
Abstract
Epidermal growth factor (EGF) is a mitogen widely used when culturing adult neural stem cells in vitro. Although proliferative effects can also be observed in vivo, intracerebroventricular infusion of EGF has been found to counteract neuronal determination and promote glial differentiation instead. However, EGF receptor activation has different effects on the subventricular zone (SVZ) in mice and rats, possibly because of species differences in SVZ cell composition. Specifically in the rat, EGF stimulation of the SVZ induces the formation of hyperplastic polyps. The present study aims at molecular and morphological characterization of these subventricular polyps. Using immunohistochemistry, electron microscopy, and gene expression analysis, we demonstrate in hyperplastic EGF-induced polyps an upregulation in protein expression of Sox2, Olig2, GFAP, nestin, and vimentin. We found polyp-specific dysplastic changes in the form of coexpression of Sox2 and Olig2. This highly proliferative, Sox2/Olig2 coexpressing dysplastic cell type is >10-fold enriched in the hyperplastic polyps compared with control SVZ and most likely causes the polyp formation. Unique ultrastructural features of the polyps include a lack of ependymal cell lining as well as a large number of cells with large, light, ovoid nuclei and a cytoplasm with abundant ribosomes, whereas other polyp cells contain invaginated nuclei but fewer ribosomes. EGF also induced changes in the expression of Id genes Id1, Id2, and Id4 in the SVZ. Taken together, we here demonstrate dysplastic, structural, and phenotypical changes in the rat SVZ following EGF stimulation, which are specific to hyperplastic polyps.
Introduction
T
All newly generated SVZ cells are believed to originate from a quiescent glial fibrillary acidic protein (GFAP)-positive, radial glia-like stem cell, also termed B-cell [1,4]. Following asymmetric division, one of the daughter cells develops into a highly proliferative transit-amplifying progenitor, a C-cell, which expresses markers such as epidermal growth factor receptor (EGFR), Dlx2, and Olig2. Whether or not the C-cell is a universal feature in the mammalian SVZ is being debated. Although well defined in mice and humans, in terms of both protein expression and ultrastructural properties [5 –7], the existence of a C-cell in the rat, based on ultrastructural features, has been questioned [8]. The C-cells give rise to migrating neuroblasts also termed A-cells. Positive for doublecortin (DCX) and the polysialylated form of the neural cell adhesion molecule, these cells migrate from the SVZ via the rostral migratory stream to the olfactory bulb where they eventually become mature granule cells or periglomerular neurons [9].
EGF was first studied in the context of adult neural stem cells by Weiss and colleagues, when culturing stem cells from the adult murine striatum/ventricle, as so-called neurospheres required the presence of EGF to induce proliferation of multipotent cells [10]. The EGF responsive stem cells are located to the SVZ [11]; however, the primary endogenous ligand to the EGFR is believed to be transforming growth factor alpha (TGFα) [12], which has similar effects to EGF in vivo [13]. EGFR-mediated signaling in the central nervous system is involved in regulating a wide range of functions besides proliferation, including migration and cell fate determination [14 –17]. Growth factor infusion into the lateral ventricle has become a well-established experimental model for exploring therapeutic paradigms or as a model for disease [18]. In mice and rat, intracerebroventricular (ICV) infusion of EGF results in SVZ stem and progenitor cell proliferation [19] and also in a shift toward glial fate at the expense of neuronal differentiation [20]. The highly proliferative transit amplifying C-cells, the major EGF responsive cell population, change their phenotype toward a migratory glial-like cell upon extended EGF infusion [6]. In mice, these migratory cells have been described as oligodendrocyte progenitors, capable of homing to demyelinated lesion sites and of differentiation to oligodendroglial cells [21].
Hyperplastic polyp formation is persistently observed after ICV and intrastriatal infusion of EGF or TGFα in rats [13,20,22,23], but not in mice [6,19,24], except in the study by Ninomiya and colleagues [25]. The hyperplastic polyps were first described to be proliferatively active and EGF dependent [20] and later found positive for vimentin [22], nestin, and GFAP [13].
EGF stimulation is currently being explored for therapeutic purposes and as a model for pathological events, such as Parkinson's disease [23], demyelinating diseases [24], and ischemia [26]. However, fundamental knowledge regarding the effect of EGF on stem cells in the subventricular neurogenic niche is missing. The present study characterizes EGF-induced structural and dysplastic cellular changes, specifically prominent in hyperplastic polyps in the rat SVZ at the ultrastructural, protein, and mRNA levels.
Methods
Animals
All experiments were approved by the Gothenburg Committee of the Swedish Animal Welfare Agency (application nos. 214-07 and 145-10). About 8–9-week-old male Fischer-344 rats (166–235 g, 37 in total; Charles River, Germany) were housed in a barrier facility with ad libitum access to food and water with a 12-h light/dark cycle. Animals processed for immunohistochemistry were sacrificed by overdose of pentobarbital administered intraperitoneally, followed by transcardial perfusion using 0.9% saline solution followed by phosphate-buffered (0.1 mol/L, pH 7.4) 4% paraformaldehyde (Thermo Fisher Scientific, Waltham, MA). For mRNA expression analysis, animals were anesthetized using isofluorane (Merck, Whitehouse Station, NJ) and sacrificed by cervical decapitation.
