
Abstract
Increasing evidence shows that neural stem/progenitor cells (NSPCs) can be activated in the nonconventional neurogenic zones such as the cortex following ischemic stroke. However, the precise origin, identity, and subtypes of the ischemia-induced NSPCs (iNSPCs), which can contribute to cortical neurogenesis, is currently still unclear. In our present study, using an adult mouse cortical infarction model, we found that the leptomeninges (pia mater), which is widely distributed within and closely associated with blood vessels as microvascular pericytes/perivascular cells throughout central nervous system (CNS), have NSPC activity in response to ischemia and can generate neurons. These observations indicate that microvascular pericytes residing near blood vessels that are distributed from the leptomeninges to the cortex are potential sources of iNSPCs for neurogenesis following cortical infarction. In addition, our results propose a novel concept that the leptomeninges, which cover the entire brain, have an important role in CNS restoration following brain injury such as stroke.
Introduction
S
Increasing evidence indicates that NSPCs [10,11] and neuronal progenitors [12] are induced at the subpial/cortical layer 1 regions in response to cortical ischemia/injury, suggesting that NSPCs can be activated in the cortical surface adjacent to an injured region. Leptomeninges (including pia mater and arachnoid mater), which cover the entire brain cortex, regulate NSPC development [13] and cortical neurogenesis [14]. In addition, a recent study has shown that stem/progenitor cells with neuronal differentiation potential reside in the leptomeninges during development [15]. However, no study has examined pia mater as a source of iNSPCs following cortical infarction.
In the present study we investigated whether the leptomeninges (pia mater), which is widely distributed within and closely associated with blood vessels throughout CNS including the cortex, have NSPC activity in response to ischemia and can generate neurons. Further, we investigated whether microvascular pericytes that are distributed from the pia mater to the cortex are the origin of iNSPCs following cortical infarction.
Materials and Methods
Animal studies
All procedures were approved by the Animal Care Committee of Hyogo College of Medicine and were in accordance with the criteria outlined in the “Guide for the Care and Use of Laboratory Animals” prepared by the National Academy of Science of the USA. Quantitative analyses were performed by investigators who were blinded to the experimental protocol and identity of the samples under study.
Induction of focal cerebral ischemia
Cerebral ischemia was induced in 6-week-old male CB-17/Icr-+/+Jcl mice (Clea Japan Inc., Tokyo, Japan). Permanent focal cerebral ischemia was produced by ligation and disconnection of the distal portion of the left middle cerebral artery (MCA) [5 –8,16]. In brief, the left MCA was isolated, electrocauterized, and disconnected just distal to its crossing of the olfactory tract (distal M1 portion) under halothane inhalation. The infarct area in mice of this background has been shown to be highly reproducible and limited to the ipsilateral cerebral cortex [5 –8,16].
Histological analysis
Immunohistochemistry was performed as described previously [5,6,8]. In brief, mice were deeply anesthetized with sodium pentobarbital (50 mg/kg) and perfused transcardially with 4% paraformaldehyde. Next, coronal brain sections (20 μm) were prepared and they were stained with antibodies to nestin (Chemicon, Temecula, CA), Ki67 (BD Pharmingen, San Diego, CA), 5-bromo-2′-deoxyuridine (BrdU; Abcam, Cambridge, United Kingdom), CD31 (Santa Cruz Biotechnology, Santa Cruz, CA), NG2 (Chemicon), platelet-derived growth factor receptor alpha (PDGFRα; Santa Cruz Biotechnology), PDGFRβ (Santa Cruz Biotechnology), glial fibrillary acidic protein (GFAP; DAKO, Glostrup, Denmark), A2B5 (Chemicon), Olig2 (Chemicon), Sox2 (Chemicon), Sox10 (R&D Systems, Minneapolis, MN), and glutamic acid decarboxylase 67 (GAD67; Sigma, St. Louis, MO). To quantify cell proliferation in vivo, 50 mg/kg of BrdU (Sigma) was administered subcutaneously daily for 3 days before sacrifice. Primary antibodies were observed using Alexa Fluor 488- or 555-conjugated secondary antibodies. Nuclei were stained with 4′,6-diamino-2-phenylindole (DAPI; Kirkegaard & Perry Laboratories, Gaithersburg, MD). Images of sections were captured using a confocal laser microscope (LSM510; Carl Zeiss, Jena, Germany). The number of nestin-positive cells was counted at the poststroke pia mater and cortex, the nonischemic ipsilateral area, and regions within or around the subventricular zone (SVZ; at a distance of 1–100 and 101–200 m from the ventricular wall, respectively). Each area (10 sections/mouse, n=5) was measured using ImageJ software from NIH Image as described previously [5 –7]. Results were expressed as the number of cells/mm2. In another set of experiments, mice were anesthetized and placed in a stereotactic apparatus 2 days before induction of ischemia. A lentiviral vector encoding green fluorescent protein (GFP; 5.2×105 transducing unit (TU)/μL) [17] was injected into the pia mater as described previously [5].
Cell culture
To examine whether nestin-positive cells developed in the poststroke pia mater were NSPCs, we cultured pial cells under the conditions that promoted the formation of neurospheres as described previously [5,6,8]. In brief, mice were deeply anesthetized with sodium pentobarbital on poststroke day 3, and the pia mater was carefully removed under a microscope (Carton, Pathumthani, Thailand). Absence of cortical contamination was confirmed by histological analysis. The removed tissues were then mechanically dissociated by passage through 18-, 23-, and 27-gauge needles to prepare a single-cell suspension. The resulting cell suspensions were incubated in Dulbecco's modified Eagle's medium (DMEM) containing 20 ng/mL of basic fibroblast growth factor (bFGF; Peprotech, Rocky Hill, NJ), 20 ng/mL of epidermal growth factor (EGF; Peprotech), and 1% N2 supplement (Invitrogen, Carlsbad, CA). Total number of neurosphere-like cell clusters exceeding 80 m in diameter was counted every day for 30 days using a phase-contrast microscope (Olympus, Tokyo, Japan). On day 7 of incubation, cell clusters were fixed, cut on a cryostat (8-μm thick sections), and stained with antibody to nestin, NG2 and GFAP as described previously [5,6,8]. In another set of experiments, whole brains were removed on poststroke day 3, and coronal brain slices (6 mm thickness) were incubated in DMEM containing bFGF and EGF. On day 7 of incubation, brain sections were fixed, cut on a cryostat into 20-μm-thick sections, and subjected to immunohistochemistry.
Differentiation of neurospheres
To induce cell differentiation, neurosphere-like cell clusters were incubated on poly-L-lysine-coated glass coverslips for 6 days in neurobasal medium (Invitrogen) containing the B-27 supplement (Invitrogen) and all-trans retinoic acid (0.2 μM; Sigma) [6]. Differentiated cells were stained with antibodies to microtubule-associated protein-2 (MAP2; Chemicon), neurofilament (Chemicon), Tuj-1 (Chemicon), GFAP (Chemicon), O4 (Chemicon), and myelin-associated glycoprotein (MAG; gift from Prof. Shinobu Inagaki, University of Osaka, Osaka, Japan).
Electron microscopy
Brains at poststroke day 3 were sequentially fixed in 1% glutaraldehyde, 1% OsO4, ethanol, and propylene oxide, and embedded in Epon as described previously [18]. Sections were stained with uranyl acetate/lead citrate and examined under a HITACHI 7100 electron microscope (Tokyo, Japan) operated at 75 kV.
