
Abstract
Stem cell therapy is a promising therapeutic strategy for treating myocardial infarction (MI). However, it is necessary to identify ideal adult stem cells for transplantation and explore mechanisms of the transplanted cells in improving cardiac functions after MI. In this study, a population of embryonic-like stem cells (ELSCs) was isolated from rat bone marrow. The cells express pluripotent stem cell transcriptional factors and present high proliferative activity on mouse embryonic fibroblast feeder. ELSCs retain clonal expansion and may form embryoid-like bodies in soft agarose containing leukemia inhibitory factor and basic fibroblast growth factor. The cells of the embryoid-like bodies can differentiate into the cells from 3 germ layers. Under induction, the cells can differentiate into cardiomyocytes and endothelial cells. In MI models of female rats, the transplantation of preinduced ELSCs of male rats reduce scar area and improve cardiac function significantly. Comparing with marrow-derived mesenchymal stem cells and ELSCs without induction, effects of the preinduced ELSCs on myocardial repair and improvement of cardiac function are greater. Survival of the transplanted cells in the peri-infarcted and infarcted regions was examined by fluorescence in situ hybridization. Y chromosome-positive cells may differentiate toward cardiomyocytes and express cTnT and Cx43. Cx43 expression was observed at conjunction of Y chromosome-positive cells and recipient cardiomyocytes. Some Y chromosome-positive cells express CD31 and incorporate into the microvessels in the infarcted tissue. These results suggest that a population of ELSCs resides in rat bone marrow and display similar biological characteristics of ESCs. ELSCs can differentiate into cardiomyocytes and endothelial cells and contribute to cardiomyogenesis and angiogenesis in vivo. Cardiac function after MI may be significantly improved with transplantation of the preinduced ELSCs. Therefore, ELSCs are novel seed cells for stem cell transplantation in regenerative medicine.
Introduction
M
The results of current studies demonstrate that effects of the transplanted stem cells are closely related to cell category [8,9]. Embryonic stem cells (ESCs), induced pluripotent stem (iPS) cells, and pluripotent stem cells (PSCs) in adults are expected to be sources of stem cells because of their self-renewal and multi-differentiation potentiality. However, ESCs are restricted in clinical application owing to their immunogenicity and ethical issues [10 –12]. iPS cells have high risk in forming tumor and teratoma after transplantation [13,14]. The major advantage of adult stem cells is that the cells can be obtained from adult self-body so that the risk of immune reaction may be avoided and ethical issues do not exist. Therefore, adult PSCs may be an optimal source of stem cells for cell transplantation after MI. Recently, a population of very small embryonic-like stem cells (ELSCs) have isolated from bone marrow of mice, umbilical cord blood and peripheral blood in adult humans [15,16]. However, whether the very small ELSCs exist in adult rat bone marrow or not remains unclear.
Recently, a study showed that null mutations in human and mouse orthologs frequently resulted in different phenotypes [17]. Accordingly, mouse as an experimental model is invariably restricted. Comparing with mouse, rats are much closer to humans in genetic, individual development, anatomical structures, and behaviors [18]. Therefore, the rat as a model in research of the genetics, ethology, pharmacology, and pathophysiology has obvious advantages over the mouse. In the MI model, change of cardiac function of the rat with slower heart rate, larger ventricular weight, and volume is more easily evaluated than that of the mouse. Therefore, it will be of great significance to explore mechanisms of cardiac functional improvement with rat ELSCs transplantation in the MI model.
In this study, ELSCs were isolated from the bone marrow of adult rats. Proliferative property, clonal expansion, and formation of embryoid-like bodies of the cells were performed on feeder layer of mouse embryonic fibroblasts (MEFs) and in soft agarose, respectively. Biological characteristics of ELSCs were analyzed by electron microscopy, immunocytochemistry, and reverse transcription-polymerase chain reaction (RT-PCR). The differentiation of the cells in embryoid-like bodies into the cells from 3 germ layers was detected, differentiation toward cardiomyocytes and endothelial cells was induced with cytokines in vitro. Therapeutic effects of ELSCs transplantation were explored in rat MI models. Improvement of cardiac function after transplantation was evaluated by echocardiography, survival and differentiation of the cells in vivo was traced by Y chromosome fluorescence in situ hybridization and immunohistochemical staining.
Materials and Methods
Cell isolation and sorting
Eight-week-old male Sprague-Dawley (SD) rats were purchased from Slac Laboratory Animal Corporation and housed in Department of Laboratory Animal Science of Fudan University, China. The investigation was permitted by the Law of the People's Republic of China on the Protection of Wildlife, and the protocol was approved by the Institutional Animal Care Committee of Fudan University, China. The femurs and tibias of rats were cut off under sterile conditions. The bone marrow was harvested by washing with phosphate-buffered saline (PBS) containing 800 U/mL heparin. Then, suspension of the bone marrow was added on Percoll (GE Healthcare) solution at the density of 1.073. After centrifugation (25 min, 1,750 rpm) at 4°C, the fraction of the mononuclear cells was collected.
For fluorescence-activated cell sorting, the mononuclear cells were blocked with 10% goat serum, and then, the cells were incubated with mouse anti-SSEA-1 antibody (1:200; Santa Cruz Biotechnology) for 50 min at 4°C. After rinsing with PBS, the cells were incubated with goat anti-mouse IgG conjugated with FITC (1:300; Chemicon Millipore) for 30 min. After washing, the cells were resuspended with Dulbecco's modified Eagle's medium (DMEM; Gibco Invitrogen) supplemented with 2% fetal bovine serum (FBS; Gibco Invitrogen), 500 U/mL penicillin, and 0.5 mg/mL streptomycin (Beyotime). The cells incubated only with second antibody were used as control. SSEA-1+ cells were sorted by a fluorescence-activated cell sorter (FACS Aria II; BD Biosciences), and collected in DMEM containing 20% FBS, 200 U/mL penicillin, and 0.2 mg/mL streptomycin [19]. Purity of SSEA-1+ cells was examined with the FACS again.
Detection of lineage markers
The sorted SSEA-1+ cells were incubated with CD45-PE antibody (Biolegend) and Ter-119-APC antibody (eBioscience) or Gr-1-PE antibody (eBioscience) and CD11b-APC antibody (Biolegend) for 50 min at 4°C. Incubation with PBS as control was performed. Then, expression of the lineage markers of the cells was detected by a FACS.
