anti-CD133 (Millipore; species: rat, dilution: 1:400), anti-tripartite motif-containing protein 32 (TRIM32) (Abnova; species: mouse, dilution: 1:400), anti-S100β (Sigma-Aldrich; species: mouse, dilution: 1:1,000). Rabbit polyclonal anti-JAM-C antibodies were generated by immunizing rabbits with the extracellular domain of murine JAM-C fused to the fragment crystallizable (Fc) part of human immunoglobulin G (IgG) as described previously for JAM-A [24]. The antibody was used in a dilution of 1:600. The antibodies were tested for specificity and found to be not cross-reactive with JAM-A and JAM-B (Supplementary Fig. S1; Supplementary Data are available online at
Mice
Mice were kept under standard conditions according to governmental rules and regulations. All stainings were repeated on sections from at least 3 different mice. The age of adult mice was between 60 and 90 days.
PCR and western blot from ventricular zone cells
To obtain samples of total RNA or protein from cells from the ventricular zone and subventricular zone, these regions were dissected from total brain vibratome sections. The tissue obtained by this procedure was used for either total RNA isolation according to the manufacturer's instructions (RNeasy Kit; Qiagen) or for production of protein lysates (described previously in reference 25). The RNA was used for the reverse transcription into cDNA. The obtained cDNA was used as template for polymerase chain reactions (PCRs) with specific primers directed against JAM-C (forward primer: TGCCTACAGATTCCAGAG; reverse primer: CTAATGCTACCCGTAGACGACAC). As positive control, a JAM-C coding plasmid was used. The protein lysates were used for western blotting with anti-JAM-C antibodies.
Immunohistochemistry of brain sections
Adult mouse brains were fixed via perfusion with 120 mM phosphate buffered saline (PBS), pH 7.4 followed by perfusion with 4% paraformaldehyde in PBS. After dissection the brains were post-fixed overnight with 4% paraformaldehyde in PBS. Sagittal sections (40μm) were prepared with a Vibratome (Leica). Sections were blocked with blocking buffer (100mM Tris buffer, 0.5% Triton X-100, 0.1% sodium azide, 0.1% sodium citrate, and 5% normal goat serum). Blocking was followed by incubation with primary and secondary antibodies, both diluted in the blocking solution. Images were collected by confocal microscopy using CEN software (Zeiss); image analysis was performed with the CEN software, Adobe Photoshop, and the IMAGE J software. To investigate the enrichment of JAM-C and TRIM32 on the apical or basal pole of the cell (Fig. 4) the fluorescence intensities for both proteins on the two poles were measured with IMAGE J. Then the value for the apical pole was divided by the value for the basal pole. A positive result indicated an apical enrichment, while a negative result indicated a basal enrichment.
All experiments involving mice have been conducted according to German Animal Welfare Act and have been approved by the responsible authorities (Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen). All stainings were performed in at least 2 brain sections from at least 3 different mice.
Immunocytochemistry of cells in culture
Chinese hamster ovary (CHO) cells were grown in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% heat-inactivated fetal calf serum (FCS), 2 mM
Neurospheres and homogenous cultures of neural stem cells were produced according to protocols published previously [26,27]. Self-maintaining neural stem cells were derived from the embryonic mouse brain (E12.5–E14.5) as described previously [27] and cultured in NS-A medium (Euroclone, Milan) supplemented with 10 ng/mL epidermal growth factor (EGF) (Peprotech), 10 ng/mL basic fibroblast growth factor (bFGF)-2 (Peprotech), 1×N2 supplement (Invitrogen),
The cells were processed for immunohistochemistry by fixation with 4% paraformaldehyde in 120 mM PBS, pH 7.4, permeabilized with 0.05% Triton X-100 in PBS, blocked with 10% goat serum in PBS and subjected to immunohistochemistry stainings with primary antibodies and secondary antibodies diluted in the blocking solution. Images were collected by confocal microscopy using ZEN software (Zeiss); image analysis was performed with the ZEN software, Adobe Photoshop, and the Image J software.
All stainings were performed independently in cells from at least 4 different cultures.
Results
JAM-C is expressed on the apical surface of neural progenitor cells in the developing mouse cortex
Neural stem cells are highly polarized cells with a clearly defined apical and basolateral membrane compartment [20]. JAM-C is expressed by a variety of cells where it shows a polarized distribution, such as at the tight junction (TJs) in epithelial and endothelial cells, the paranodal loops and Schmidt-Lanterman incisures in Schwann cells, and the spermatid–Sertoli cell interface in developing spermatids [7]. An expression on cells specific for the developing central nervous system has not been described so far. Therefore, in a first set of experiments we wanted to find out whether JAM-C is expressed on cells from the embryonic mouse brain.