Reverse transcription-polymerase chain reaction
Total RNA was extracted from embryonic stem (ES) cells as well as pia mater- and SVZ-derived spheres. cDNA was amplified under the following conditions: 15 s at 94°C, 30 s at 56°C, and 1 min at 68°C (40 cycles). Primer sequences were as follows: GFAP forward, TCGGCCAGTTACCAGGAGG and GFAP reverse, ATGGTGATGCGGTTTTCTTCG (amplicon size, 176 bp); aquaporin-4 (AQP4) forward, AACTGGGCAAACCACTGGATA and AQP4 reverse, TCTCCCGAAGAGTCTTTCCCC (amplicon size, 278 bp); glyceraldehyde-3-phosphate dehydrogenase (GAPDH) forward, TGGTATCGTGGAAGGACTCATGAC and GAPDH reverse, ATGCCAGTGAGCTTCCCGTTCAGC (amplicon size, 180 bp); Snail forward, CTTGTGTCTGCACGACCTGT and reverse, CTTCACATCCGAGTGGGTTT (amplicon size, 161 bp); Slug forward, AACATTTCAACGCCTCCAAG and Slug reverse, GCCGACGATGTCCATACAGT (amplicon size, 162 bp); Twist forward, ACGCAGTCGCTGAACGAGGC and Twist reverse, GTACAGGAAGTCGATGTACC (amplicon size, 119 bp); Sox9 forward, AAGAGGCCACGGAACAGACTCA and Sox9 reverse, GACCCTGAGATTGCCCAGAGTG (amplicon size, 132 bp); NG2 forward, ATGCTTCTCAGCCCGGGACA and NG2 reverse, GGTTGCGGCCATTGAGAATG (amplicon size, 424 bp); PDGFRβ forward, ACAATTCCGTGCCGAGTGACAG and PDGFRβ reverse, AAAAGTACCAGTGAAACCTCGCTG (amplicon size, 114 bp); Olig2 forward, ACGCCAGCCTGGTGTCTAGT and Olig2 reverse, TCGGGCTCAGTCATCTGCTT (amplicon size, 304 bp); nestin forward, CACTAGAAAGCAGGAACCAG and nestin reverse, AGATGGTTCACAATCCTCTG (amplicon size, 307 bp); Sox2 forward, TTGGGAGGGGTGCAAAAAGA and Sox2 reverse, CCTGCGAAGCGCCTAACGTA (amplicon size, 312 bp); Oct4 forward, AGCTGCTGAAGCAGAAGAGG and Oct4 reverse, GGTTCTCATTGTTGTCGGCT (amplicon size, 138 bp); Klf4 forward, GTGCCCCGACTAACCGTTG and Klf4 reverse, GTCGTTGAACTCCTCGGTCT (amplicon size, 185 bp); c-myc forward, ATGCCCCTCAACGTGAACTTC and c-myc reverse, CGCAACATAGGATGGAGAGCA (amplicon size, 228 bp); and β-actin forward, GCTCGTCGTCGACAAGGGCTC and β-actin reverse, CAAACATGATCTGGGTCATCTTCTC (amplicon size, 353 bp).
Western blot analysis
Samples (10 μg) were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Separated proteins were transferred electrophoretically onto nitrocellulose membranes. The membranes were incubated with antibodies to Sox2 (Chemicon), Oct4 (Chemicon), Klf4 (Santa Cruz Biotechnology), or c-myc-antibody (Santa Cruz Biotechnology) and then with peroxidase-labeled secondary antibodies. Antibody labeling of protein band was detected with enhanced chemiluminescence reagents (Chemi-Lumi One, Nacalai Tesque, Kyoto, Japan), according to the manufacturer's instructions.
Statistical analysis
Results are reported as the mean±standard deviation. Statistical comparisons among groups were determined by 1-way analysis of variance. Individual comparisons were performed using Student's t-test when appropriate. P<0.05 was considered to be statistically significant.
Results
Nestin-positive cells develop in the poststroke pia mater after ischemia
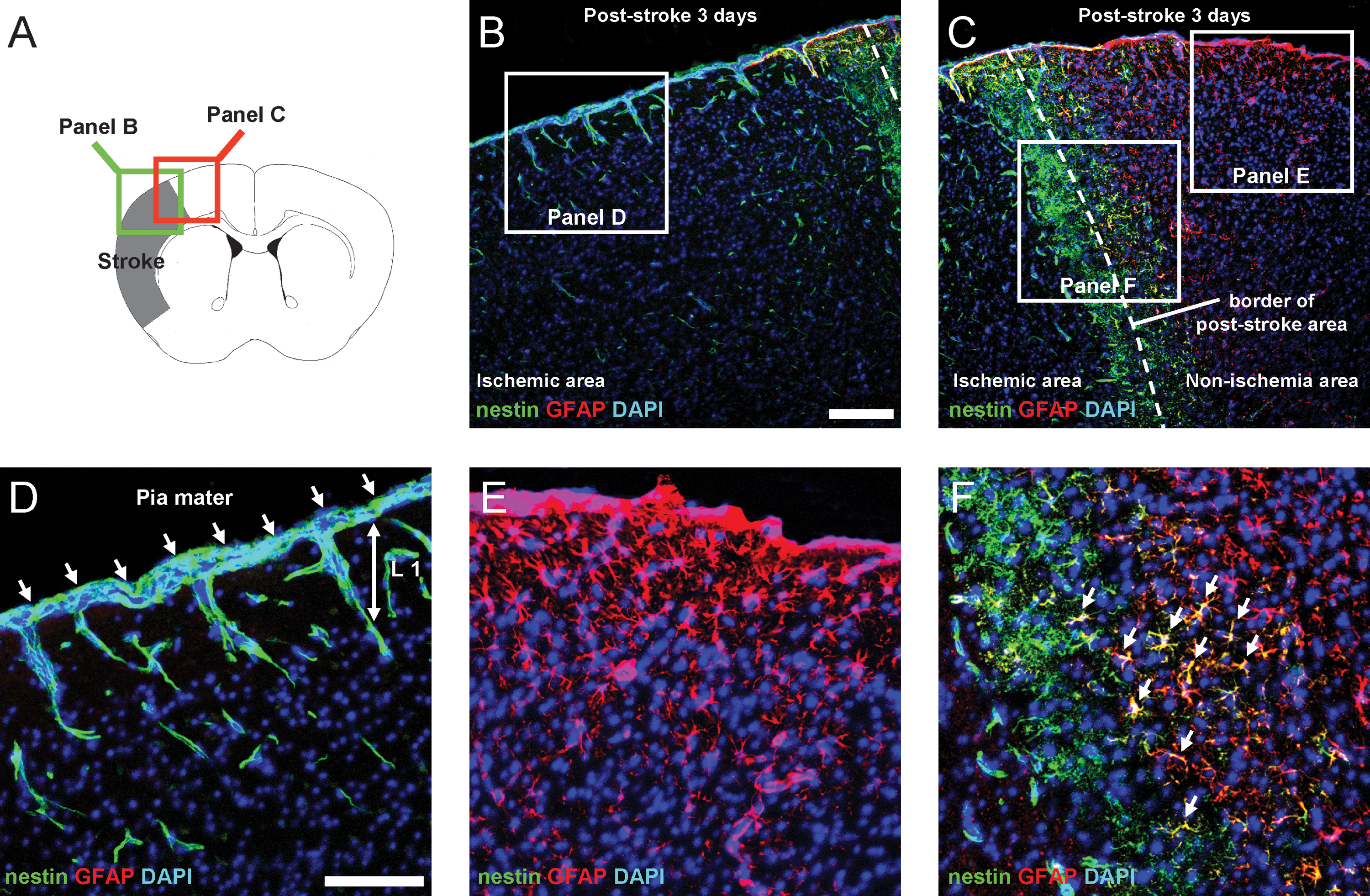
We reported previously that nestin-positive iNSPCs develop in the poststroke cortex in the brains of adult mice [5 –8]. Thus, we examined the location of nestin-positive cells on poststroke day 3 by immunohistochemistry to determine their precise origin. Consistent with our previous study [5], nestin-positive cells were observed in the poststroke cortex (Fig. 1A, B), whereas they were rarely detectable in the nonischemic cortex (Fig. 1A, C). Nestin-positive cells were expressed consistently in the conventional neurogenic zones such as SVZ (Fig. 1A, D). In addition to these areas, we found that nestin-positive cells developed in poststroke, but not in nonischemic pia mater (Fig. 1A–C). To evaluate cell proliferation in the poststroke pia mater, Ki67 was used as a marker of dividing cells [19]. On day 3 after stroke, extensive expression of Ki67 was observed in the poststroke pia mater/cortex, whereas few Ki67-positive cells were observed in the nonischemic ipsilateral pia mater/cortex (Fig. 2A–C). Further, cells that emerged in the poststroke pia mater/cortex could be birthdated with daily BrdU injections for 3 days, indicating that the nestin/BrdU double-positive cells in these regions were proliferative. In contrast, proliferative cells were rarely observed in the nonischemic ipsilateral pia mater/cortex (Fig. 2A, D–G).