Cell proliferation and clone formation assays
To amplify the sorted cells, a feeder layer of MEFs was prepared. MEF line CF-1 (Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences, Shanghai) was incubated in DMEM supplemented with 15% FBS. Then, the cells were collected and treated with 20 μg/mL mitomycin C (Hisun Pharmaceutical) at 37°C for 1 h. After mitomycin C solution had been removed, the cells were seeded in a culture dish at the density of 1×106 cells/mL and incubated in a 37°C and 5% CO2 incubator (Sanyon Electric). The cells were used as a feeder layer when they spread into a monolayer [20]. The sorted SSEA-1+ cells were seeded on a feeder layer of MEFs and incubated in DMEM supplemented with 10% FBS, 1% nonessential amino acids, 1,000 U/mL leukemia inhibitory factor (LIF; Chemicon Millipore), and 20 ng/mL basic fibroblast growth factor (bFGF; Gibco Invitrogen). LIF and bFGF were supplemented every 24 h, the medium was replaced every 2–3 days. At intervals of 4–7 days, colonies of SSEA-1+ cells were selected for passage. The SSEA-1+ colonies were mechanically detached into small clumps with a micropipette tip and seeded on a new MEF feeder layer. The cells seeded in a culture dish without feeder layer were used as a contrast experiment.
To evaluate the clone formation of SSEA-1+ cells, 0.6% agarose gel containing 1× DMEM, 20% FBS, 2,000 U/mL LIF, and 40 ng/mL bFGF was prepared in a 24-well plate. Density of SSEA-1+ cells was adjusted to 1×103 cells/mL, 330 cells/mL, 110 cells/mL, and 37 cells/mL, respectively, with 0.3% agarose containing 1× DMEM and 20% FBS. Then, suspension of the cells (200 μL/well) was added on the agarose gel and incubated in an incubator for 2 weeks.
Electron microscopy
For scanning electron microscopy, the cells incubated on MEF feeder layer for 1 day and 7 days were fixed with 2.5% glutaraldehyde overnight at 4°C, respectively, and underwent a graded ethanol series and final twice rinsing in 100% ethanol for 15 min. Then, the cells went through another graded series to replace ethanol with hexamethyldisilazane. After being dried in air, the specimens were coated with carbon, and examined under a scanning electron microscope (SEM; FEI QUANTA200, Philips) [21].
For transmission electron microscopy, SSEA-1+ cells were fixed with 2.5% glutaraldehyde at 4°C, and then fixed with 1% osmium tetroxide. After being dehydrated with gradient alcohol, the cells were soaked with anhydrous acetone and Jpurr resin, and embedded with Jpurr resin. Ultrathin sections were gained with Reichert-ultracut E ultrathin microtome (Leica), and stained with 3% uranyl acetate and lead citrate. The ultrastructures of the cells were viewed by a CM120 transmission electron microscope [22].
Immunocytochemistry
For evaluating pluripotency of SSEA-1+ cells, the cells were added on coverslips and fixed with acetone and methanol (1:1) for 20 min at −20°C. After treatment with goat serum, rabbit anti-Oct4 antibody (1:200; Abcam), rabbit anti-Nanog antibody (1:200; Chemicon Millipore), and rabbit anti-Sox2 antibody (1:200; Sigma) were added on coverslips, respectively, and incubated at 4°C overnight. After incubation at 37°C for 2 h, the cells were incubated with goat anti-rabbit IgG conjugated with TRITC (1:300; Jackson ImmunoResearch Laboratories) for 30 min [23]. The nuclei were counterstained with 4', 6-Diamidino-2-phenylindole (DAPI, 1:1,000; Sigma). The cells were mounted with glycerophosphate and viewed under a fluorescent microscope (Olympus Corporation).
To examine expression of stem cell antigen-1 (Sca-1), SSEA-1+ cells were incubated with rabbit anti-Sca-1 antibody (1:200; eBioscience) at 37°C for 1 h. Subsequently, the cells were incubated with goat anti-rabbit IgG conjugated with TRITC (Jackson) at 37°C for 30 min. The nuclei were counterstained with DAPI (1:1,000; Sigma). The cells were viewed with an LSM 510 confocal lazer scanning microscope (Carl Zeiss MicroImaging GmbH) [24].
RT-PCR analysis
Expression of PSC transcriptional factor mRNAs Oct-4, Nanog, Rex1, Sox2, and Fgf4 in the cells was analyzed by RT-PCR. Total RNA of rat SSEA-1+ cells, mouse ESC line D3 cells (ES-D3 cells, Chinese Academy of Sciences, Shanghai), and rat marrow-derived mesenchymal stem cells (MMSCs) [25,26] was extracted, respectively. Primers (Sangon Biotech) used for amplification are listed in Table 1. The mRNA levels were normalized by using Gapdh as a housekeeping gene. PCR reactions were performed under the following conditions: 1 cycle at 45°C for 30 min and 94°C for 5 min, followed by 35 cycles at 94°C for 30 s, 60°C for 30 s and 72°C for 1kb/min, and 1 cycle at 72°C for 5 min. The PCR products were size-fractionated with 1.5% agarose (Takara Bio Inc.) gel electrophoresis [4]. The images were analyzed by software (Furi Science and Technology).
Tumorigenicity assay
For investigating possibility of tumorigenesis and teratoma formation, 0.5 mL suspension of SSEA-1+ cells at the density of 2×106 cells/mL was subcutaneouly injected in the inguinal region on both sides of 10 mice (5 male and 5 female) with severe combined immunodeficiency (SCID; Shanghai Slac Laboratory Animal Corporation). The mice were sacrificed to examine formation of neoplasms at 4 weeks after the injection.
Differentiation assay in vitro
To evaluate potency of differentiation of the cells in the embryoid-like bodies toward the cells from 3 germ layers, expression of ectodermal, mesodermal, and endodermal markers was examined by immunocytochemistry. Nestin, Brachyury, and E-cadherin were used for ectodermal, mesodermal, and endodermal markers, respectively. The cells in the embryoid-like bodies were picked out with a pipette and incubated in a 24-well plate with DMEM containing 5% FBS. At 4 weeks after incubation, the cells were collected and added on slides. The cells were labeled by rabbit anti-Nestin antibody (1:200; Chemicon Millipore), mouse anti-Brachyury antibody (1:200; Santa Cruz), and rabbit anti-E-cadherin antibody (1:200; Abcam), respectively. Subsequently, the cells were incubated with goat anti-mouse IgG conjugated with FITC (1:300; Chemicon Millipore) or goat anti-rabbit IgG conjugated with TRITC (1:300; Jackson) and viewed with a fluorescent microscope (Olympus Corporation).
For induction of differentiation toward cardiomyocytes, SSEA-1+ ELSCs of 2–3 passage were seeded on coverslips, and then incubated in DMEM supplemented with 10% FBS, 10 ng/mL bone morphogenetic protein 2 (BMP-2; PeproTech), 10 ng/mL insulin-like growth factor 1 (IGF-1; GeneWay, Biotech), and 10 ng/mL bFGF. The cytokines were supplemented daily, and the medium was replaced every 2–3 days. The cells without induction by cytokines were used as control. At 2 weeks after induction, expression of mRNAs of cardiac transcriptional factors GATA-4 (Gata4) and Nkx2-5 and mRNAs of cardiac specific proteins cTnI (Tnni3k), cTnT (Tnnt2), and Cx43 (Gja1) was analyzed by RT-PCR. The cardiomyocytes isolated from rat hearts were used as positive control. Cardiomyocytes were prepared according to the method described by Egorova et al. [27]. The primer sequences were listed in Table 1. PCR reactions were performed with the protocol as just described. At 4 weeks after induction, expression of cardiac specific proteins in the differentiated cells was examined with mouse anti-cTnT antibody (1:200; Santa Cruz) and rabbit anti-Cx43 antibody (1:200; Abcam) in double staining. After incubation with secondary antibodies conjugated with FITC and TRITC, the cells were viewed with a fluorescent microscope.