To detect JAM-C we used an anti-JAM-C antibody that is specific for JAM-C and is not detecting other JAM-proteins like JAM-A or JAM-B (see Materials and Methods and Supplementary Fig. S1A and B). Stainings of the embryonic brain with this antibody revealed a strong staining only for those cells in the ventricular zone that are in direct contact with the lateral ventricle (Fig. 1A–C). Interestingly, these cells are known to be radial glia cells, the neural stem cells of the developing cortex. Furthermore, the obtained staining pattern appears to be very specific for the apical surface of those radial glia cells (Fig 1D). Finally, we also detected the expression of JAM-C specific mRNA in cells from the embryonic ventricular and subventricular zone (Supplementary Fig. S1C).
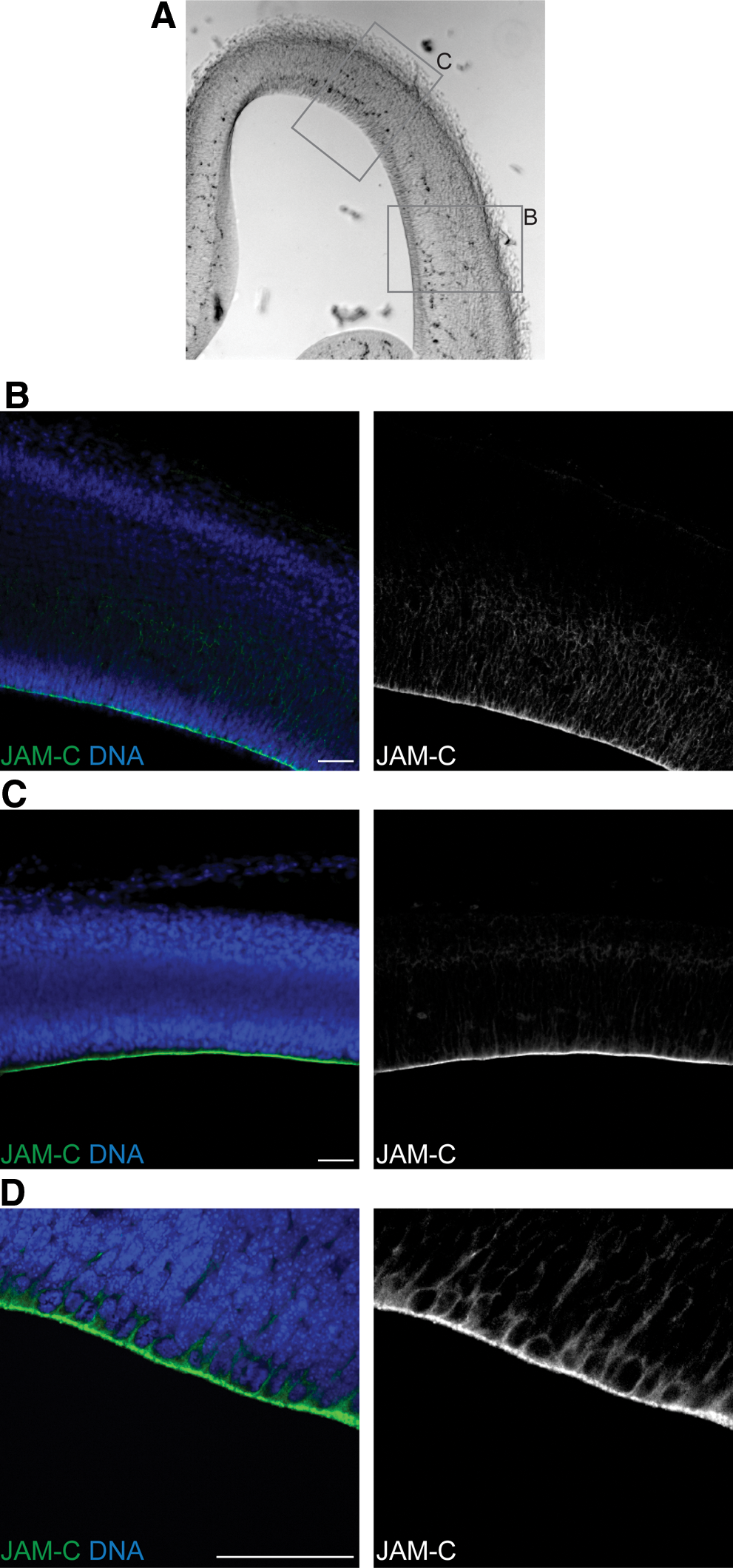
JAM-C is expressed in embryonic mouse brains. Bright field images
To verify the specific staining of JAM-C at the apical surface, in the next step we performed co-stainings with various markers. A co-staining of JAM-C with fluorophore-coupled phalloidin (to label actin) resulted in a partial co-labeling at the apical membrane (Fig. 2A). As expected from the non-exclusive localization of actin at the apical side (Fig. 2A), JAM-C staining showed only a partial overlap with actin at the apical pole of the cells. To further specify the localization of JAM-C we stained the brain sections with an antibody against ZO-1, a TJ specific scaffolding protein that directly interacts with JAM-C [6]. We found that ZO-1 is localized exclusively at the apical side of radial glia cells where it is specifically localized at the junctions (Fig. 2B). Moreover, JAM-C showed a significant co-localization with ZO-1 (Fig. 2B). As described previously [28], tight junction markers like Occludin and E-Cadherin are absent from radial glia cells. Therefore, co-stainings of these markers with JAM-C were not possible.