Surgery
The animals were anesthetized with a cocktail of ketamine (Ketalar; Pfizer New York, NY) (33 mg/mL) and xylazine (Rompun, Beyer, Leverkusen, Germany) (6.67 mg/mL) administered intraperitoneally at a dose of 3 mL/kg. Anesthetized rats were mounted into stereotactic frames (David Kopf, Tujunga, CA and Stoelting Co, Wood Dale, IL) and surgical procedures were performed as described by Kuhn et al. [20]. The stereotactic coordinates used for cannula placement were the same as previously described [20] (anteroposterior [AP] +8.5 mm, lateral +1.2 mm from the center of the interaural line at flat skull position; cannula length, 5 mm below skull). The cannula was connected by a 3-cm vinyl tubing (Brain Infusion Kit 2; Alzet-Durect, Cupertino, CA) to an osmotic minipump (Model 2002; Alzet-Durect). Minipumps were filled with 30 μg/mL human recombinant EGF (Invitrogen, Carlsbad, CA) dissolved in artificial cerebrospinal fluid (aCSF) containing 100 μg/mL rat serum albumin (Sigma-Aldrich, St. Louis, MO). aCSF consisted of NaCl (148 mM), KCl (3 mM), CaCl2·2H2O (1.4 mM), MgCl2·6H2O (0.8 mM), Na2HPO4 (1.5 mM), and NaH2PO4·H2O (0.2 mM). Dental cement (Heraeus Kulzer, South Bend, IN) and 1/16-inch-diameter screws (Plastic One, Roanoke, VA) were used to fix the cannula to the skull.
Histology
Following perfusion, the brains were dissected and postfixed in phosphate-buffered (0.1 M, pH 7.4) 4% paraformaldehyde for 24 h and subsequently transferred to 0.1 M phosphate-buffered 30% sucrose at 4°C to be stored until further processed. The brains were coronally cut into 40-μm-thick serial sections using a sliding microtome (Leica Microsystems, Wetzlar, Germany). Sections were stored at 4°C in a cryoprotectant solution (glycerol, ethylene glycol, and 0.1 M phosphate buffer, pH 7.4, 3:3:4 by volume) as 1 in 12 series with 480 μm distance between sections.
Immunohistochemistry
Immunofluorescent labeling
The sections were washed 3 times in Tris-buffered saline (TBS) followed by blocking for 1 h in TBS, 3% normal donkey serum (Jackson ImmunoResearch, West Grove, PA), and 0.1% Triton-X at room temperature. For immunostaining against phospho-histone H3 (pHH3), an antigen retrieval step in 0.1 M NaCl (pH 6) at 80°C for 20 min was employed. Primary antibodies were diluted in blocking solution and sections were incubated for 24–72 h at 4°C. Sections were then washed 3 times in TBS. The following antibodies and concentrations were used: goat anti-DCX (1:250; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), rabbit anti-GFAP (1:1,000; DakoCytomation, Glostrup, Denmark), mouse anti-rat nestin (1:500; BD Pharmigen, San José, CA), rabbit anit-NG2 (1:500; Millipore, Billerica, MA), goat anti-human Olig2 (1:500; R&D Systems, Minneapolis, MN), rabbit anti-pHH3 (1:1,000; Millipore), goat anti-Sox2 (1:200; Santa Cruz Biotechnology, Inc.), rabbit anti-Sox2 (1:500; Millipore), and chicken anti-vimentin (1:2,000; Millipore). The sections were then incubated in the secondary antibodies donkey anti-goat 488, donkey anti-goat 546, donkey anti-mouse 488, donkey anti-mouse 555, donkey anti-mouse 647, donkey anti-rabbit 488 (1:2,000, Alexa Fluor; Molecular Probes, Eugene, OR), donkey anti-chicken FITC (1:2,000, Jackson ImmunoResearch), and donkey anti-rabbit CF 568 (1:1,000; Biotium, Hayward, CA) for 2 h at room temperature, followed by 5 rinses in TBS. The sections were mounted on slides and coverslipped with Pro-Long Gold (Molecular Probes/Invitrogen, Carlsbad, CA).
Microscopy and quantification
The SVZ was visualized by DAPI (4,6-diamidino-2-phenylindole, dihydrochloride, ProLong Gold with DAPI; Molecular Probes/Invitrogen) and the area was measured on at least 3 sections per animal between anterioposterior coordinates 10.3 and 7.2 mm from interaural line for volume extrapolation. The contralateral SVZ volume was calculated using Leica DM6000 B Stereomicroscope (Leica Microsystems) and StereoInvestigator 8 software (MBF Bioscience, Williston, VT).
Immunofluorescence for cell-specific protein expression was analyzed using a Leica SP2 scanning confocal microscope (Leica Microsystems). About 100–300 cells per animal were randomly selected in the contralateral SVZ of control and EGF-treated animals using ToPro3 (Molecular Probes/Invitrogen), along with 100 cells per contralateral hyperplastic polyp. The contralateral SVZ was analyzed to avoid possible artifacts due to tissue damages from the cannula. Analysis was performed on confocal stacks with 6–7 μm thickness acquired at 1.5 μm increments. Double labeling was assumed when cells exhibited direct colocalization or when nuclei and cytosol or processes from the same cell were individually labeled.
Proliferation
Analysis of the proliferating population in the SVZ was performed using the M-phase marker pHH3. The percentage of mitotic cells, as labeled by pHH3, that were also positive for either Sox2 or Olig2 was determined using confocal microscopy. For Sox2/pHH3 colocalization, 64.4±9.38 pHH3+ cells per EGF animal and 50.6±6.16 pHH3+ cells per aCSF animal (n=5 for both groups) were analyzed. For Olig2/pHH3 colocalization, 62±10.8 pHH3+ cells per EGF animal and 35±3.65 pHH3+ cells per aCSF animal were analyzed (EGF n=4, aCSF=5).