For inducing differentiation toward endothelial cells, ELSCs of 2–3 passage were seeded on coverslips, and incubated in DMEM supplemented with 10% FBS, 10 ng/mL vascular endothelial growth factor (VEGF; R&D Systems), 10 ng/mL bFGF, and 10 ng/mL transforming growth factor beta (TGF-β; R&D Systems). The cytokines were supplemented daily, and the medium was replaced every 2–3 days. The cells incubated with the medium without cytokines were used as control. At 2 weeks after induction, expression of Pecam1, Flt1, and Flk1 was analyzed by RT-PCR. The endothelial cells isolated from cerebral microvasculatures of rats by using the method previously described by Lu [28] were used as positive control. At 4 weeks after induction, the expression of endothelial markers in the differentiated cells was examined with mouse anti-Von Willebrand factor (vWF) antibody (1:200; Abcam) and rabbit anti-CD31 antibody (1:200; BD Biosciences) in double staining.
Establishing of MI models and cell transplantation
Sixty adult female SD rats (240–260 g) were anesthetized with ketamine (60 mg/kg) and xylazine (5–10 mg/kg) by a peritoneal injection. After endotracheal intubation and ventilation with a rodent ventilator (Harvard Apparatus), the heart was exposed through a 2 cm long left lateral thoracotomy. Left anterior descending coronary artery (LADCA) was ligated [4,29]. Successful MI was determined by observing a pale discoloration of the myocardium and a high T-wave on the electrocardiogram. After LADCA ligation, the chest of the rats was sutured. 2×104 U penicillin G benzathine was intramuscularly injected daily for 3 days after the operation.
Nine rats died after LADCA ligation, and 5 rats were kicked out because of the lack of an ideal echocardiogram. At 1 week after MI, the remaining 46 rats were randomly divided into control, MMSC, ELSC, and induced-ELSC groups. In induced-ELSC group, the cells were preinduced with 10 ng/mL BMP-2, 10 ng/mL IGF-1, and 10 ng/mL bFGF for 24 h before transplantation. The rats were anesthetized, and their hearts were exposed as just mentioned. The cells (1×105) were suspended with 80 μL PBS and injected into the border of the infarcted myocardium through a micro-syringe at 4 spots (20 μL per spot). In control group, 80 μL PBS was injected [4,30]. 2×104 U penicillin G benzathine was intramuscularly injected daily for 3 days after the operation.
Echocardiography
To evaluate the improvement of cardiac function, changes of echocardiograms were recorded with an ultrasonocardiograph (Visual Sonics) under anesthetization before MI, at 1 week after MI (baseline echocardiogram), and 4 weeks after cell transplantation. After adequate two-dimensional images were obtained, the M-mode cursor was positioned to the parasternal long axis view at the level papillary muscles. Left venticular (LV) end-diastolic diameter (LVEDD) and LV end-systolic diameter (LVESD) were measured from at least 3 consecutive cardiac cycles. LV end-diastolic volume (LVEDV), LV end-systolic volume (LVESV), Index of ejection fraction (EF, LVEDV-LVESV/LVEDV×100%), and fractional shortening (FS, LVEDD-LVESD/LVEDD×100%) were measured for examination of systolic function [4]. In ideal medels, EF was <50% at 1 week ligation of LADCA.
Morphological analysis
After echocardiography, the rats were sacrificed, and their hearts were rapidly excised. The tissue masses of upper, central, and lower infarcted regions were prepared and fixed in 4% paraformaldehyde for 36 h. After being dehydrated in sucrose solution, the masses were embedded in Tissue-Tek OCT Compound (Sakura Finetek) and frozen at −80°C. Frozen sections of 5 μm thickness were prepared by a Leica CM 1900 microtome (Leica Microsystems). Nine discontinuous sections were selected from each model and stained with Masson's trichrome stain (Sigma). The blue regions indicate scar tissues [25]. Scar area was defined as the percentage of blue region in whole myocardium of the cross-sectional area in LV wall by using computer-based planimetry with ImageJ analysis software (National Institutes of Health).
Survival and differentiation of the transplanted cells
For investigating the survival and differentiation of the transplanted cells within infarcted myocardium, Y chromosome specific gene of the cells was detected by fluorescence in situ hybridization [4,31]. The frozen sections were postfixed and immersed in 70% formamide to denature the fixed chromosome specimens. Then, the sections were dehydrated through gradient ethanol and dried in air. After being labeled with biotin, the Y chromosome specific probes (Shan Jing, Shanghai) were denatured in Hybrisol VII (Qbiogene) at 95°C for 5 min; then, the sections were incubated with the denatured probe at 42°C overnight. The sections were washed twice with 2× saline sodium citrate buffer and then incubated with streptavidin-FITC or streptavidin-Cy3 (Biolegend) at 37°C for 30 min. After treatment with blocking solution, the sections were incubated with rabbit anti-cTnT antibody (1:200; Santa Cruz), rabbit anti-Cx43 antibody, or mouse anti-CD31 antibody (1:200; BD Biosciences). Then, the sections were incubated with goat anti-rabbit IgG conjugated with TRITC or goat anti-mouse IgG conjugated with FITC. The nuclei were counterstained with DAPI. The Y chromosome-positive cells expressing cTnT, Cx43, or CD31 were counted under the fluorescence microscope, respectively.
Statistical analysis
Data are expressed as the means±standard deviation. To statistically analyze the data, we performed Student's t-test and 1-way analysis of variance with Scheffe's post hoc multiple-comparison analysis. A value of P<0.05 was considered significant, and a value of P<0.01 was considered very significant.
Results
Identification of Lin-SSEA-1+ cells
After the procedures of FACS, the region R1 of lymphocytes was selected for gating of control sample. The same region was selected for gating of SSEA-1 labeling sample. The result of FACS shows that SSEA-1+ cells in the mononuclear cells were 0.015% (Fig. 1A, B). SSEA-1+ cells did not express lineage markers such as CD45, Ter-119, CD11b, and Gr-1 (Fig. 1C, D). In the sorted SSEA-1+ cells, percentage of the positive cells was 97.2±1.7.