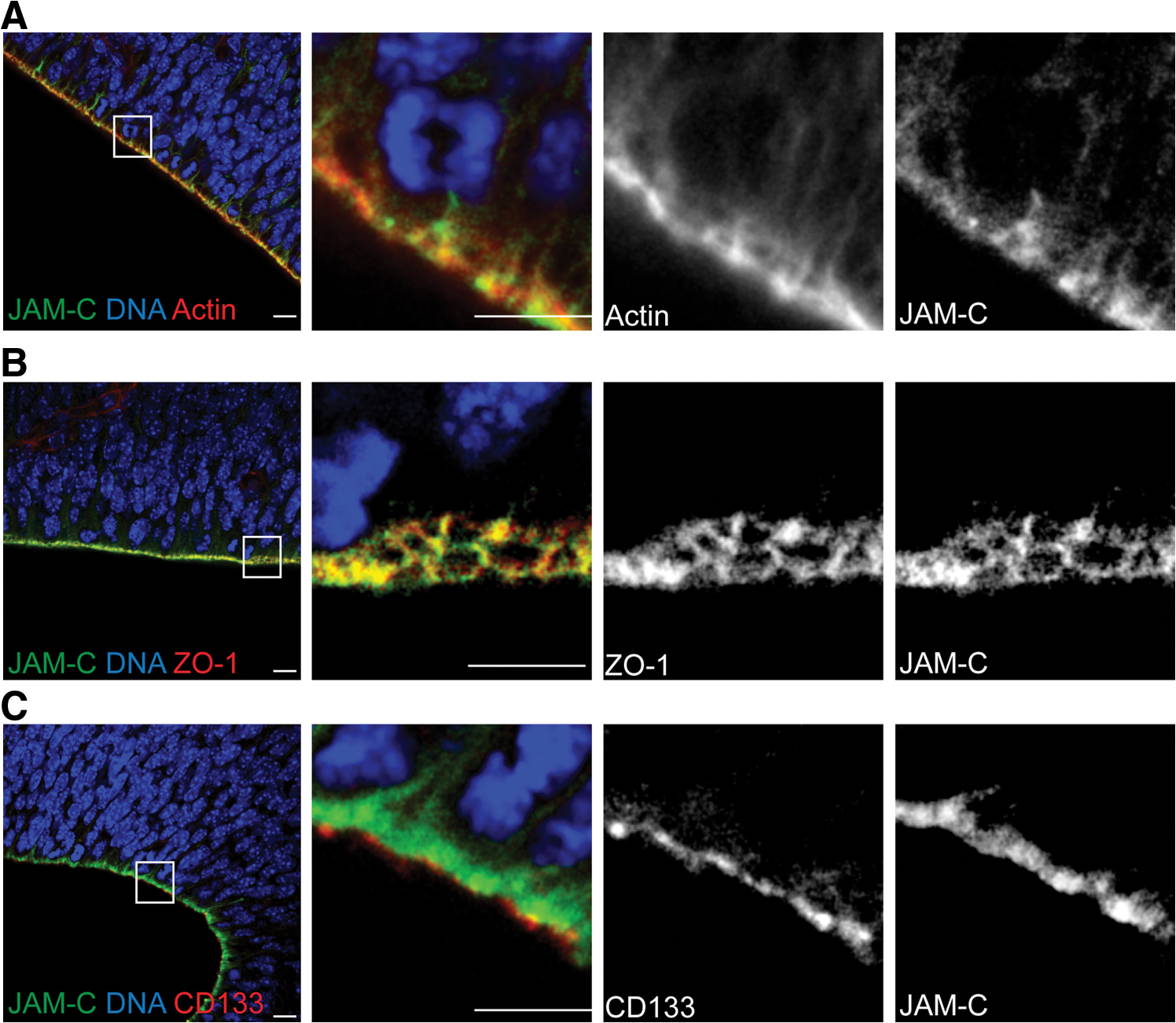
JAM-C is localized in the apical tight junctions.