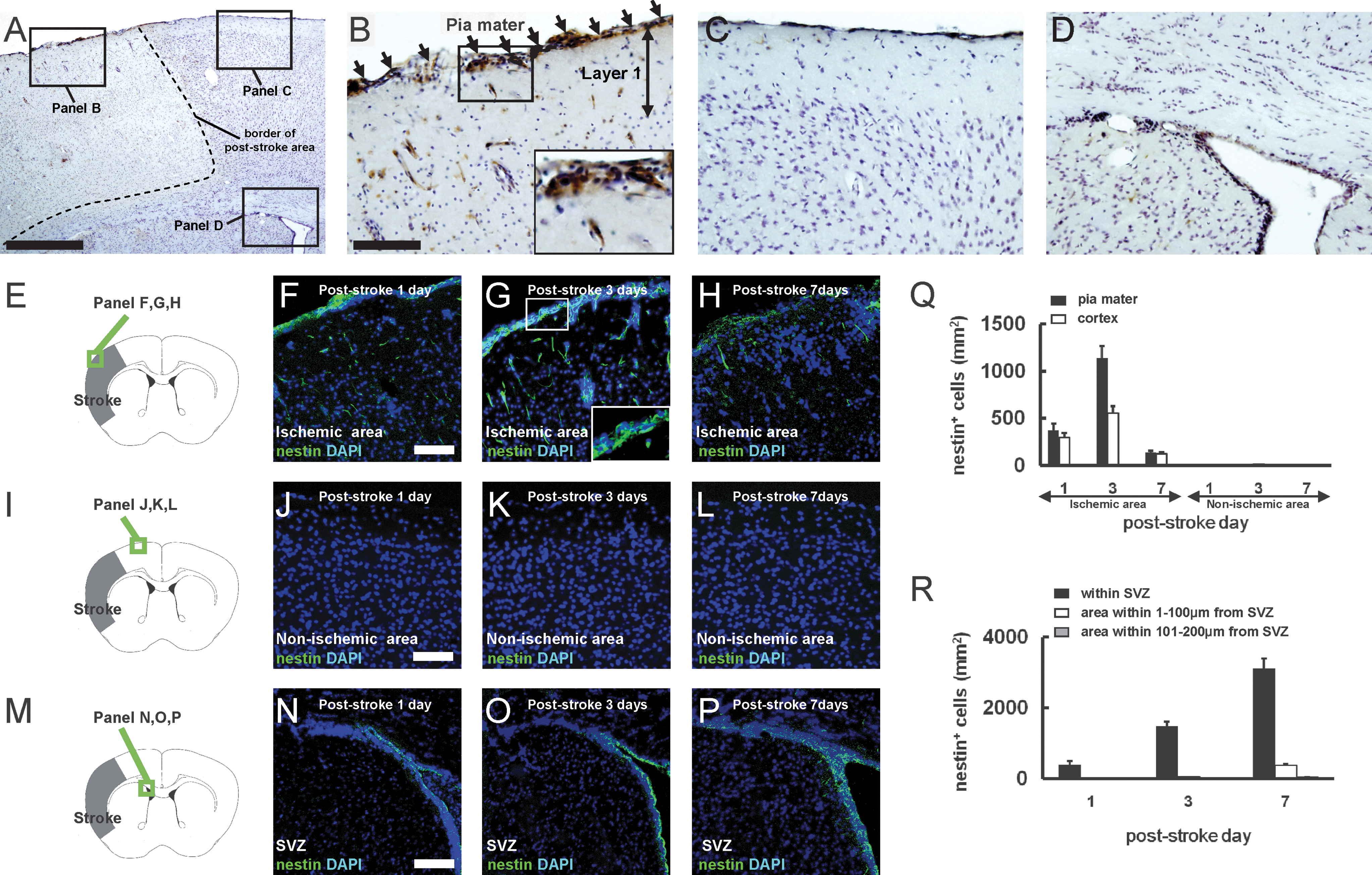
Nestin-positive cells develop in the poststroke pia mater. Immunohistochemistry for nestin using the diaminobenzidine (DAB) reaction 3 days after stroke
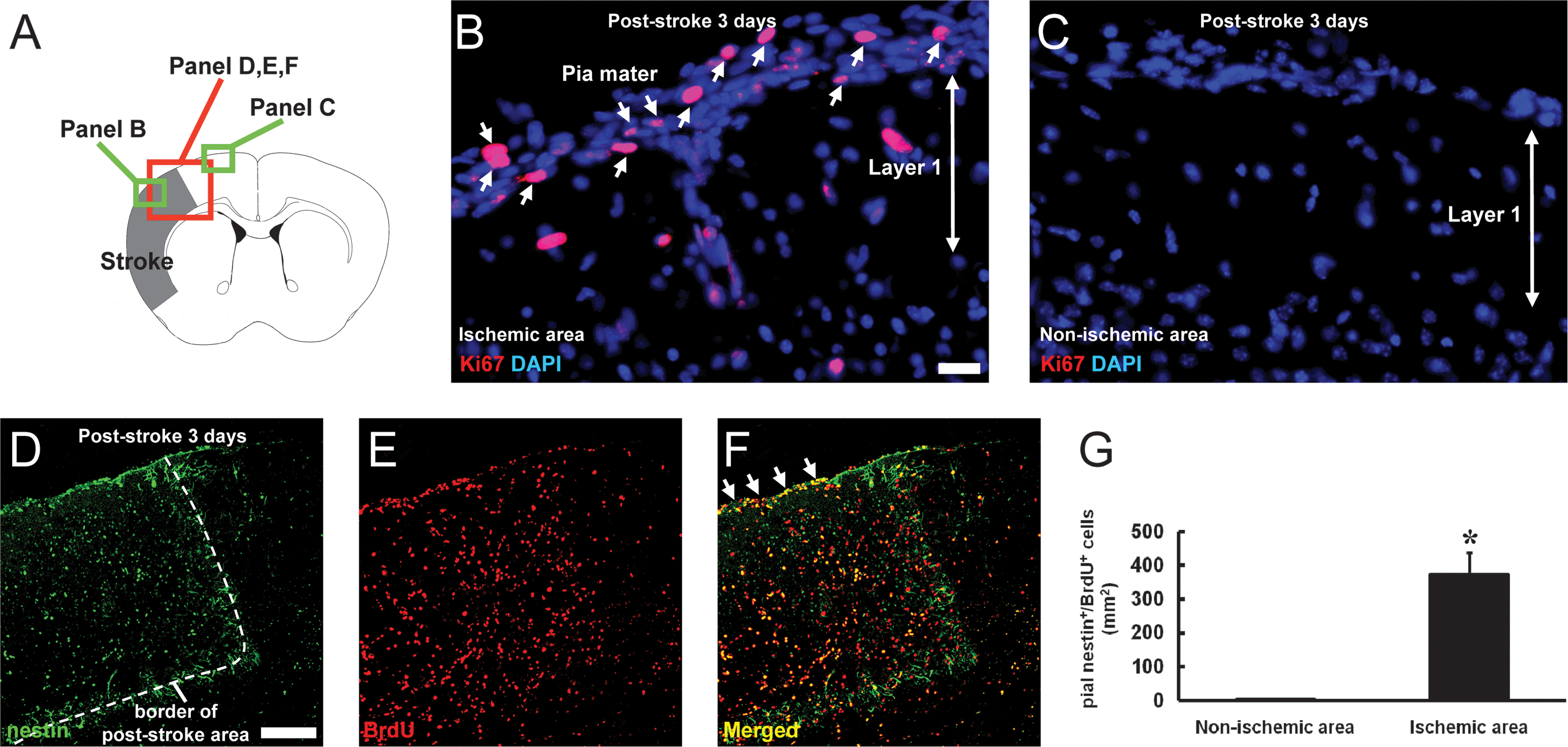
Nestin-positive cells in the poststroke pia mater proliferate after ischemia. Brain sections on poststroke day 3
We also investigated the time course of nestin expression after stroke (Fig. 1E–H). On poststroke day 1, some weakly immunoreactive cells were observed in the poststroke pia mater and cortex (Fig. 1F, Q). The number of nestin-positive cells in these areas increased on poststroke day 3 (Fig. 1G, Q). On poststroke day 7, these nestin-positive cells decreased in number (Fig. 1H, Q). These findings were consistent with our previous study [7]. Nestin-positive cells were rarely observed in the nonischemic ipsilateral pia mater or cortex for any of the ages studied (poststroke days 1–7; Fig. 1I–L, Q). Nestin-positive cells were observed in the ipsilateral SVZ (Fig. 1M–P) as early as poststroke day 1 (Fig. 1N, R), followed by an increase in number by day 3 (Fig. 1O, R). However, these cells were still restricted to the area close to SVZ (also see Fig. 1D). On poststroke day 7, some nestin-positive cells migrated out of SVZ, but they did not reach the poststroke cortex at that time (Fig. 1P, R). These findings are consistent with our previous study showing that GFP-labeled SVZ cells migrate out of the dorsal horn of the rostral migratory stream on poststroke day 7, but they do not yet reach the poststroke cortex at that time [5].