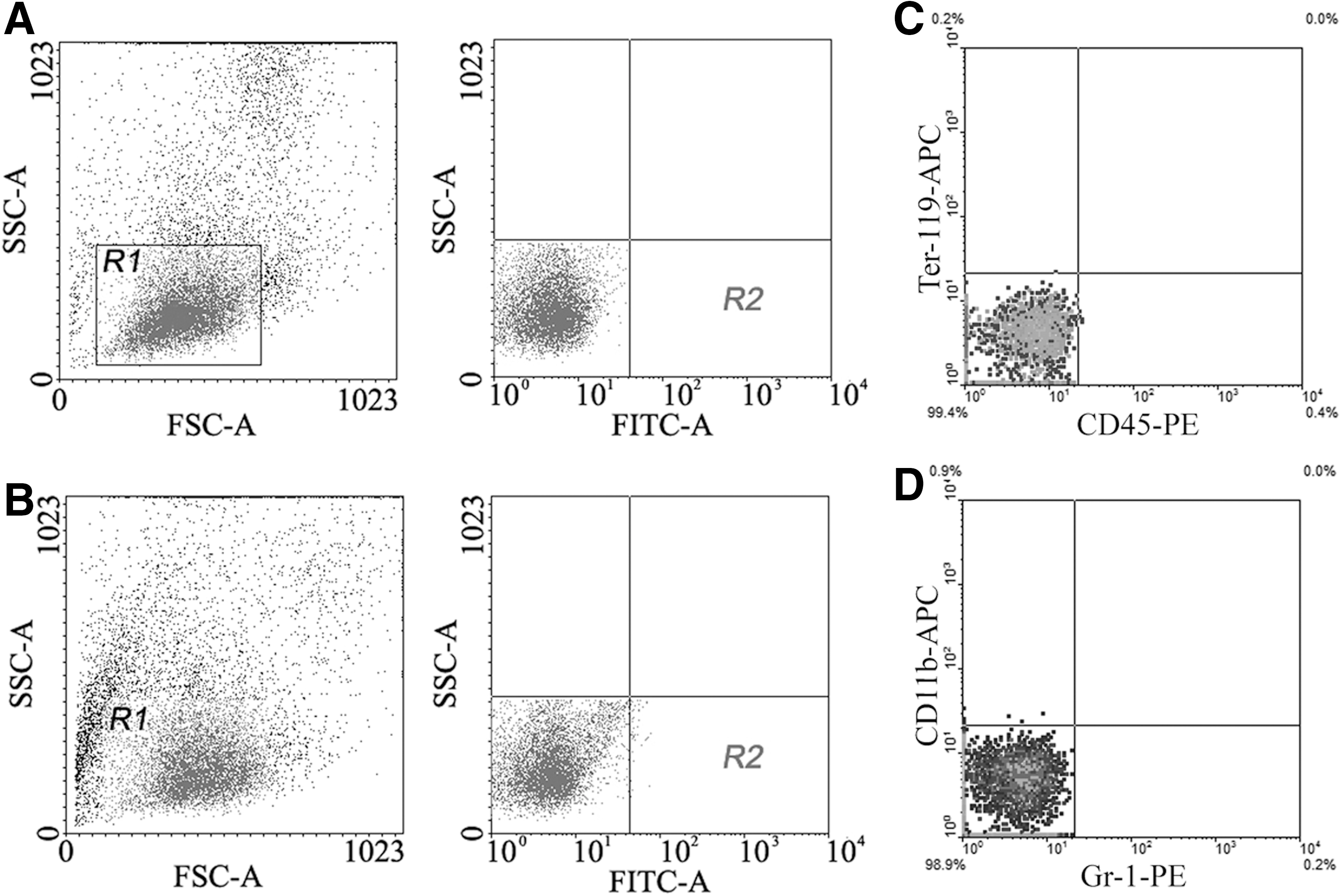
Sorting of SSEA-1+ cells from mononuclear cells of rat bone marrow by FACS.
Proliferative ability and morphological features
Diameter of SSEA-1+ cells is 4–5 μm. The cell contains less cytoplasm, the nucleus is large, and the ratio of the nucleus to cytoplasm is very high. The organelles are not rich (Fig. 2A). At 1 day after incubation on MEF feeder layer, SSEA-1+ cells were round, and singly and dispersedly distributed. Microvilli on SSEA-1+ cells were rich. At 7 days, the cells proliferated into clusters, and the feeder cells under the cluster were pushed away (Fig. 2B). When SSEA-1+ cells were incubated in 0.3% agarose, the cells were dispersed at 1 day. At 1 week after incubation, the cells proliferated into clusters. The diameter of the most clusters is 30–50 μm. At 2 weeks after incubation, the cells are grown to embryoid-like bodies (Fig. 2C). In the culture dish without 0.3% agarose, proliferation of SSEA-1+ cells was not obvious.
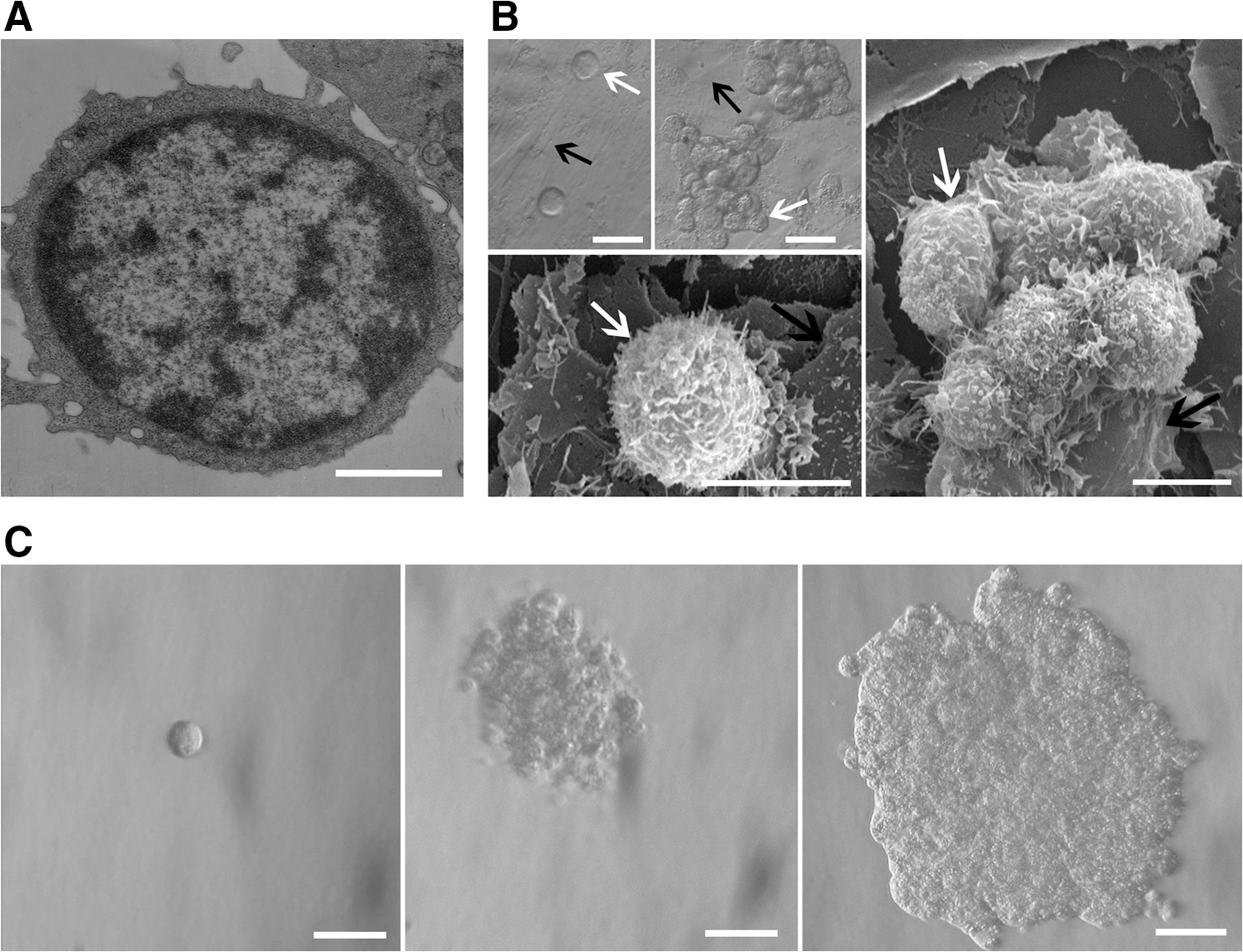
Ultrastructural features of SSEA-1+ cells and proliferation of the cells on MEF feeder.