Finally, we conducted a co-staining of JAM-C together with CD133. CD133 is specific for the apical surface of radial glia cells, where it is selectively associated with microvilli and microvilli-related structures [29]. However, although CD133 is expressed in the apical membrane, it is not part of the tight junctions. Consequently, in a co-staining of JAM-C and CD133 we always found the JAM-C staining below (seen from the apical surface) the CD133 staining (Fig. 2C).
Taking those observations together we conclude that, as shown previously for endothelial cells [30], on radial glia cells of the developing mouse brain, JAM-C is specifically localized in the tight junctions.
During asymmetric divisions JAM-C shows a polarized distribution
During brain development symmetric, proliferative, and asymmetric neurogenic divisions of radial glia cells co-exist. During symmetric proliferative divisions, the apical membrane is bisected by the cleavage plane, and therefore both daughter cells inherit one of the 2 adherens junction complexes. In contrast

The expression pattern of JAM-C in mitotic embryonic neural stem cells is discontinuous.
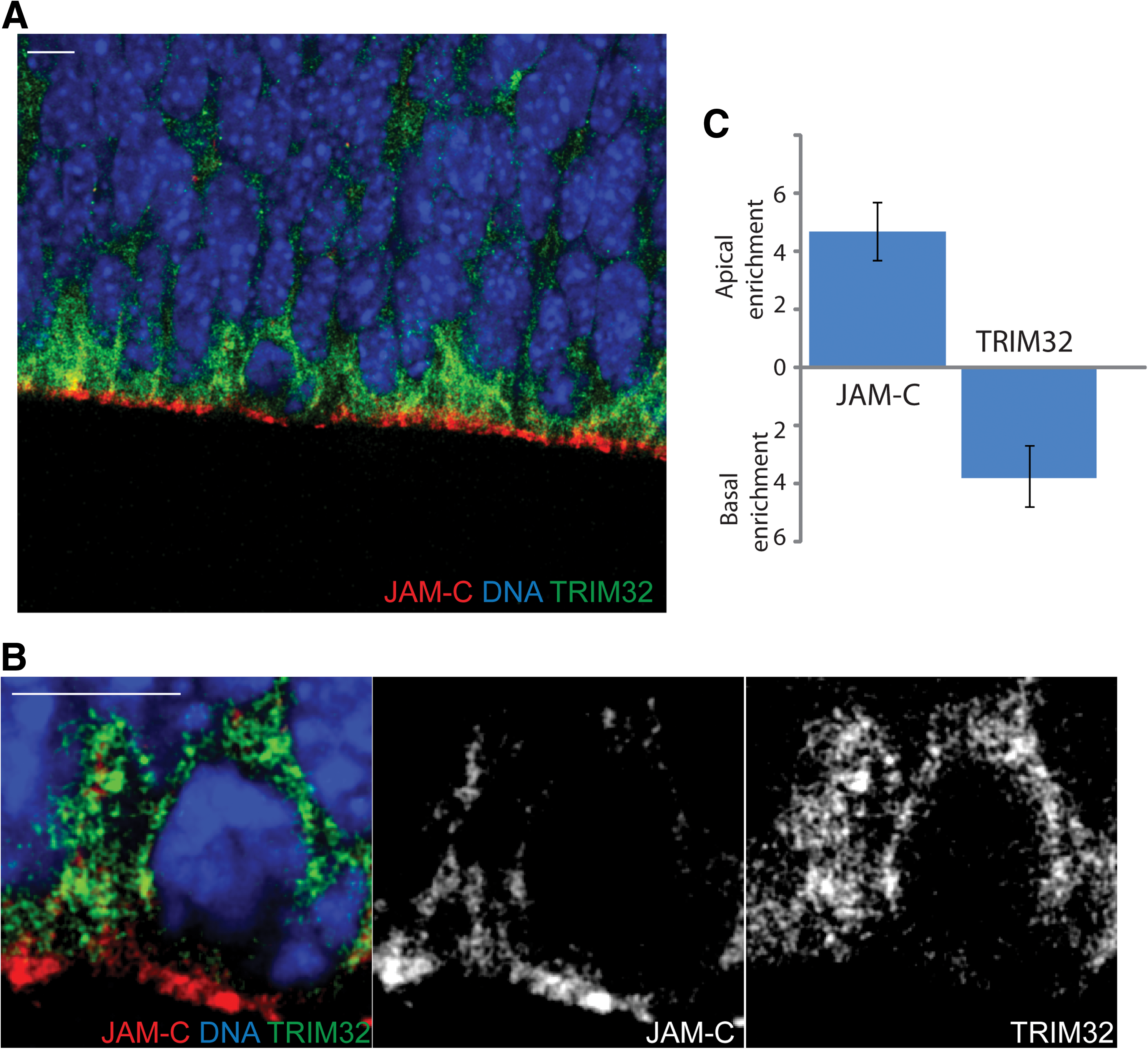
Tripartite motif-containing protein 32 (TRIM32) and junctional adhesion molecule-C (JAM-C) localize to opposite poles of mitotic cells.