Pial nestin-positive cells spread into the cortical parenchyma as microvascular pericytes/perivascular cells
Until now, our data indicate that nestin-positive cells appear in the poststroke pia mater [5]. Pia mater is histologically continuous with the cortical parenchyma along the blood vessels [20,21]. Therefore, we further investigated the relationship between the pial nestin-positive cells and parenchymal blood vessels by double staining for nestin and CD31, a marker for ECs. We observed that nestin-positive cells in the poststroke pia mater developed in proximity to CD31-positive ECs and that some pial nestin-positive cells spread into the cortex through cortical layer I along the blood vessels (Fig. 3A–C). Whole-mount specimens [22] of poststroke pia mater also showed that nestin-positive cells were present near CD31-positive ECs (Fig. 3D–F). We further investigated whether nestin-positive cells developing in these regions coexpressed NG2, a marker of microvascular pericytes [23]. Double labeling for nestin and NG2 showed that some nestin-positive cells in the poststroke pia mater (∼60%) as well as those in the poststroke cortex [5] coexpressed NG2 lying next to ECs (Fig. 3G–I). Further, a large proportion of nestin-positive pial cells (∼95%) coexpressed PDGFRβ, another marker for microvascular pericytes [24] (Fig. 3J–L).
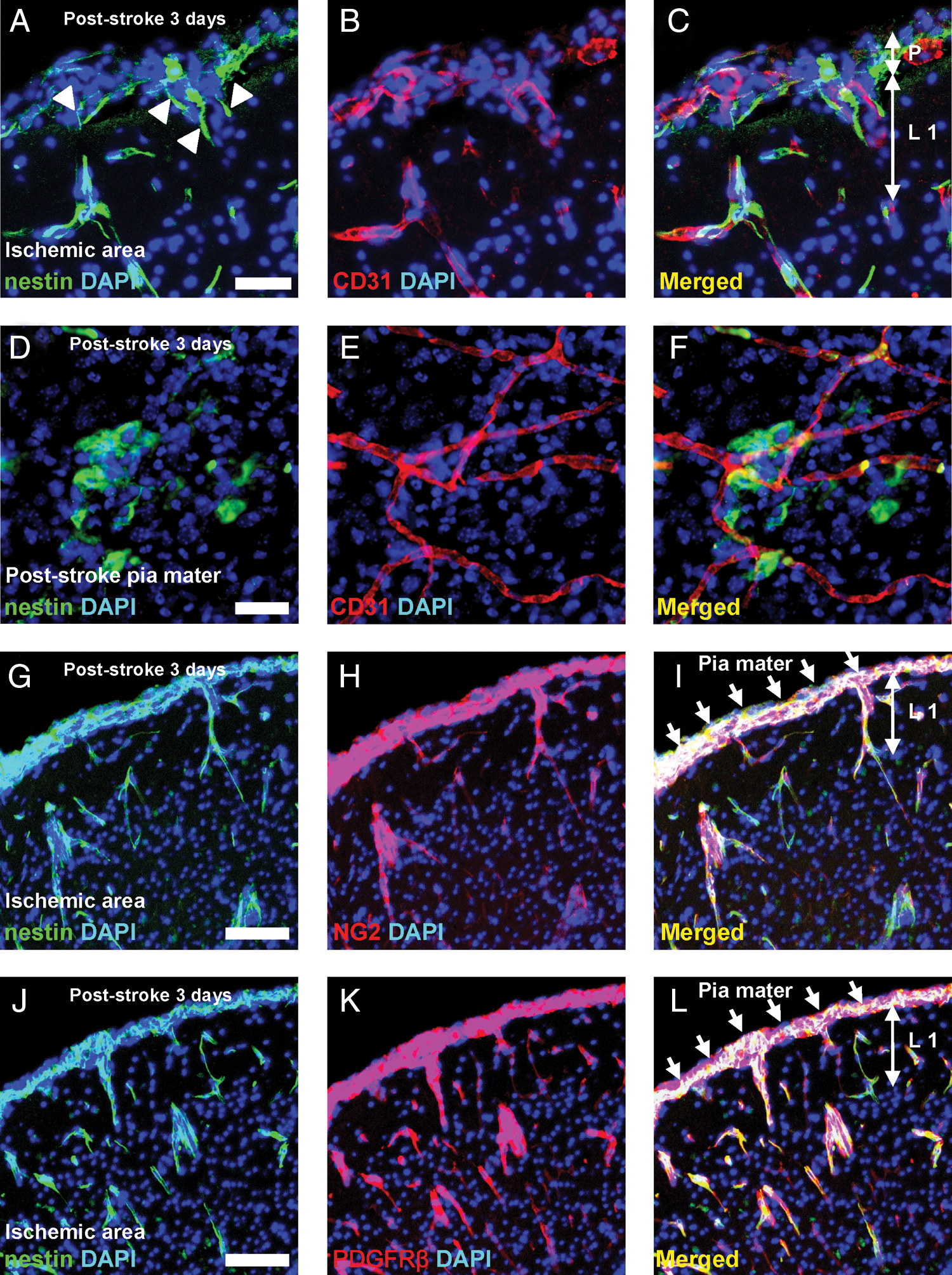
Nestin-positive cells observed in the perivascular area by microvascular pericyte marker expression. Coronal brain sections on poststroke day 3 showed that nestin-positive cells developed in close association with CD31-positive ECs from the poststroke pia mater to the cortex [nestin
Pial nestin-positive cells have NSPCs activity to generate neurons
Previous studies have demonstrated that nestin/NG2-positive CNS microvascular pericytes [25] and PDGFRβ-expressing pericytes [26] have NSPC activity and can generate neurons. In addition, we have recently shown that nestin-positive iNSPCs develop in the perivascular region of the postischemic cortex in proximity to ECs [6,9]. Based on these findings, we investigated whether pial nestin-positive cells developing near ECs have NSPC activity. To study this, we cultured pial cells as mentioned above. Neurosphere-like cell clusters appeared on day 4 of the incubation (Fig. 4A). Their number then gradually increased at later time points (day 4, 1.2±0.4; day 10, 6.0±1.2; day 20, 9.5±2.5). Thirty days after incubation, the number of cell clusters reached ∼14 (13.6±3.8). In contrast, we did not obtain neurosphere-like cell clusters from nonischemic pia mater. The pial iNSPCs also possessed self-renewal capacity after being incubated in the same medium at a clonal density of 5 cells/μL [5,6], but cluster formation was limited to between 3 and 5 cell passages. These observations were consistent with the traits of the iNSPCs obtained from the poststroke cortex [5] and suggest that pial iNSPCs are more likely to include neural progenitors than neural stem cells (NSCs) similar to neurospheres derived from other brain subregions such as the hippocampus [27] and subcortical white matter [28]. Immunohistochemistry revealed that virtually almost all cells forming spheres expressed nestin, and some coexpressed NG2 (Fig. 4B–D). The population of nestin- and NG2-positive cells was counted in an adherent monolayer culture [8] of pial cells. Similar to the neurospheres, ∼95% of the cells expressed nestin, and ∼60% of them expressed NG2 (Fig. 4E–G).