Expression of PSC transcriptional factors
SSEA-1+ cells expressed PSC transcriptional factors Oct4, Nanog, and Sox2. These transcriptional factors were expressed mainly in the nuclei, while SSEA-1 was expressed mostly on the cell membrane (Fig. 3A). The SSEA-1+ cells also expressed Sca-1 (Fig. 3A, the lowest row of panels). RT-PCR analysis demonstrated that SSEA-1+ cells expressed PSC transcriptional factor mRNAs Oct4, Nanog, Rex1, Sox2, and Fgf4 (Fig. 3B). The expression levels of Oct4, Nanog, and Rex1 in ELSCs were lower than those in ES-D3 cells, but the expression levels of Sox2 and Fgf4 were higher than that in ES-D3 cells. The expression levels of Oct-4, Nanog, Rex1, Sox2, and Fgf4 in ELSCs and ES-D3 cells were significantly higher than those in MMSCs (Fig. 3C).

Expression of PSC transcriptional factors in SSEA-1+ cells and 3 germ layer markers on the cells of embryoid-like bodies.
Expression of 3 germ layer markers
At 1 week after incubation in DMEM without LIF and bFGF, morphological changes of the cells of the embryoid-like bodies were not obvious. At 3–4 weeks after incubation, the cells became flat, oval, or spindle. Expression of ectodermal marker Nestin, mesoderm marker Brachyury, and endoderm marker E-cadherin on the cells could be detected by immunocytochemistry (Fig. 3D). Neither tumor nor teratoma was observed in the inguinal region of SCID mice at 4 weeks after transplantation of ELSCs.
Differentiation toward cardiomyocytes and endothelial cells in vitro
After induction with BMP-2, IGF-1, and bFGF for 2 weeks, SSEA-1+ cells became spindle and arranged in parallel. At 3–4 weeks after induction, the cells changed into short columnar shape and conjuncted each other. RT-PCR analysis revealed that the cells expressed Gata4, Nkx2-5, Tnni3k, Tnnt2, and Gja1 at 2 weeks after induction (Fig. 4A). Statistical analysis showed that the expression levels of cardiac early transcriptional factor mRNAs Gata4 and Nkx2-5, and cardiac specific protein mRNAs Tnni3k, Tnnt2, and Gja1 in the induced cells were lower than the levels of the rat cardiomyocytes (Fig. 4B). At 4 weeks after induction, the differentiated cells expressed cTnT and Cx43 (Fig. 4C). In control group, shape of the cells did not obviously change, and expression of cTnT and Cx43 was not observed (Fig. 4C). After induction with VEGF, bFGF, and TGF-β for 2 weeks, RT-PCR analysis demonstrated that the cells expressed Pecam1, Flt1, and Flk1. The expression level of Pecam1 in the induced cells is lower than that in microvascular endothelial cells of rats, but the expression levels of Flt1 and Flk1 in the cells after induction are higher (Fig. 4D, E). At 4 weeks after induction, the differentiated cells became polygon and expressed vWF and CD31 (Fig. 4F). Expression of vWF and CD31 was not observed in the cells without induction by cytokines (Fig. 4F).
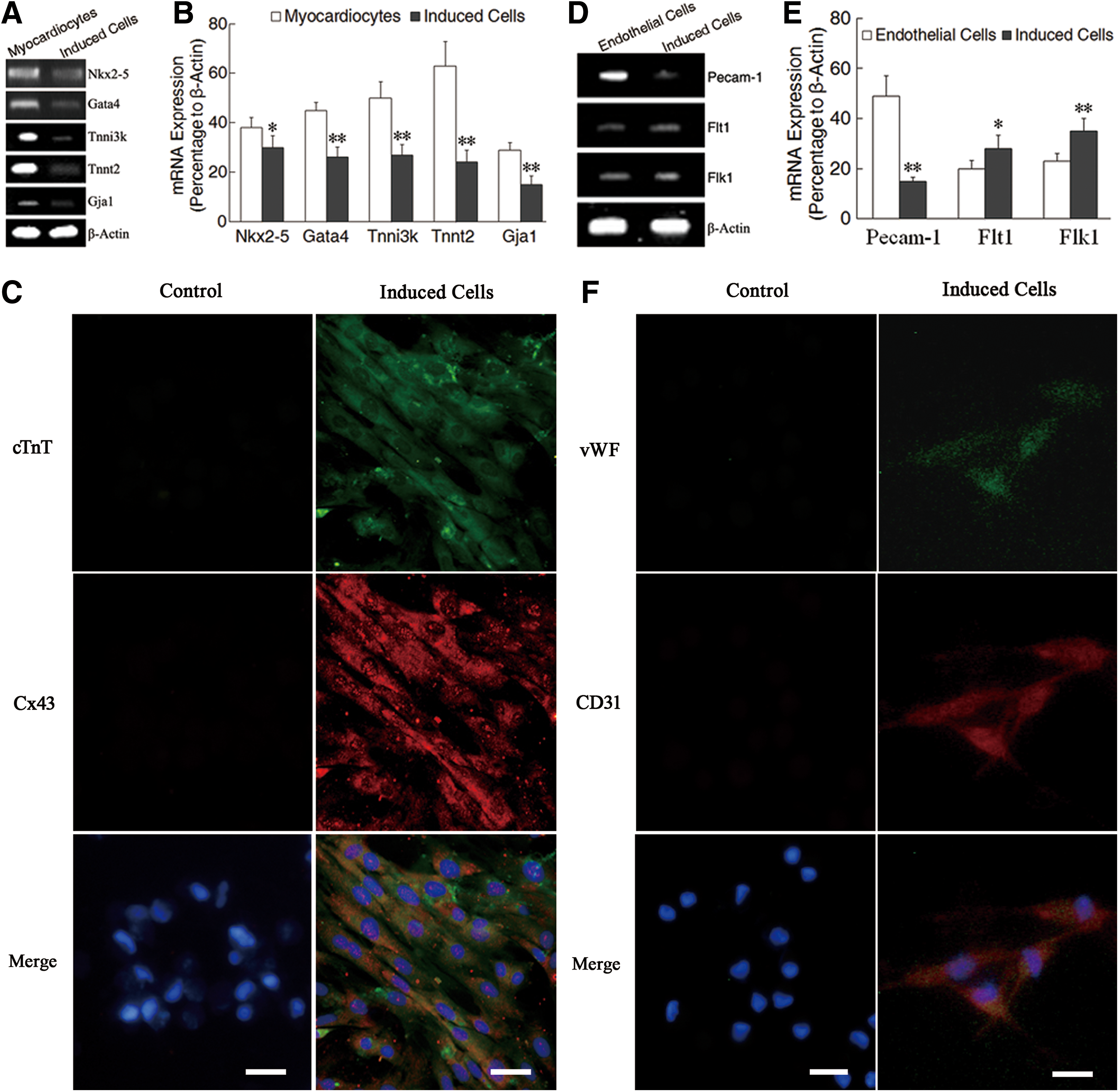
Differentiation of ELSCs toward cardiomyocytes and endothelial cells after induction with cytokines in vitro.