Polymerase chain reaction
The ipsi- and contralateral SVZ of aCSF- and EGF-infused animals was microdissected and flashfrozen in liquid nitrogen (n=5 for both groups). After homogenizing the tissue in trizol (Qiazol; Qiagen, Hilden, Germany), the mRNA was precipitated using chloroform (Merck) and then extracted using RNeasy mini or micro kits (Qiagen), depending on sample size. The concentration and quality of the mRNA was measured using a Nanodrop ND-1000 spectrophotometer (Thermo Fisher Scientific) followed by cDNA synthesis using random hexamer primers (High-Capacity cDNA Reverse Transcription kit; Applied Biosystems, Foster City, CA). The purity of the mRNA was estimated using the 260/280 nm ratio, with a mean value of all samples at 2.02±0.012. Two micrograms of total RNA was synthesized into cDNA and 8 ng cDNA was loaded to each 20 μL qPCR reaction. No template controls (reactions lacking cDNA) and −RT controls (mock cDNA synthesis performed without reverse transcriptase) were included to ensure that no contamination or genomic DNA was present. Quantitect primer assays (Qiagen) for β-actin (Actb QT00193473), inhibitor of DNA binding 1 (Id1 QT00374220), inhibitor of DNA binding 2 (Id2 QT00367640), inhibitor of DNA binding 4 (Id4 QT00383929), EGFR (Egfr QT00189707), and glyceraldehyde-3-phosphate dehydrogenase (Gapdh QT00199633) together with Maxima SYBR green master mix (Fermentas, Burlington, Ontario, Canada) were used for qPCR, according to the primer manufacturer's instructions. A Roche Lightcycler 480 (Roche, Basel, Switzerland) was used for PCR amplification. The efficiency of each primer was determined using a 1:4 dilution standard curve and taken into account when calculating the relative expression level. Calculations of C q values were made in LightCycler 480 software version 1.5 (Roche). The final fold change (aCSF vs. EGF) was calculated after correction for efficiency and normalization against 2 reference genes (Gapdh and Actb) according to Vandesompele et al. [27], using the C q values.
Electron microscopy
For transmission electron microscopy (TEM), tissue was processed as previously reported [28]. In short, animals were perfused as described earlier and the brains were postfixed overnight in a modified Karnowsky's solution (2% paraformaldehyde, 2.5% glutaraldehyde, and 0.05 Na-azide in 0.05 mol/L Na-cacodylate), followed by incubation in 1% OsO4 (osmium tetroxide) and 1% K4Fe(CN)6 (potassium ferrocyanide) in 0.1 mol/L Na-cacodylate for 2 h. Following treatment with uranylacetate, the tissue was dehydrated using a graded series of ethanol and then infiltrated using acetone and Agar 100 resin (Agar 100; Agar Scientific Ltd., Stanstead, United Kingdom). After evaluation of semithin sections (0.5 mm), 60–70-nm sections were cut using a Reichert-Jung Ultracut E (Leica Microsystems). Sections were counterstained using lead citrate and uranylacetate and collected on copper grids. The sections were imaged using a LEO912AB transmission electron microscope (Zeiss, Oberkochen, Germany) equipped with a Megaview III CCD camera (Olympus Soft Imaging Solutions, Hamburg, Germany).
For scanning electron microscopy (SEM), the ipsilateral SVZ was dissected after perfusion and postfixed in modified Karnowsky's solution. After rinsing the tissue with sodium cacodylate buffer, the tissue was made conductive by repeated treatment with OsO4 [29]. The tissue was then dehydrated in a graded series of ethanol and dried using hexamethyldizilasane. When mounted, the tissue was sputtered with palladium and examined using a Zeiss DSM 982 Gemini scanning electron microscope (Zeiss).
Statistics
When comparing 3 groups (aCSF SVZ, EGF SVZ, and hyperplastic polyps), 1-way analysis of variance was used in conjunction with Tukey's post hoc test using GraphPad Prism 4 (GraphPad Software, Inc., San Diego, CA). When comparing 2 groups for mRNA analysis and characterization of proliferating cells (aCSF and EGF), Student's t-test was employed. Error bars represent standard error of the mean (SEM) and differences of P<0.05 were considered significant (*). Cell numbers are presented as mean±SE and percentages as mean±SEM.
Results
To analyze the cellular composition of EGF-induced hyperplastic polyps, we infused EGF at 360 ng/day for 14 days into the lateral ventricle using osmotic minipumps. Control animals received vehicle (aCSF) infusion. Animals were sacrificed at day 14 for electron microscopy, immunocytochemical, and gene expression analyses.
No hyperplastic polyps were observed in vehicle-infused animal in either hemisphere (Fig. 1A). In all EGF-infused animals, polyp-like hyperplasias were observed in the lateral wall of the lateral ventricle of the ipsilateral hemisphere as well as in the majority (7 of 11 EGF-infused animals processed for immunohistochemistry) in the contralateral ventricle wall (Fig. 1B). In addition to polyp formation, we observed a general enlargement of the contralateral SVZ after EGF infusion, indicated by an increased SVZ volume (Fig. 1C). The nonpolyp-forming EGF-stimulated SVZ will be referred to as EGF SVZ.