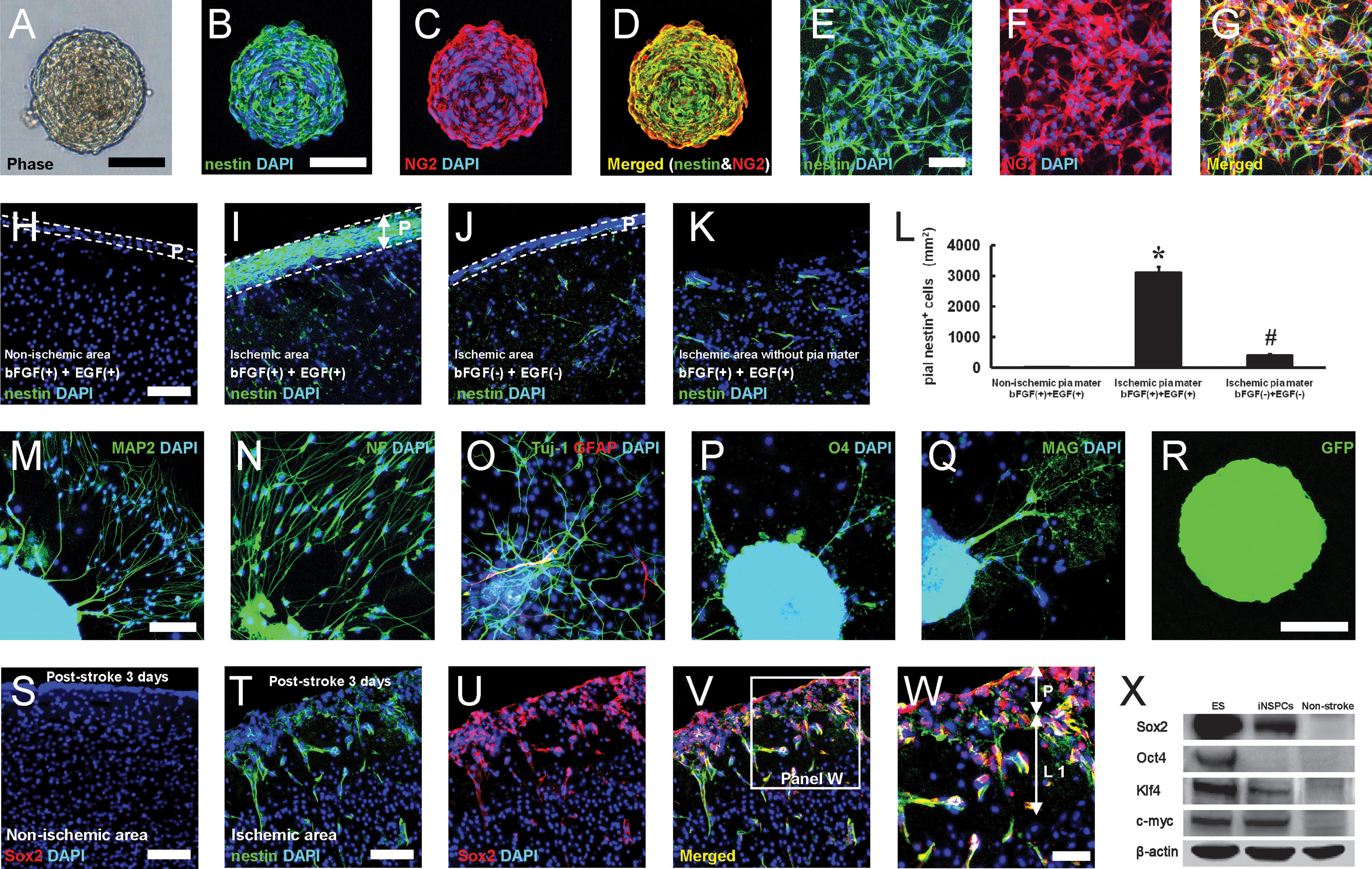
Pial nestin-positive cells show NSPC activity. Neurosphere-like cell clusters were obtained from the poststroke pia mater cultured in Dulbecco's modified Eagle's medium (DMEM) containing bFGF and EGF
We used organotypic cortical slice cultures to obtain additional independent evidence that nestin-positive cells isolated from poststroke pia mater could expand in response to bFGF and EGF. Although nestin-positive cells were rarely observed in normal pia mater/cortex (data not shown) or the nonischemic ipsilateral pia mater (Fig. 4H, L; 3.2±1.3 cells/mm2) and cortex (Fig. 4H), many nestin-positive cells were observed in the poststroke pia mater (Fig. 4I, L; 3100.8±189.4 cells/mm2) and cortex (Fig. 4I). However, in the absence of bFGF and EGF, fewer nestin-positive cells were observed, even in the poststroke pia mater (Fig. 4J, L; 393.8±55.5 cells/mm2) and cortex (Fig. 4J). After removing the poststroke pia mater, few nestin-positive cells were observed in the cortex (Fig. 4K). These observations strongly suggest that nestin-positive cells in the poststroke pia mater proliferate in response to bFGF and EGF and they are responsible for forming neurosphere-like cell clusters in vitro.
Double staining for nestin and CD31 was performed because nestin has also been shown to be expressed in cells with endothelial phenotype [29]. Pial nestin-positive cells proliferated in response to bFGF and EGF but did not coexpress CD31 (Supplementary Fig. S1A–C; Supplementary Data are available online at
Next, we investigated whether pial nestin-positive neurosphere-like cell clusters could differentiate into mature neural cells, including neurons, astrocytes, and oligodendrocytes. Cell clusters were plated on poly-L-lysine-coated glass coverslips for 7 days under conditions conducive to differentiation [6]. After differentiation, the cells expressed neuronal (Fig. 4M: MAP2, 4N: neurofilament, 4O: Tuj-1), astrocyte, (Fig. 4O: GFAP), and oligodendrocyte (Fig. 4P: O4, 4Q: MAG) markers. Cell population analysis for differentiated spheres showed that these cells predominantly differentiated into neuronal cells and oligodendrocytes rather than astrocytes (Tuj-1-positive neuronal cells 36.7%±6.6%; GFAP-positive astrocytes 4.8%±2.3%; and O4-positive oligodendrocytes 30.0%±6.1%). To confirm that these nestin-positive cells originated from pial cells, pia mater was labeled with a GFP-expressing vector. Then, the poststroke pia mater isolated 3 days after stroke was incubated in DMEM containing bFGF and EGF. GFP-positive neurospheres were obtained (Fig. 4R) and were found to have the same properties as shown above (Fig. 4A–D, M–Q).
To further explore the stem-cell nature of pial iNSPCs in vivo, we examined Sox2 expression, a marker of persistent NSPCs [30]. Sox2 expression was observed in nestin-positive cells located in SVZ and ependyma (data not shown), as described previously [31,32]. In contrast, Sox2-positive cells were rarely detectable in the nonischemic pia mater/cortex (Fig. 4S). However, Sox2-positive cells were observed in the poststroke pia mater and cortex, and some of them (∼60%) coexpressed nestin (Fig. 4T–W). Although the pial cells revealed several NSPC characteristics following ischemia, recent studies have demonstrated that NSPCs derived from other regions (e.g., SVZ-derived NSCs [33] and cortex-derived NSPCs [5]) also possess pluripotent/undifferentiated cell markers. Thus, these markers, including Sox2, Oct4, Klf4, and c-myc, which are expressed in induced pluripotent stem (iPS) cells [34], were examined by western blot analysis. As a control, we used ES cells [5], which express these 4 factors as described previously [34], in our western blot analysis. Although none of these 4 factors were observed in nonischemic pia mater, Sox2, Klf4, and c-myc, but not Oct4, expression was observed in pial iNSPCs (Fig. 4X), consistent with previous studies [5,33].
Pial nestin-positive iNSPCs originate, at least in part, from brain pericytes
Thus far, our data indicate that pial nestin-positive iNSPCs spread into the cortical parenchyma as perivascular cells and express pericyte markers. These results strongly suggest that these cells originate, at least in part, from microvascular pericytes. To confirm this, we isolated PDGFRβ-positive cells from poststroke mice by magnetic cell sorting [26]. Immunohistochemistry showed that PDGFRβ-positive cells were positive for nestin (Fig. 5A–C) and NG2 (Fig. 5D–F). These cells formed cell clusters in the medium that promoted neurosphere formation, as mentioned above (Fig. 5G). After differentiation (Fig. 5H), they generated MAP2- and Tuj-1-positive neurons (Fig. 5I, J), GFAP-positive astrocytes (Fig. 5J), and O4-positive oligodendrocytes (Fig. 5K). Because brain pericytes are considered to have a neural crest origin, similar to the pia mater [35,36], we further investigated whether the pial iNSPCs possess neural crest markers in addition to those of pericytes. The pial nestin-positive iNSPCs largely (∼95%) expressed Sox10, a neural crest marker (Fig. 5L–N). Polymerase chain reaction (PCR) analysis showed that iNSPCs, which expressed a NSPC marker (nestin) and pericytic markers (NG2 and PDGFRβ), displayed various neural crest markers such as Snail, Slug, Twist, and Sox9 (Fig. 5O), although these markers were rarely observed in ES cells, as described previously [37]. These observations support our present finding that pial iNSPCs are derived from brain pericytes, which are neural crest derivatives [36].