So far, all of our observations are focused on the apical side of neural stem cells. However, after an asymmetric division, differentiation of one of the two daughter cells is induced not only by absence of the apical membrane components, but also by the inheritance of basally localized cell fate determinants. Previously we have shown that the basally localized protein TRIM32 is asymmetrically distributed during neurogenic divisions and is inducing neuronal differentiation in the inheriting daughter cell [23]. A co-staining of TRIM32 and JAM-C in radial glia cells revealed that the 2 proteins are localized at the opposite poles in mitotic cells. While TRIM32 is enriched at the basal pole, JAM-C showed a clear localization at the apical side (Fig. 1).
During embryonic neurogenesis, neurons are not only born at the apical surface; so-called basal progenitor cells in the deeper layers of the tissue are able to undergo mitosis and to produce neurons as well [31]. However, those basal progenitors do not show a typical apical-basal polarity. Consequently, they only weakly express JAM-C, and the JAM-C expression is not showing any subcellular polarization (Supplementary Fig. S2). We conclude that JAM-C, as part of the tight junctions, is strongly localized to the apical side of mitotic neural stem cells in the embryonic mouse brain. As a consequence of this apical localization, it is asymmetrically distributed during asymmetric cells divisions.
JAM-C is expressed in adult ependymal cells and Type-B neural stem cells
To this point our investigation focused on the neural stem cells of the embryonic mouse brain. However, the adult mammalian brain also contains neural stem cells. The most proliferative zone of the adult mouse brain is the SVZ, a region where neural stem and progenitor cells are present and where new neurons for the olfactory bulb are produced. It is still under debate which cell type in this region can function as real adult neural stem cells. Some publications argue for the ependymal cells as the neural stem cells of the adult SVZ [32], while others claim that a specialized subtype of astrocytes (the so-called Type-B cells) are the only real neural stem cells [33]. While ependymal cells are typical epithelial cells, the Type-B cells are more similar to the radial glia cells of the embryonic brain.
To first find out whether JAM-C is expressed on ependymal cells, we stained sagittal sections of the adult brain with our anti-JAM-C antibodies together with an antibody against S100β that serves as an ependymal cell marker. This labeling strategy revealed JAM-C localization at the apical side (facing the cerebrospinal fluid filled lateral ventricle) of the S100β positive ependymal cells (Fig. 5A).
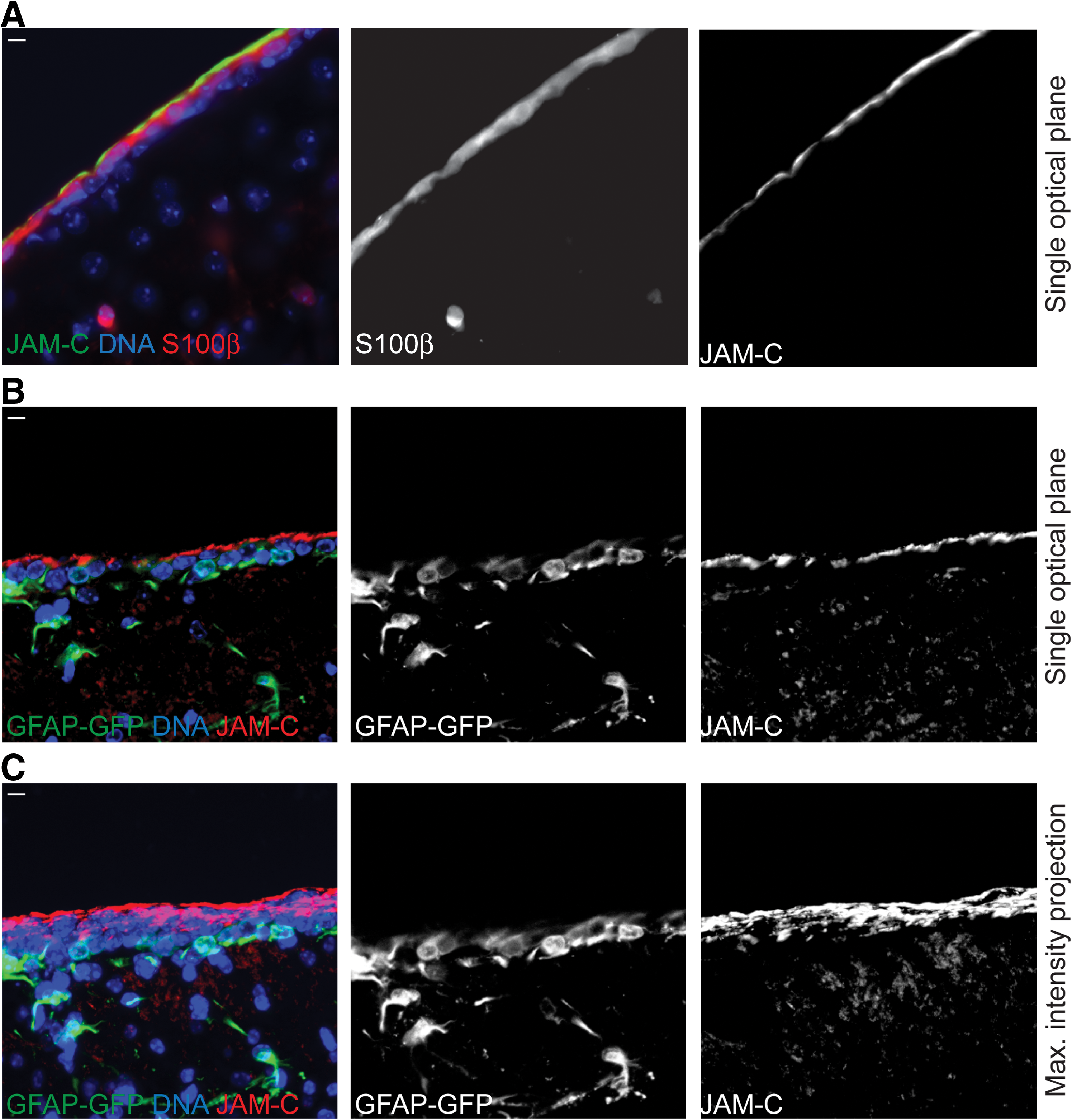
JAM-C is expressed at the apical surface of adult ependymal cells.