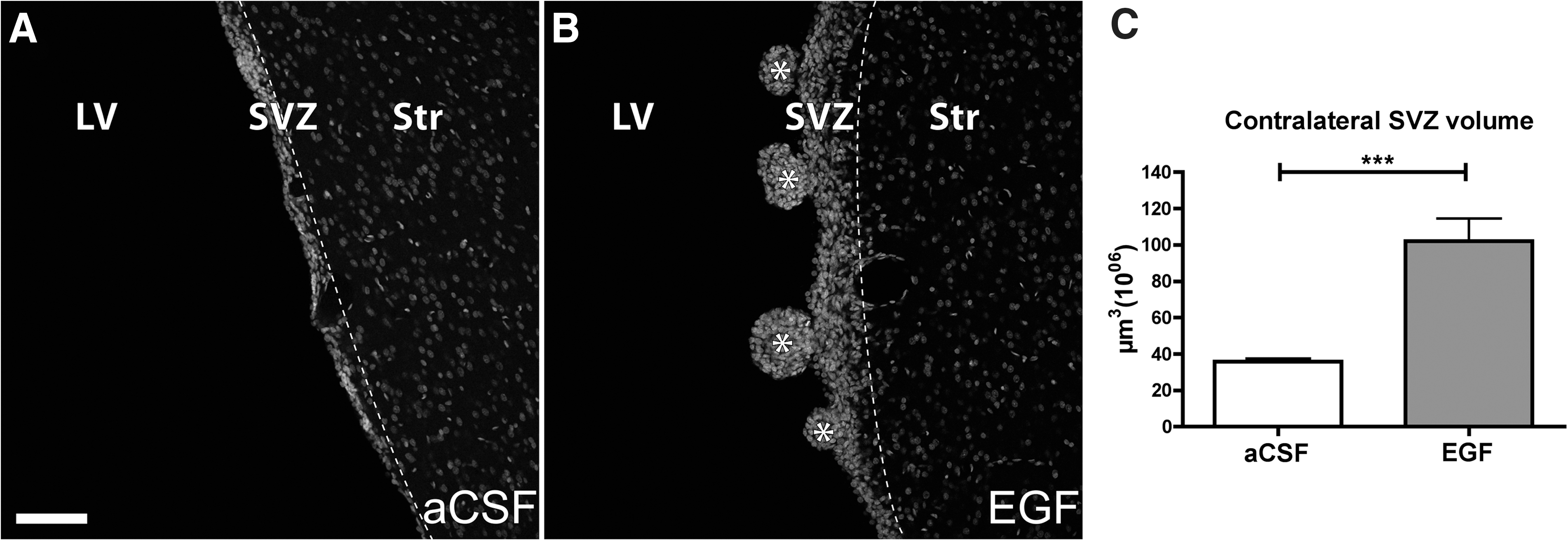
Contralateral SVZ.
Profound structural changes following EGF infusion; polyps devoid of ciliated ependymal cell lining
The surface of the EGF-treated ventricle wall was studied using SEM. In regions of the EGF-treated ventricle wall lacking hyperplastic polyps, we observed ciliated ependymal cells covering the lumen of the lateral ventricle wall (Fig. 2A, B). The hyperplastic polyps protruding from the ventricle wall ranged in size from 25 to 200 μm in diameter (Fig. 2C). On the luminal side of the polyps, a ciliated ependymal cell layer was absent. Instead, loosely associated nonciliated cells, rich in excess membrane, constituted the outermost cell layer of the polyps (Fig. 2D).
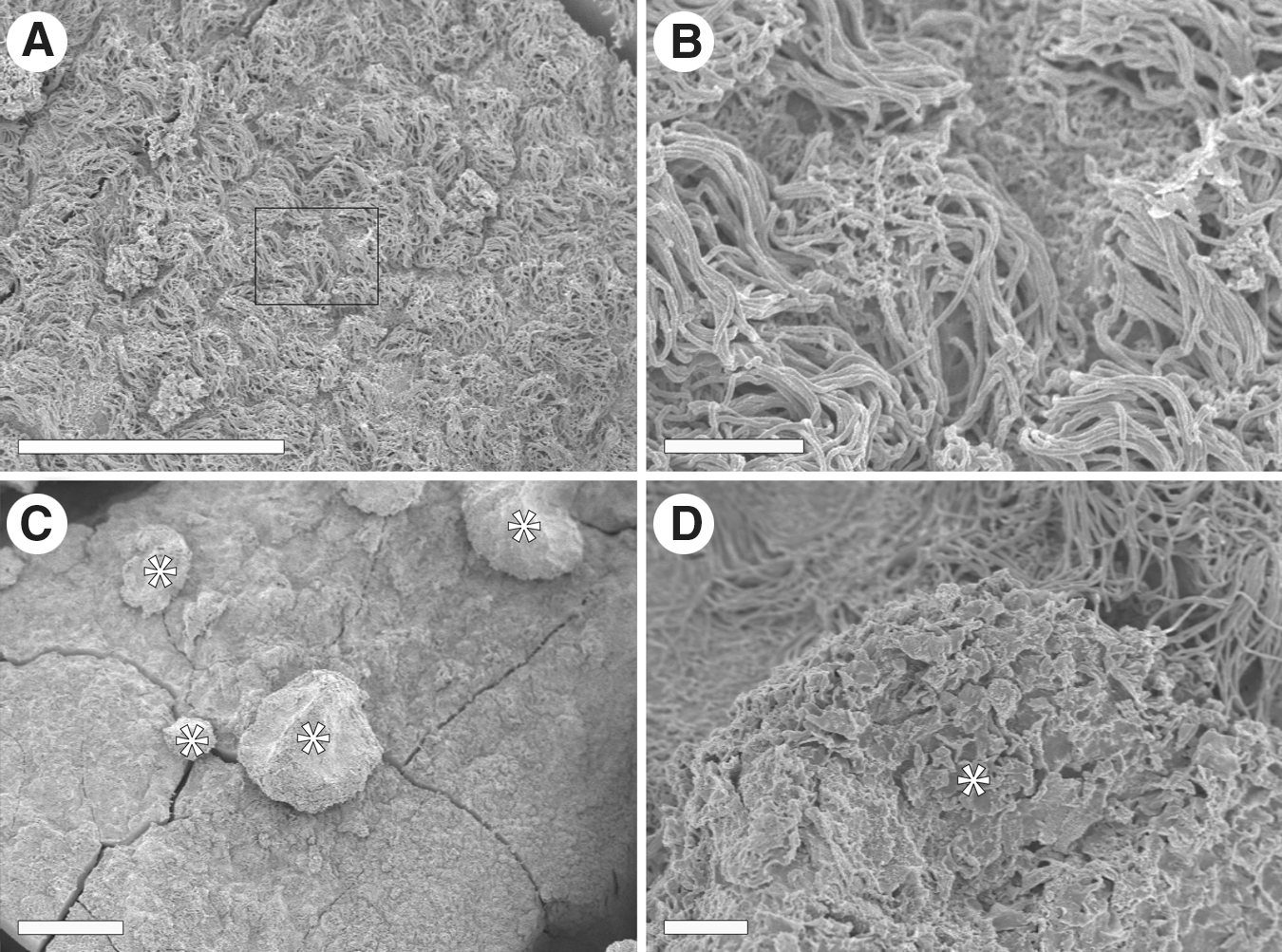
Scanning electron microscopy of the lateral ventricle wall after EGF infusion in whole mount.
Hyperplastic polyps are enriched for immature stem cells, but devoid of neuroblasts
Being separate structural entities protruding from the SVZ (Fig. 3A), hyperplastic polyps were characterized using markers for neural stem and progenitor cells, in relation to control SVZ and EGF SVZ. The neuroepithelial marker nestin was expressed in 44.7%±6.9% of the cells within the polyps, a 2-fold increase compared with the control SVZ (22.5%±2.6%) (Fig. 3B). The glial and stem cell marker Sox2 was expressed heterogeneously, but abundantly in the polyps. Overall, a majority of the polyp cells (65.8%±12%) expressed Sox2, compared with only 29.4%±1.9% in the control SVZ (Fig. 3C). Similar differences in the expression ratios were evident for astrocyte markers GFAP (Fig. 3D) and vimentin (Fig. 3E), for which twice as many cells in the polyps were found positive (39%±8.3% and 35.7%±7.5%, respectively) compared with control SVZ (22.3%±2.5% and 19.5%±1.8%). For all markers analyzed, the proportion in the EGF SVZ were in between EGF polyps and aCSF SVZ and not significantly different from either.