Improvement of LV function after cell transplantation
Serial echocardiographic analysis indicated that heart function in all rats was severely compromised after LADCA occlusion for 1 week. Significant functional loss continued to the fourth week in control group. The hearts transplanted stem cells demonstrated an increase in heart function during the same time interval (Fig. 5A). The echocardiograms showed that LVEDD and LVESD in induced-ELSC group were smaller than that in baseline, and control, MMSC and ELSC groups. LVESD in MMSC and ELSC groups was also smaller than that in control group (Fig. 5B, C). In control group, LVEDV and LVESV obviously increased at 4 weeks after MI. Comparing with baseline, and control, MMSC and ELSC groups, LVEDV and LVESV in induced-ELSC group significantly decreased. In MMSC and ELSC groups, LVEDV and LVESV also decreased as compared with control group (Fig. 5D, E). EF and FS significantly increased in MMSC, ELSC, and induced-ELSC groups comparing with control group, and EF in induced-ELSC group also increased comparing with MMSC group (Fig. 5F, G).
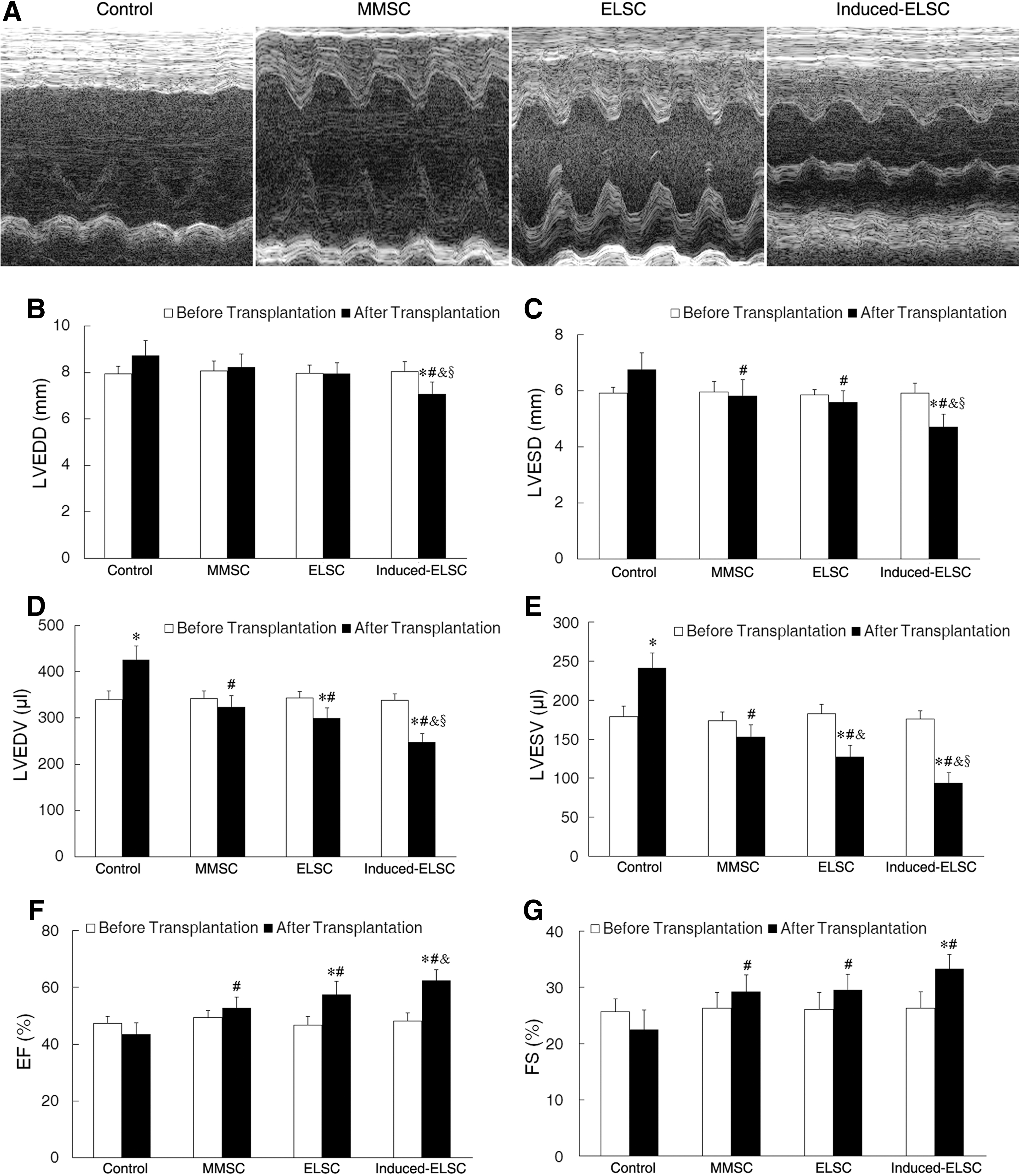
Improvement of cardiac function at 4 weeks after cell transplantation.