Next, to find out whether JAM-C is also expressed on Type-B neural stem cells, we used glial fibrillary acidic protein-green fluorescent protein (GFAP-GFP) knock-in mice. In those mice, Type-B neural stem cells are recognizable by GFP fluorescence [34]. However, in sagittal brain sections we hardly found any co-labeling between GFP and the JAM-C antibody staining (Fig. 5B and C). This lack of co-labeling could mean that JAM-C is not expressed on the apical membrane of those Type-B neural stem cells. However, the apical membrane of these cells is extremely tiny [35]. In fact, those Type-B cells extend only one single process through the ependymal cell layer to get in contact with the lateral ventricle, and only this single process is considered to have a real apical membrane [35]. As a consequence of this tissue organization, our approach of imaging sagittal sections is probably not suitable to detect the apical membrane of Type-B cells. Therefore, as an alternative we used a whole-mount staining approach that allowed us to directly look at the surface of the intact lateral ventricle (method described in reference 35). When we employed this imaging approach using the GFAP-GFP mice we were indeed able to see GFP positive processes (or probably bundles of processes) that project through the S100β positive ependymal cells and establish a direct surface to the lateral ventricle (Fig. 6). Strikingly, when we imaged the surface of those GFP-positive processes at high magnification, we indeed found a strong JAM-C co-labeling indicating that JAM-C localizes to the apical side of cellular processes that emanate from Type-B-cells and contact the lateral ventricle. In summary, in adult neural stem cells, just as in embryonic radial glia cells, JAM-C is specifically expressed at the apical side of the cell.

JAM-C is expressed at the apical surface of adult Type-B neural stem cells.
Additionally, we never found a significant JAM-C staining in glia cells (S100β or GFAP-GFP positive) within the brain parenchyma (Fig. 5 and Supplementary Fig. S3).
In neural stem cells in culture JAM-C shows an unpolarized subcellular distribution
Finally, we also were interested to see if neural stem cells from in vitro cultures display a polarized distribution of JAM-C. First, as described previously [36], we cultured these neural stem cells as spheres (called neurospheres). In those spheres, we imaged JAM-C by staining with the anti-JAM-C antibody together with a co-labeling with fluorophore-coupled phalloidin (to label actin). This staining revealed a concentration of JAM-C at the positions of cell–cell contacts, but no particular polarized subcellular distribution became evident (Fig. 7A and B). To limit the possibility of those neural stem cells to form cell–cell contacts we used a protocol for 2-dimensional, dissociated cultures of neural stem cells [27,37]. However, under those conditions also, the neural stem cells did not show any asymmetric distribution of JAM-C (Fig.7C). Furthermore, even during mitosis the distribution of JAM-C was rather uniform (Fig. 7C). These data show that the polarized distribution of JAM-C is lost when neural stem cells are dissected out of the tissue and cultured in vitro.