Comparison of cell type-specific markers between control SVZ (aCSF SVZ), EGF SVZ, and hyperplastic polyps (EGF Hyp).
Olig2 is normally expressed by both neural and oligodendrocyte progenitors and has been demonstrated to be upregulated after EGF infusion in mice [24]. We found a substantial increase in the proportion of Olig2-expressing cells in the polyps. The percentage of Olig2-expressing cells was 9-fold higher in hyperplastic polyps (21.4%±2.7%) compared with the control SVZ (2.4%±0.5%) (Fig. 3F). Chondroitin sulfate proteoglycan NG2, which is expressed in oligodendrocyte progenitors and upregulated in the periventricular area after EGF infusion in mice [24], was expressed at low levels in all groups. About 2.3%±0.5% of cells in polyps were NG2 positive, which did not significantly differ from EGF SVZ (2.0%±0.6%) or control SVZ (1.6%±0.3%) (Fig. 3G). In EGF-treated animals, the percentage of DCX-expressing, neuronally committed progenitor cells was downregulated 30-fold in polyps (0.7%±0.2%) and 8-fold in EGF-treated SVZ (2.6%±0.9%) compared with control SVZ (21.3%±2.0%) (Fig. 3H).
Abundance of stem cell-like cells and dysplastic changes in hyperplastic polyps
SVZ stem cell lineages are rarely defined by single marker expression, and thus, we decided to analyze coexpression of several markers (Fig. 4A–D). Nestin and GFAP were used to label early stem/progenitor cells. In the hyperplastic polyps, a 9-fold increase of colabeled cells (26.6%±10.4%) was evident compared with the control SVZ (3.0%±0.8%) (Fig. 4A), indicating an enrichment of stem cells within the hyperplastic polyps after EGF infusion. This notion is further strengthened by the increase in the percentage of GFAP/Sox2 double-labeled cells in the polyps, which is nearly 3-fold higher (36.7%±8.3%) compared with both control (13.7%±3.5%) and EGF-infused SVZ (12.4%±2.8%) (Fig. 4B).