Histological changes after cell transplantation
Masson's trichrome staining showed that myocardial tissue in the infracted region was mainly replaced by fibrous tissue in control group at 4 weeks after MI. In groups of cell transplantation, the regenerated myocardium in the infarcted region could be observed. Cardiomyocytes in the fibrous tissue obviously increased in induced-ELSC group (Fig. 6A). The percentage of scar area in control, MMSC, ELSC, and induced-ELSC groups was 20.12±2.91, 15.81±1.82, 13.53±1.59, and 8.89±1.25, respectively. Scar area in induced-ELSC group significantly decreased comparing with control, MMSC, and ELSC groups. Scar area in MMSC and ELSC groups also decreased comparing with control (Fig. 7).
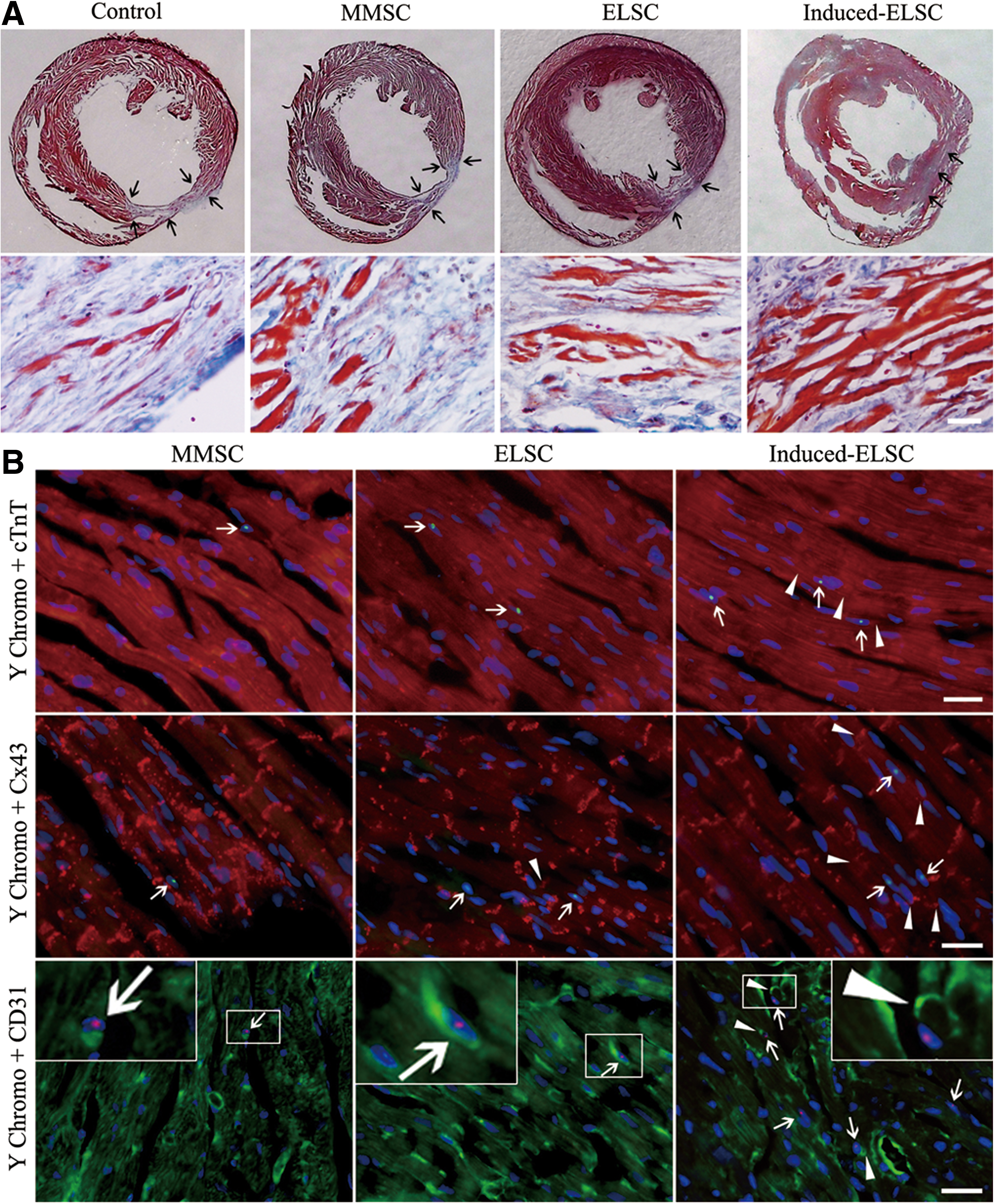
Scar area, collagen deposition, and survival and differentiation of the cells at 4 weeks after transplantation.
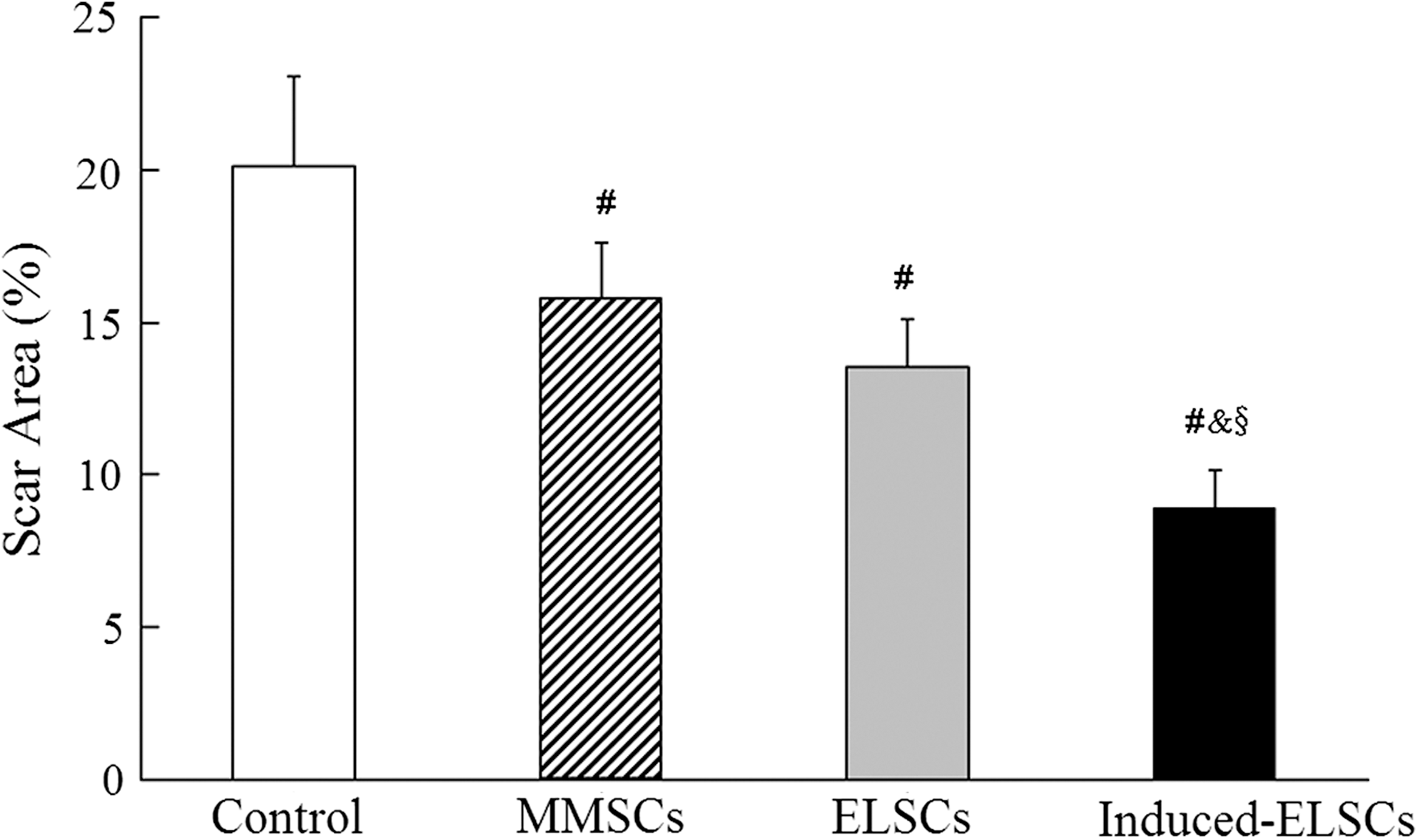
Quantity analysis of scar areas after cell transplantation. Scar area in induced-ELSC group significantly decreases comparing with control, MMSC, and ELSC groups. Scar areas in MMSC and ELSC groups also decrease comparing with the control group. # P, & P, and § P<0.01. n=8 (control group), 10 (MMSC group), 9 (ELSC group), and 11 (induced-ELSC group).