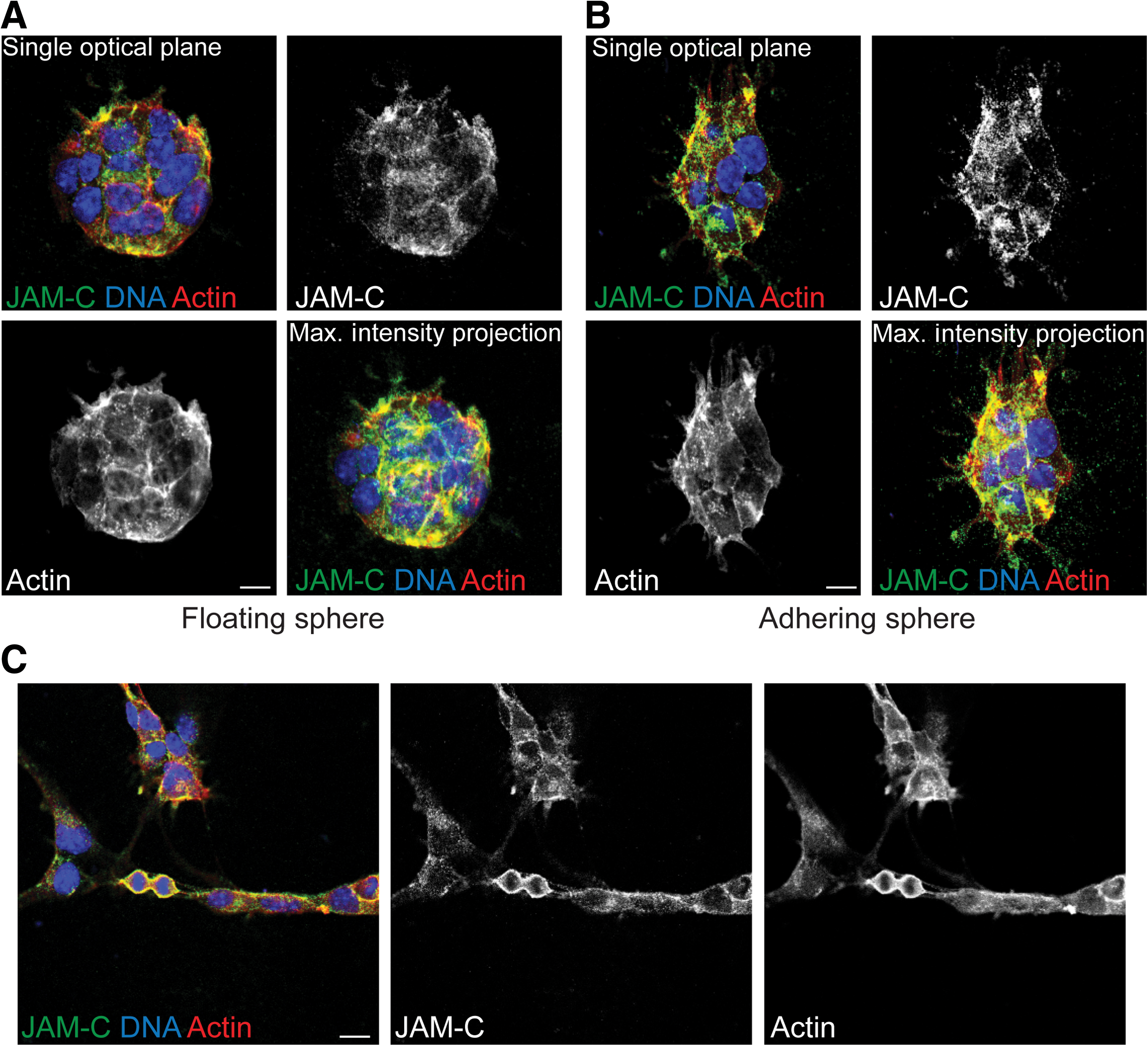
Neural stem cells in vitro are unpolarized.