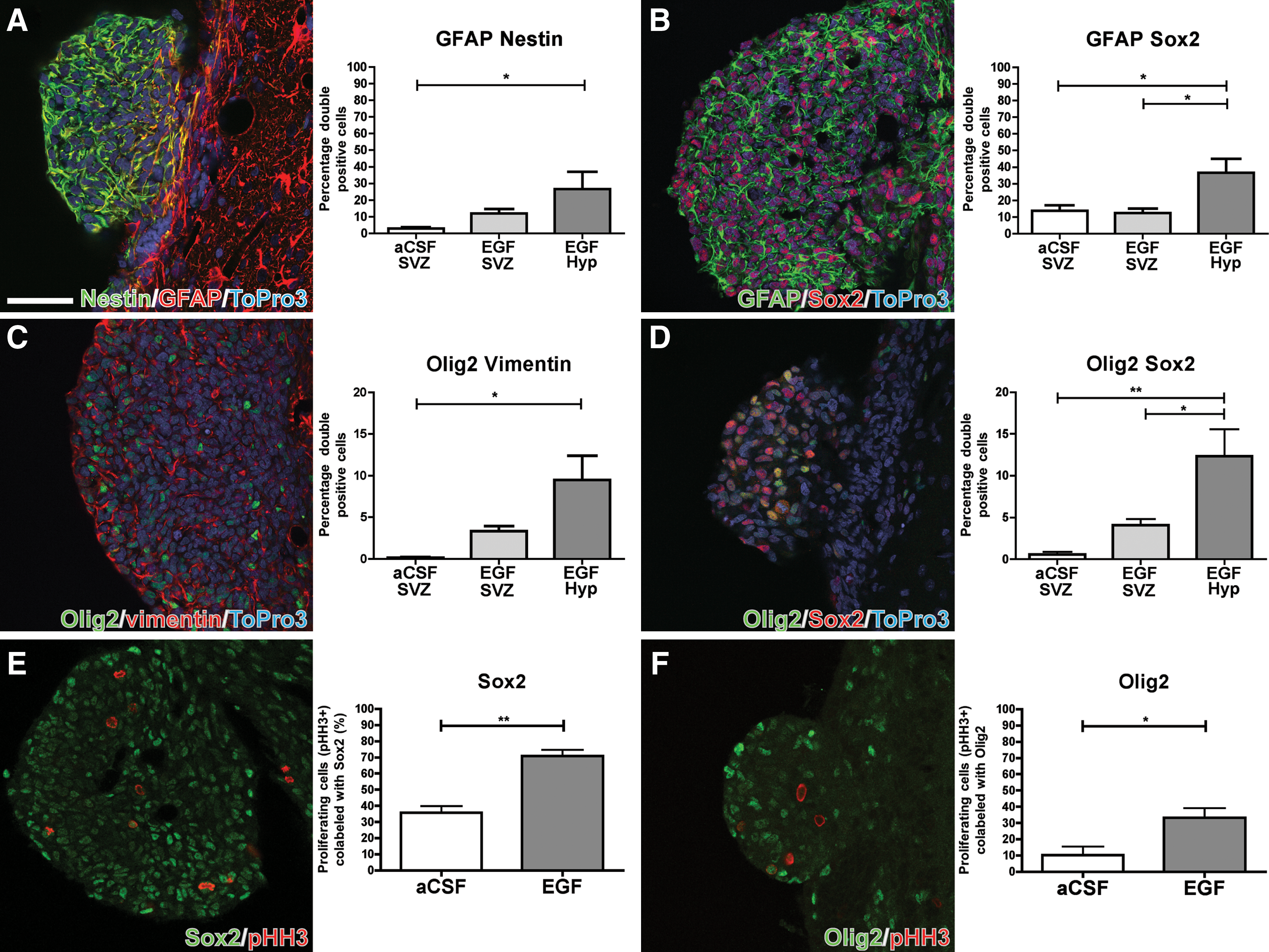
Confocal microscopy images and quantifications of protein coexpression of
Olig2 and vimentin, markers for oligodendrocyte/progenitor and glial cells, respectively, normally label cells of different lineages; however, we found a significantly higher percentage of Olig2/vimentin-colabeled cells in the polyps (9.5%±2.9%) compared with the control SVZ (0.17%±0.09%) (Fig. 4C). The fraction of Olig2/vimentin double-positive cells in the nonhyperplastic EGF-infused SVZ (3.3%±0.6%) was not significantly different from control SVZ. We further characterized Olig2-positive progenitors for coexpression of Sox2 (Fig. 4D). Similar to Olig2/vimentin colabeling, roughly 1/10th (12.3%±3.2%) of polyp cells coexpressed Sox2/Olig2, which was a significantly higher amount than in both EGF-treated SVZ (4.1%±0.7%) and control SVZ (0.57%±0.3%), in which we rarely found colabeling. Because of the heterogeneity of Olig2-expressing cells in the SVZ, Olig2/NG2 colabeling was quantified to target oligodendrocyte committed progenitors. The percentage of Olig2/NG2-colabeled cells was nearly identical in all groups, being 0.7% in hyperplastic polyps and EGF SVZ and 0.6% in control SVZ.
Dysplastic cells are part of the proliferating population after EGF treatment
EGF infusion has been shown to increase proliferation in the SVZ 10-fold [20]. To determine the phenotype of the proliferating cells in the EGF-stimulated SVZ, we used pHH3 to label cells in the M-phase of the cell cycle, in combination with Sox2 (Fig. 4E) and Olig2 (Fig. 4F), which showed the greatest increase after EGF stimulation. Sox2 and Olig2 were found in a large portion of the pHH3-positive population in the EGF-treated lateral ventricle wall, with 71%±3.7% and 33.2%±5.9%, respectively, compared with 35.8%±4.2% and 10.4%±5.1% in control SVZ (Fig. 4E, F).