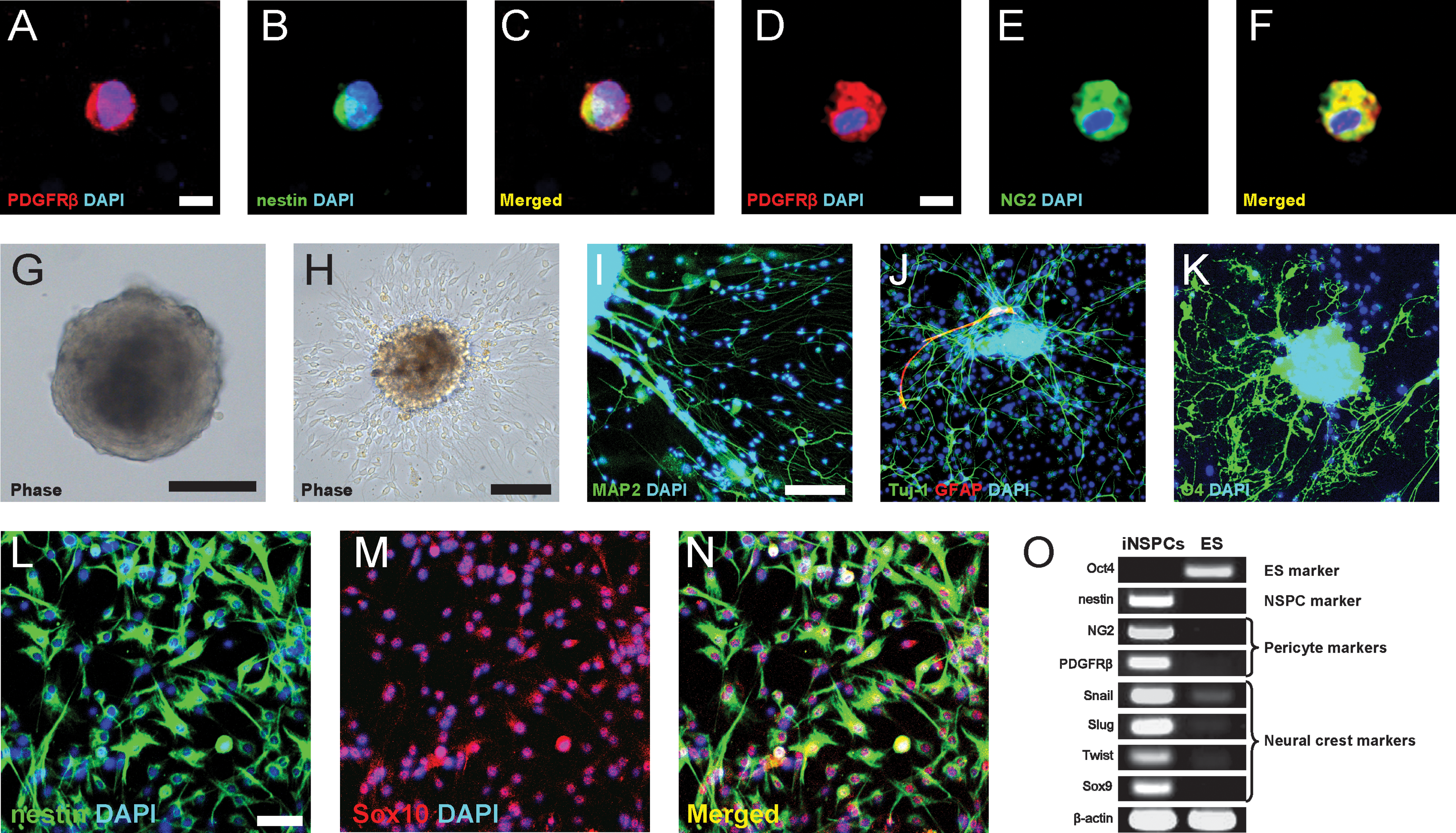
Pial nestin-positive iNSPCs are, at least in part, derived from brain pericytes. PDGFRβ-positive cells were isolated by MACS. Immunohistochemistry showed that these cells coexpressed nestin [PDGFRβ
Comparison of pial iNSPCs to other types of CNS stem/progenitor cells
Radial glia cells are derived from neuroepithelial cells during development and give rise to several types of CNS stem/progenitor cells such as SVZ astrocytes [38,39], ependymal cells [40,41], and oligodendrocyte precursor cells (OPCs) [42,43] in adults [44]. Although pial iNSPCs expressed pericyte markers, such as NG2 and PDGFRβ, the precise characteristics of NG2-expressing cells in poststroke pia mater remain unclear. Previous studies have reported that NG2 is also expressed in resident glia/glial precursors with NSPC activity [45,46], and in ependymal cells (a specialized glia), which acquire NSPC activity after CNS injury [41]. Anatomically, the pia mater spreads into the ventricular wall where it connects with ependymal cells to form the choroid plexus, which produces cerebrospinal fluid [14]. Ependymal cells were originally considered to be the resident stem cell population in the perivascular zone [47]. Although it is controversial whether ependymal cells have NSPC activity or not, recent studies confirm that ependymal cells do not play a role in adult neurogenesis under normal conditions, but they do possess NSPC activity and can differentiate into neurons, astrocytes, and oligodendrocytes in response to CNS injuries including ischemic stroke [40,41]. Further, ependymal cells strongly express PDGFRα [48] and are penetrated by the apical process of NSCs in SVZ (type B cells), which express GFAP [38,39]. Based on these findings, we investigated whether pial iNSPCs are similar to periventricular NSPCs such as ependymal cells and/or SVZ NSCs (astrocytes). Consistent with previous studies, PDGFRα (Fig. 6A) and GFAP (Fig. 6B) expression was confirmed in the ependymal [48] and SVZ cells [38,39], respectively. PDGFRα-positive cells were present in the poststroke pia mater, and some nestin-positive cells in the poststroke pia mater co-expressed PDGFRα (Fig. 6C–E). As expected, nestin and GFAP double-positive reactive astrocyte-like cells were observed in the peri-stroke area (Fig. 7A, C, F) [49,50]. However, cells neither in the poststroke pia mater nor in the poststroke cortex expressed GFAP (Figs. 6F and 7A, B, D). In contrast, many GFAP-positive astrocytes were observed within the nonischemic ipsilateral pia mater/cortex (Fig. 7A, C, E). Similarly, most (90%) of the cultured pial nestin-positive cells were PDGFRα positive (Fig. 6G–I), but did not express GFAP (Fig. 6J–L), indicating that pial iNSPCs are not reactive astrocytes. Because the pial NSPCs did not express GFAP in vivo (Figs. 6F and 7A, B, D) and differentiated into astrocytes at a very low frequency (Figs. 4M–Q and 5I–K), it is also possible that they are different from SVZ NSCs [38,39]. Thus, we further compared several astrocyte markers in pial neurosphere-like cell clusters with those in SVZ-derived neurospheres. Similar to the in vivo findings, pial neurosphere-like cell clusters largely expressed nestin but not GFAP (Fig. 6M–O). In contrast, virtually almost all cells forming SVZ-derived neurospheres expressed GFAP rather than nestin (Fig. 6P–R). In accordance with these findings, reverse transcription (RT)-PCR analysis showed that GFAP expression in the pial spheres was much weaker than that in SVZ-derived neurospheres. In addition, AQP4, which is predominantly expressed in astrocyte foot processes [51], was not expressed in pia-derived spheres, although it was rich in SVZ-derived neurospheres (Fig. 6S; GFAP, 176 bp; AQP4, 278 bp; GAPDH, 180 bp). These observations are consistent with previous studies showing that ependyma with NSPC activity display weak [41] or no GFAP expression [47]. Taken together, these results suggest that pial iNSPCs may have similar characteristics to ependymal cells rather than to SVZ astrocytes. To confirm this, we further investigated whether the poststroke pial cells have the structures of lipid droplets, microvilli, and cilia, which ependyma cells possess [47,48]. These structures were confirmed in the ependyma by electron microscopy (Fig. 6T). However, microvilli-like structures were not observed in cells located in the poststroke pia mater (Fig. 6U). These findings indicate that pial iNSPCs do not have completely identical characteristics to those of ependymal cells.