Discussion
In this study we have analyzed the expression of the cell surface molecule JAM-C on mouse neural stem cells in the embryonic brain, the adult brain, and mouse neural stem cells in culture. Indeed, we found that all of these neural stem cells express JAM-C. Interestingly, only neural stem cells that reside in their in vivo niche, either in the embryonic ventricular zone or the adult subventricular zone, show a polarized apical enrichment of JAM-C. In contrast, neural stem cells that were grown in in vitro cell cultures express JAM-C homogenously on their surface. Especially for embryonic neural stem cells, the correlation between subcellular distribution of certain proteins and asymmetric cell division has been studied in great detail [38]. Because of the apical localization of JAM-C, it is very likely to be asymmetrically segregated into only one daughter cell during asymmetric (neurogenic) cell divisions. In good agreement with this we found JAM-C as a novel marker for the so-called “Cadherin hole” and the associated junctions [21]. This apical structure is a well-described asymmetrically segregated component of asymmetric cell divisions in the developing mouse brain [21]. Furthermore, because JAM-C is localized on the apical surface of mitotic neural stem cells, opposite to the basally enriched neuronal cell fate determinant TRIM32 (Fig. 1), upon asymmetric cell division it will be inherited by the daughter cell that maintain stem cell characteristics. Because JAM-C is a cell adhesion molecule, it is tempting to speculate that this inheritance in the daughter cell that maintains stem cell characteristics is important to hold this cell, via junctional interactions, in the stem cell compartment (ventricular zone) and to prevent its migration into the deeper cortical layers.
In the adult brain, it is still under debate which cell type in the subventricular zone possesses stem cell abilities. Both ependymal cells and Type-B astrocytes from this region have been claimed to be the real adult neural stem cells in the adult brain [32,33]. Regardless of this debate, in our study both cell types show a clear apical expression of JAM-C (Figs. 5 and 6). Ependymal cells were identified by their expression of the marker S100β and their characteristic localization at the lateral ventricle, while Type-B cells were identified by expression of GFAP (utilizing GFAP-GFP mice) and their extension of a single fiber toward the lateral ventricle. However, other glia cells in the adult brain also express S100β and/or GFAP (see Supplementary Fig. 3 and S100β positive cells in the SVZ in Fig. 5). Interestingly, in those non–stem cell glia cells we never found a significant expression of JAM-C. Those findings argue for our hypothesis that JAM-C is a NSC marker. Furthermore, this observation supports the concept that probably both cell types, ependymal cells and Type-B astrocytes, possess stem cell characteristics. However, one could argue that the JAM-C expression has nothing to do with stemness but is a simply consequence of the epithelial organization of those cells. Although this epithelial organization is certainly important for stem cell characteristics, this argument would fail to explain why neural stem cells, even in dissociated cultures, strongly express JAM-C (Fig. 7). Therefore, we conclude that JAM-C is a marker for NSCs and plays a role in the maintenance of their stemness.
In several cell types and tissues, the JAM-proteins are important for the integrity and tightness of epithelia [12,39,40]. Interestingly, a recent study in humans described the identification of a homozygous mutation in JAM-C in a large consanguineous family from the United Arab Emirates [13]. Among other symptoms, these patients also suffered from sub-ependymal calcification. Mutation of occludin, another tight junction protein, also leads to a calcification phenotype [41]. So far it has been assumed that the reason for the pathology in patients with mutant JAM-C or mutant occludin is a disturbed integrity of the cerebral vasculature. However, especially in the case of JAM-C this is not a very likely explanation, because JAM-C has been reported to be absent from vasculature of the adult mouse brain [3]. Based on the observed expression of JAM-C in the tight junctions in the embryonic VZ and the adult ependymal cell layer, an alternative explanation, where JAM-C is involved in the regulation of the permeability of the epithelium (VZ or ependymal layer) towards the cerebrospinal fluid, would be conceivable. However, it is important to note that the brains of JAM-C knockout mice are relatively unaffected [42]. Therefore, a comparison of the human and the mouse situation might be of limited value. In summary, here we describe for the first time the expression of JAM-C on neural stem cells of the embryonic and adult mouse brain as well as on cultured neural stem cells. Furthermore, we imply a role for JAM-C in asymmetric stem cell divisions.
Footnotes
Acknowledgments
The authors would like to thank Dr. Frank Kirchhoff (Göttingen) for the GFAP-GFP mice, and Dr. Ruth Beckervordersandforth-Bonk (Munich) for help with the whole-mount staining technique. T.P. is supported by a Marie Curie Fellowship. K.E.'s lab is supported by the German Research Foundation (DFG, EB160/4-1) and by the Innovative Medical Research fund of the Medical Faculty of the University of Münster (EB120323) and the Interdisciplinary Center for Clinical Research (IZKF) Münster (Eb2/028/09).
J.C.S.'s lab is supported by the German Research Foundation (DFG, Emmy Noether Program, SCHW1392/2-1 and SFB629), Kompetenznetzwerk Stammzellforschung NRW, Fonds der Chemischen Industrie, Fritz Thyssen Foundation (Az. 10.10.2.152), German-Israeli Foundation for Scientific Research and Development (G-2226-2034.1/2009), and the European Union Framework Program 7 (EU FP7) network EuroSyStem. Furthermore, this work was supported by the Innovative Medical Research fund of the University of Münster Medical School (SC120901 and SC411003) and the IZKF Münster (SchwJ3/001/11).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.