Unique ultrastructural features of polyp cells further indicate dysplastic changes
We further characterized the SVZ and the hyperplastic polyps of EGF-infused animals using TEM. The ependymal cells of the EGF-treated SVZ were phenotypically normal, with apically oriented cytoplasm, cilia (9+2 microtubule pairs; not shown), and desmosomes indicating tight junctions (Fig. 5A). Cells residing in the control SVZ showed high levels of heterochromatin (Fig. 6A). In contrast, in EGF-infused animals, subependymal cells contained mostly euchromatin (Fig. 6B), the overall structure appeared disorganized, and cells found within the polyps displayed ultrastructural characteristics of different stages of immaturity. The periphery of the hyperplastic polyps was completely devoid of ependymal-like cells, confirming the SEM observations. Instead, cells lacking junctional complexes were most abundant. These cells had a light, elliptical nucleus with dispersed chromatin and a cytoplasm with abundant ribosomes, suggesting higher transcription and protein synthesis levels (Fig. 5B, C). In contrast, near the core of the polyps, cells with invaginated nuclei-prominent nucleolus, clumped chromatin along the inside of the nuclear membrane, and few ribosomes were found (Fig. 5D). In the control SVZ, no cells were found matching these characteristics.

Transmission electron microscopy (TEM) of ependyma and polyps.

TEM of SVZ.
EGF treatment induces heterogeneous changes in Id gene expression
Changes in gene expression in the SVZ after EGF infusion were studied using quantitative real-time RT-PCR. To estimate the magnitude of the effect resulting from EGF infusion, Egfr expression was measured. Compared with control SVZ, Egfr expression was 6-fold upregulated in EGF-treated animals (Fig. 7A). We also compared expression levels of inhibitor of DNA binding (Id) basic helix-loop-helix (bHLH) transcription factors, Id1, Id2, and Id4, which have been implicated in stem cell maintenance and oligodendrocyte differentiation [30 –33]. Id1, which is normally expressed in quiescent GFAP-positive neural stem cells and Olig2-positive progenitors, was upregulated 2-fold in the EGF-treated SVZ (Fig. 7B), whereas both Id2 and Id4 (Fig. 7C, D), mediators of the inhibitory effects of BMP4 signaling on oligodendrogenesis [31], were downregulated to 50% compared with aCSF controls.

mRNA quantification. EGF-induced changes in relative mRNA expression in SVZ of
Discussion
In summary, our results demonstrate substantial changes in ultrastructure and protein and mRNA expression, following EGF infusion, specific to EGF-induced hyperplastic polyps in the rat SVZ. A general increase in cells expressing stem cell and glial markers (summarized in Fig. 8), and acquired dysplasia in the form of Sox2/Olig2 coexpression, was observed in the polyps. The ultrastructural properties of polyp-forming cells indicate an immature phenotype with features not observed in either the control or EGF-infused SVZ. In addition, we show heterogeneous changes in the expression of several Id genes in the lateral ventricle wall after EGF infusion.

Illustration of changes in marker expression of Olig2, Olig2/Sox2, Sox2, Sox2/GFAP, GFAP, and DCX in SVZ cells following EGF infusion. Color images available online at
We found hyperplastic polyps in all animals that received EGF infusion; however, the majority of the studies using ICV administration of EGF or TGFα in mice do not report on polyps protruding into the ventricle. This is possibly due to the shorter infusion period of 6 or 7 days [6,19,24], because we and others observed polyps after 12 days or more of EGF infusion [20,25]. However, we observe hyperplastic polyps after 7 days of EGF infusion in rats (unpublished data). As for species differences, EGF-induced hyperplastic polyps appear to be much smaller and less frequent in mice, if detected at all, indicating a difference between mouse and rat in the response of SVZ stem and progenitor cells to EGF stimulation.
As previously reported, long-term EGF stimulation induces increased expression of markers for glial cells, such as tenascin-C and S100β, along with a reduction of C-cells [6,20,24]. We here show that an increased number of stem and progenitor cells in the EGF-stimulated rat SVZ express Sox2 and Olig2, but also GFAP, nestin, and vimentin. Interestingly, coexpression of markers associated with stem cells, such as GFAP/Sox2 and GFAP/nestin, rather than markers for progenitors or differentiated cells, were increased in the polyps. In the rat SVZ, nestin is predominantly found in the EGFR-expressing progenitor cells [8], whereas in mice nestin is more ubiquitously expressed across SVZ cell types [5]. Further, the hyperplastic polyps featured unique dysplastic changes in the form of acquired Olig2/Sox2 coexpression. An increase in coexpression of Olig2 and NG2 has been reported after EGF infusion in mice [24]. However, we found no indication of an oligodendroglial commitment of dysplastic cells in polyps of the rat SVZ.
The hyperplastic polyps protruding into the ventricles displayed an irregular and flakey surface at the EM level and were lacking an ependymal cell layer. The lack of a liquor-brain-barrier may provide a unique milieu for the cells residing in these structures, including more direct access and higher concentration of EGF and other molecules present in the CSF, possibly explaining some of the differences in ultrastructure and protein expression exhibited in the polyps compared with control SVZ and EGF SVZ.