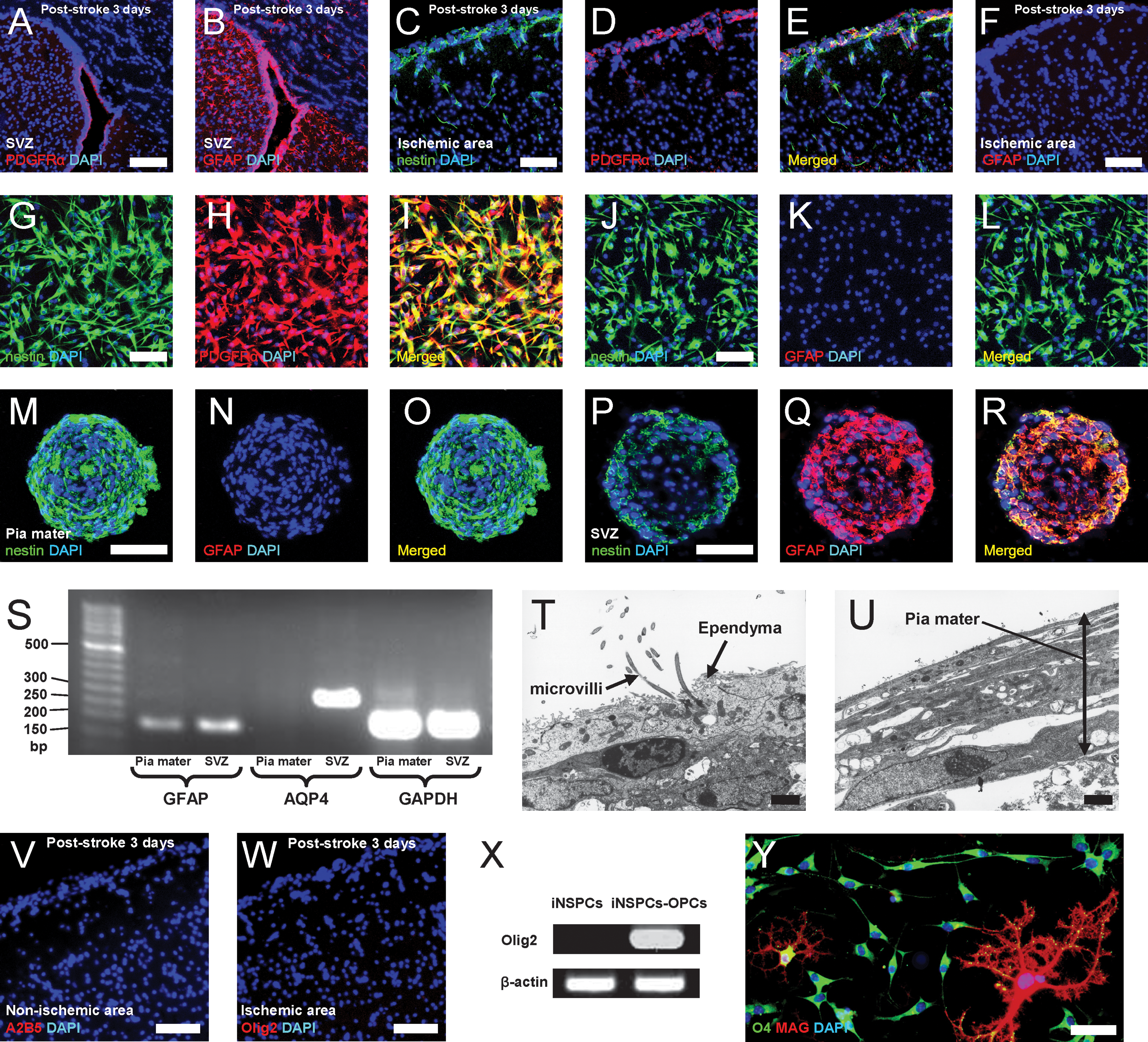
Comparison of pial iNSPCs to previously reported CNS stem/progenitor cells. The traits of pial iNSPCs were analyzed and compared with those of SVZ astrocytes, ependymal cells, and OPCs
Pial nestin-positive cells do not express the astrocytic marker GFAP. Immunohistochemistry on poststroke day 3
Because OPCs also express NG2 [52] and PDGFRα [53], and A2B5-positive OPCs have NSPC activity [42,43], we further investigated whether pial nestin-positive cells might have OPC characteristics with multipotent NSPC activity. Immunohistochemistry on poststroke day 3 showed that A2B5-positive (Fig. 6V) and Olig2-positive (another OPC marker) cells [54] (Fig. 6W) were not present in poststroke pia mater or cortex. However, following incubation in OPC-promoting medium [55], Olig2 expression was induced in pial iNSPCs (iNSPCs-OPCs) (Fig. 6X) and almost all these cells (∼98%) induced from the iNSPCs differentiated into O4- and/or MAG-positive oligodendrocytes (Fig. 6Y). These observations indicate that pial nestin-positive iNSPCs are different from reported multipotent OPCs [42,43], although they partially co-express OPC markers.
Moreover, pial iNSPCs did not express GAD67 (data not shown), which is a marker of layer I progenitors [12]. Schematic representation for the fate of iNSPCs (Fig. 8) and their distinct expression of markers are summarized in Table 1.
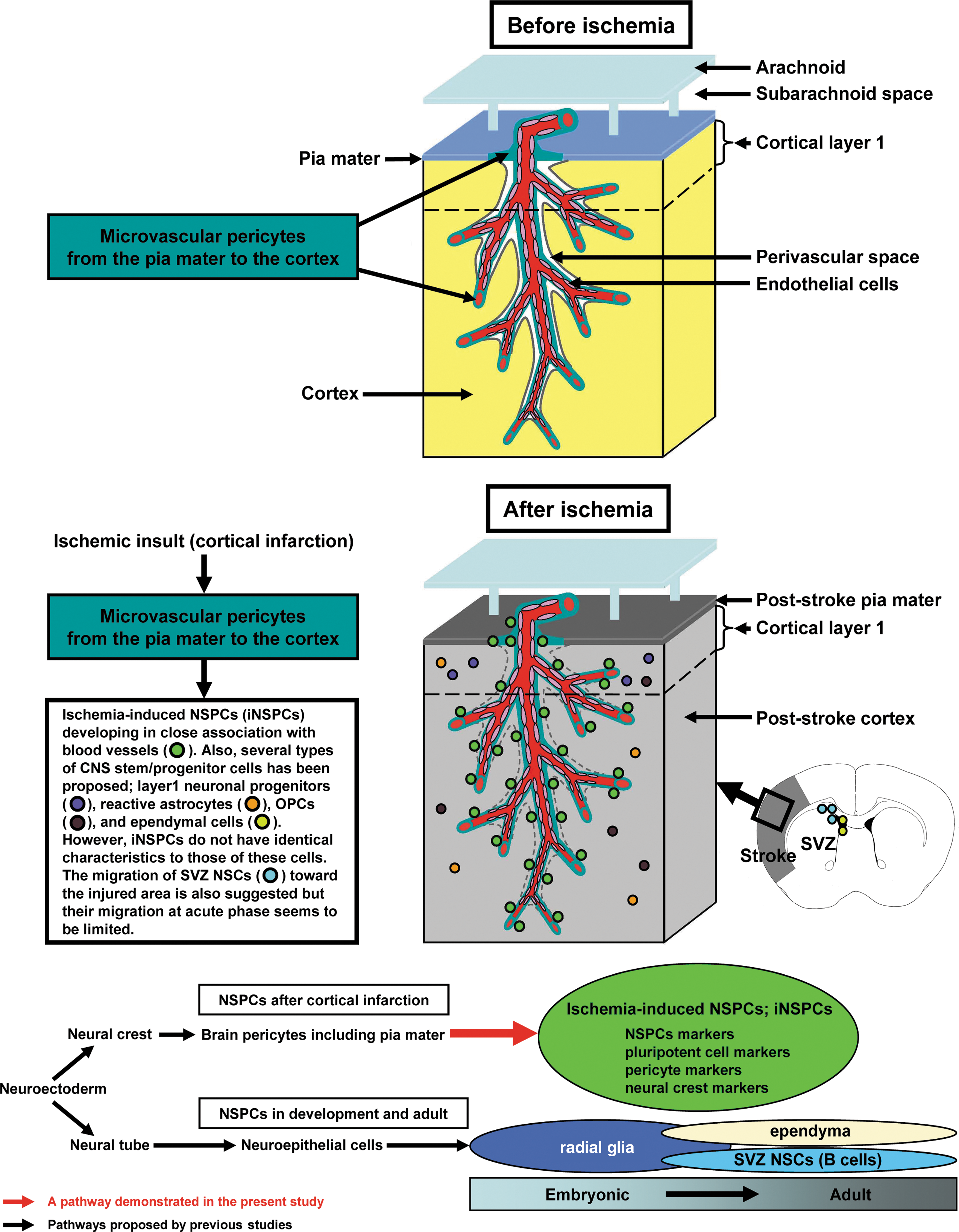
Schematic representation for the fate of iNSPCs following cortical infarction. Following cortical infarction, pial iNSPCs, which are presumably derived from the neural crest-brain pericyte lineage, develop in the poststroke pia mater in close association with blood vessels throughout the brains, including the cortex. These cells reveal markers for NSPCs and pluripotent/undifferentiated cells in addition to those for pericytes and the neural crest. Pial iNSPCs do not have identical characteristics to those of other types of proposed CNS stem/progenitor cells, such as SVZ NSCs (astrocytes), ependymal cells, OPCs, reactive astrocytes, or layer I progenitors. NSCs, neural stem cells.