The ultrastructural features of the cells found in the polyps are in line with the immunohistochemical data, indicating an immature phenotype and dysplasia. The dispersed chromatin and abundance of ribosomes in the cells located peripherally in the polyps is suggestive of active transcription and protein synthesis. A portion of these cells displayed a prominent nucleolus, although the majority of cells with a nucleolus had an invaginated nuclear membrane and few ribosomes and were located in the polyp core. This suggests a continuum of EGF-stimulated cells, at different stages of development, within the polyps. However, a clear compartmentalization was not observed in all polyps, but appeared to be more frequent in the polyps with a smaller diameter (under 100 μm). Although we cannot directly link these ultrastructural features within the polyps to the dysplastic Sox2/Olig2-coexpressing cell, no cells displaying the same EM features were found outside the polyps.
The relative abundance of euchromatin found in the EGF-stimulated SVZ indicates that cells are more transcriptionally active compared with the control SVZ. The protein expression data point toward a higher ratio of cells with a stem cell phenotype in the polyps and EGF SVZ compared with the control SVZ and the extent of euchromatin in cells residing in the EGF SVZ and hyperplastic polyps also suggests a lower level of maturity.
Id1 protein is expressed by both quiescent neural stem cells (B1-cells) and Mash1- and Olig2-positive progenitors [30]. We found the mRNA expression of bHLH gene Id1 upregulated 2-fold in the EGF-stimulated SVZ compared with controls. This upregulation in gene expression matches the phenotype of the EGF-stimulated cells that we describe and includes an increase in GFAP, Sox2, and Olig2. Increases in Id1 expression has been previously linked to both inhibition of differentiation and to induction of proliferation (for review, see Ref. [34]). More specifically, loss of Id1 and Id3 inhibits neuronal differentiation during development [33]. Id2 and Id4, members of the same family of proteins, are known to inhibit oligodendrocyte differentiation [32,35,36] and have been shown to directly bind to Olig2, inhibiting Olig2-induced transcription [31]. Reduced expression of these genes could be one of the mechanisms promoting increased Olig2 protein expression levels, which we found to be induced by EGF stimulation. Moreover, Olig2 has been demonstrated to possess cell cycle regulating properties by transcriptional repression of cyclin-dependent kinase inhibitor p21 in both neural stem cells and gliomas [37], providing one possible explanation for the proliferative and Olig2-inducing effects of EGFR activation.
A positive feedback loop between Sox2 and EGFR has been suggested for neural progenitor cells, where EGFR signaling promotes Sox2 protein expression in vitro [38]. In turn, Sox2 binds to the EGFR promoter and further stimulates EGFR expression. This could explain the substantial increase in Sox2 expression seen after EGF treatment in vivo. As Sox2 expression maintains stem cell multipotency and inhibits neural differentiation [39], it is possible that EGF-stimulated dividing progenitors remain in a stem cell-like proliferative state as a result of the high Sox2 levels. The ultrastructural properties of the polyp-forming cells further supports this notion of immature cells enriched in the hyperplastic polyps.
EGFR function is also closely related to tumor development and malignancy, and its expression level is an important diagnostic criterion [40]. Expression of the wild-type EGFR protein is increased in 50% of all glioblastoma multiforme tumors [41]. EGF is also involved in regulating self-renewal capacity and sphere formation of brain cancer stem cells in vitro [42]. A putative link between gliomas and neural stem cells has been described [43 –46]. Both tumor cells and intermediate progenitor cells are highly proliferative, migratory, and capable of differentiating into multiple lineages. Further, Olig2 and Sox2 expressions are increased after EGF stimulation. Ligon and colleagues [37] showed that Olig2 is required for glioma formation and Sox2 was found to be overexpressed in malignant gliomas [47]. Despite the similarities to the dysplastic cells in the polyps, the transient and nonmalignant nature of the polyps, which disappear after discontinuation of EGF stimulation [20], shows that EGF stimulation alone is not sufficient to create permanent neoplasms. The differential changes in Id gene expression in the SVZ after EGF infusion could represent an intermediate dysplastic state, compared with a general upregulation of Id genes often seen in cancer [48].
Previously published work has focused on progenitors migrating into the striatum after EGF infusion [6,23,24] and no detailed characterization of the changes in the rat SVZ after EGF stimulation has been performed. Here, we show that the hyperplastic polyps are separate entities, which provide a unique niche compared with the EGF-stimulated and control SVZ. Our study shows that the SVZ cells are highly plastic and can be influenced by external cues to shift from a neurogenic to a gliogenic nature. Whether EGF changes the fate of a multipotent progenitor cell or whether separate SVZ progenitor types with glial and neuronal commitment are differentially affected remains open. The inherent plasticity in neural stem and progenitor cells indicates a potential use of endogenous adult neural stem cells in therapy. However, mitogenic stimulation can induce species-specific dysplastic changes, which in many aspects resemble tumor cells. More work is required before the full repertoire of action of EGF on neural stem and progenitor cells is understood and its putative therapeutic potential properly investigated.
Footnotes
Acknowledgments
The authors thank Birgit Linder for excellent technical assistance during animal experiments and Yvonne Josefsson for EM sample preparation. The authors also thank Charlotta Lindwall and Nina Hellström for critically reviewing the manuscript. This work was supported by grants from the Swedish Medical Research Council, the Swedish Childhood Cancer Foundation, Hjärnfonden, Vinnova, Västra Götaland regional funds for biomedical research (LUA-ALF), and the Sten A Olsson Foundation for Research and Culture. This work has been presented as a poster abstract at the Winter Conference on Brain Research, Copper Mountain, CO, January 2008, and at the Society for Neuroscience Yearly Conference, San Diego, CA, November 2010.
Author Disclosure Statement
No competing financial interests exist.