+, positive expression; ND, nondetectable; iNSPC, ischemia-induced neural stem/progenitor cells; SVZ, subventricular zone; NSC, neural stem cells; OPC, oligodendrocyte precursor cells; GFAP, glial fibrillary acidic protein; PDGFRα, platelet-derived growth factor receptor alpha; PDGFRβ, platelet-derived growth factor receptor beta.
Discussion
We demonstrated for the first time that nestin-positive iNSPCs, which are potential sources of neurons driving cortical neurogenesis, developed in the poststroke pia mater. In addition, we showed that they developed in the cortex as has been described previously [5,6]. Mignone and colleagues have shown that in developing transgenic nestin-GFP embryos GFP expression was evident as early as day 7 of embryonic development (e7). At e8, a GFP signal was seen predominantly in the neural plate, and by e10 intense GFP fluorescence was seen throughout the neuroepithelium. At e10 to e12, GFP signals marked the entire thickness of the cerebral wall, but GFP expression became weaker near the pial surface and stronger in the ventricular zones starting from e12. Finally, in the adult brain, GFP was selectively expressed in SVZ and the subgranular zone (SGZ) within the dentate gyrus of the hippocampus in areas related to continuous neurogenesis [56]. However, after brain injury/ischemia, nestin-positive cells and/or nestin-positive NSPCs were observed in nonconventional neurogenic zones such as the cortex [5,9] and striatum [57], although they were rarely observed in the absence of brain ischemia/injury [5]. These findings suggest that NSPC activation and neuronal homeostasis are maintained under physiological conditions, at least in part, in specific brain regions (such as SVZ and SGZ). However, after brain injury/ischemia, it would appear that regional NSPCs are mobilized to accelerate tissue repair in adult mammalian brains, in addition to activating neurogenesis in SVZ and SVG [58].
Previous studies have demonstrated that stem/progenitor cells in SVZ have the capacity to migrate toward injured lesions, including those in the cortex [59]. However, our previous study failed to demonstrate cell migration from SVZ to the cortex after cerebral infarction in vivo but showed that iNSPCs in the poststroke cortex originated, at least in part, from within the cerebral cortex [5]. Consistent with that study, subsequent studies have shown that NSPCs developing within and around the poststroke cortex are derived from locally activated stem/progenitor cells and not from SVZ cells [12,49], indicating that it is more likely that iNSPCs from the poststroke pia mater/cortex rather than NSCs from SVZ are involved in cortical repair after stroke, particularly from the acute phase. Previous studies have indicated that NSPCs express an astrocyte marker, GFAP, after brain injury; thus, reactive astrocytes are proposed as a potential origin of injury-iNSPCs [49]. However, we could not find GFAP-positive cells within the poststroke area itself, and isolated pial iNSPCs showed few astrocytic traits compared with SVZ NSCs, suggesting that pial iNSPCs are a distinct population with a different origin. In the present study, we further demonstrated that pial iNSPCs after cortical infarction originate, at least in part, from the microvascular pericytes/perivascular cells. Pericytes with multipotent progenitor activity have been indentified in various organs [60] in addition to CNS [25]. Although a previous study showed that pericyte-derived NSPCs can be isolated from CNS of noninjured animals [25], we could not obtain iNSPCs from nonischemic pia mater and cortex [5]. Consistent with these findings, only pericytes located within ischemic pia mater/cortex but not within nonischemic pia mater/cortex expressed NSPC markers such as nestin and Sox2. These observations suggest that ischemia/hypoxia may enhance the stem-like nature of brain pericytes, although we do not yet know which signaling and/or factors are essential for their induction. Interestingly, it has been demonstrated that some CNS pericytes originate from the neural crest [36] as well as the pia mater [35], and our current study showed that pial iNSPCs expressed various neural crest markers in addition to pericytic markers. These observations also support that pial iNSPCs originated from brain pericytes, which are neural crest derivatives, providing a novel concept that neural crest-derived cells may play a crucial role in CNS repair following cortical infarction.
It is still highly controversial whether periventricular NSPCs are SVZ astrocytes (type 2 cells), ependymal cells, or both [61]. However, our combined immunohistochemical, RT-PCR, electron microscopic, and cell culture analyses demonstrated that pial iNSPCs do not have characteristics identical to those of SVZ astrocytes or ependymal cells. Although pial nestin-positive iNSPCs expressed NG2, a pericyte marker [23], it is also expressed in resident glial cells/glial progenitors [62], which have NSPC activity after brain injury [46,63]. Combined with the present results that the pial iNSPCs expressed other glia markers such as PDGFRα, and that they have the potential to convert into OPCs under certain conditions [55], it is possible that iNSPCs may also have glial cell characteristics. However, neuron production from NG2- and/or PDGFRα-positive glial cells is rare [63,64], suggesting that iNSPCs may be different from these glial cells. The precise origin, lineage, and traits of iNSPCs warrant further investigation; nevertheless, our current study revealed a novel NSPC population.
Neuronal progenitors have been reported to be induced in layer I of the cerebral cortex in response to ischemia. Such neuronal progenitors express GAD67 but not nestin [12]. In contrast, pial iNSPCs did not express GAD67 but revealed strong nestin expression. Therefore, these neuronal progenitors may not be identical to iNSPCs that we showed in the present study. Alternatively, Bifari and colleagues have shown that novel stem/progenitor cells with neuronal differentiation potential reside in the leptomeninges during development [15]. In the present study, we further demonstrated that pial iNSPCs with neuronal differentiation potential were induced in the poststroke pia mater. These observations suggest that neuronal progenitors in the poststroke cortex including neocortical layer I may originate, in part, from pial iNSPCs that migrate toward the injured cortex in addition to NSPCs that originate in the cortex (e.g., cortical pericytes-derived iNSPCs and/or other types of proposed NSPCs such as resident glia, reactive astrocytes, and layer I progenitors).
In the present study, we found that iNSPCs developed in close association with blood vessels of the brain from the pia mater to the cortex in the poststroke area. These results suggest that environmental controls surrounding iNSPCs such as ECs, which are an important niche for NSPCs [65], and perivascular (Virchow-Robin) spaces, in which many inflammatory cells infiltrate after brain injury [21], are also essential for cortical neurogenesis after ischemic stroke. In support of these view points, our previous studies showed that adequate support of iNSPCs in the in vivo environment of the poststroke cortex (e.g., regulation of ECs [6,8] and of inflammatory cells [7]) could promote iNSPCs proliferation and enhance neurogenesis.
In conclusion, we have demonstrated that iNSPCs, which are potential sources for neocortical neurogenesis, developed in the poststroke pia mater, suggesting that pia mater that covers the entire brain cortex is an important target for cortical neurogenesis following stroke. Thus, deposition of several materials, including cell sheets [66], on poststroke pia mater may make it possible to promote cortical repair/neurogenesis without cerebral parenchymal damage by intracerebral cell transplantation in the future.
Footnotes
Acknowledgments
This work was partially supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology (21700363 to T. Nakagomi and 21500359 to T. Matsuyama), Hyogo Science and Technology Association (to T. Nakagomi), and Takeda Science Foundation, 2009 (to T. Nakagomi). We would like to thank Y. Okinaka, Y. Tanaka, and Y. Tatsumi for technical assistance, and F.M. Oeschger, C.E. Turnquist-Held, F.G. Szele, H.B. Stolp, F. García-Moreno, W.Z. Wang, C.C. Young, and E.H. Chang for helpful discussions.
Author Disclosure Statement
The authors declare that no conflict of interest exists.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.