Differentiation of the transplanted cells in vivo
At 4 weeks after cell transplantation, Y chromosome fluorescence in situ hybridization demonstrated that Y chromosome-positive cells were observed in the peri-infarcted and infarcted regions. There are more Y chromosome-positive cells in induced-ELSC group comparing with MMSC and ELSC groups, and most Y chromosome-positive cells express cTnT and Cx43 (Fig. 6B). Expression of cTnT and Cx43 in the Y chromosome-positive cells indicates that transplanted ELSCs undergo cardiomyocyte differentiation in the peri-infarcted and infarcted regions. The differentiated cells are parallel in arrangement with recipient cardiomyocytes. Cx43 expression was observed at conjunction of Y chromosome-positive cells and recipient cardiomyocytes in induced-ELSC group (Fig. 6B). The result indicates that gap junction may be established between the differentiated cells and recipient cardiomyocytes.
Angiogenesis in the peri-infarcted and infarcted regions was determined by the number of CD31+ microvessels at 4 weeks after transplantation. Y chromosome fluorescence in situ hybridization demonstrated that some transplanted cells expressed CD31, and density of microvessels significantly increase in induced-ELSC group comparing with MMSC and ELSC groups. Y chromosome-positive cells expressing CD31 were observed on the wall of some microvessels in induced-ELSC group (Fig. 6B, the lowest row of panels). The results imply that the transplanted stem cells undergo endothelial differentiation and directly participate in angiogenesis in the peri-infarcted and infarcted regions.
Discussion
In this study, a population of ELSCs was isolated from the bone marrow of adult rats. The cells are small (4–5 μm), the nucleus is large, and the cytoplasm is less. Being similar with ESCs, ELSCs on feeder layer proliferate significantly, have the ability of clone formation, and may organize embryoid-like bodies in soft agarose containing LIF and bFGF. The cells of the embryoid-like bodies can differentiate into the cells from 3 germ layers in vitro. However, neither tumor nor teratoma was observed after ELSC transplantation in SCID mice. The origin of ELSCs remains unclear. Kucia et al. [32] and De Miguel et al. [33] postulated that VSEL stem cells of mice were direct descendants of the germ lineage. Ratajczak et al. [34] hypothesized that the VSEL stem cells were epiblast cells or primitive germ cells deposited into peripheral tissues. In view of the expression that Sox2 in rat ELSCs is stronger than that in ESCs and no teratoma is observed after transplantation of the cells in this study, we presume that ELSCs are descendants of epiblast cells migrating to bone marrow during development. Self-renewal or proliferation ability is one of the characteristics of stem cells and an important basis for application of the cells. ELSCs display high proliferative capacity on feeder layer of MEFs and retain clonal expansion. The cells may form embryoid-like bodies when they are incubated in soft agarose containing LIF and bFGF. However, ELSCs fail to proliferate without feeder layer or in agarose without cytokines. The proliferative features and multi-differentiation potentiality of rat ELSCs were in accordance with those of ESCs. Therefore, ELSCs are expected to become an optimal source of seed cells for stem cell transplantation therapy of MI.
Rats are much closer to humans in the genetic, individual development, anatomical structures, and behaviors. Recently, rats as the experimental animals were given more attention. Buehr et al. [35] and Li et al. [36] have captured authentic ESCs from rat blastocysts. Geurts et al. [37] have established knockout rats by microinjection of zinc-finger nucleases into embryo. These results have great significance in the targeted genetic manipulation and in providing an ideal model for tissue repair and regeneration with rats. Success isolation of ELSCs from adult rats opens up a new field for research of stem cells and MI therapy with ELSC transplantation.
The ELSCs could differentiate into cardiomyocytes and endothelial cells after induction with cytokines in vitro. In female rats with MI, transplantation of the preinduced ELSCs demonstrated that cardiac function obviously increased in EF and FS, significantly decreased in the diameter and volume of the left ventricle. Comparing with MMSC and uninduced-ELSC transplantation, function of the infarcted hearts was significantly improved by transplantation of the preinduced ELSCs. The results indicate that ELSCs, as a PSC, undergo lineage submitting process just as ESCs before differentiation toward cardiomyocytes is demanded [38]. Since a ischemic microenvironment hinders differentiation of the transplanted cells [39], selecting suitable stem cell for transplantation or effectively improving the ischemic microenvironment is important with elevating effectiveness of transplantation. Preinduction of ELSCs with BMP-2, IGF-1, and bFGF before transplantation conduces to promote the lineage submitting process of ELSCs toward cardiovascular precursors to accelerate differentiation of the preinduced ELSCs toward cardiomyocytes and endothelial cells.
Myocardial differentiation and angiogenesis of the transplanted cells in the infarcted tissue may be critical for cardiac repair and improvement of cardiac function. Many studies show that improvement of cardiac function after stem cell transplantation is implemented by the mechanism of cell paracrine [40,41]. Moreover, Guo et al. [4] suggested that the transplantation of marrow-derived cardiac stem cells in fibrin promotes cardiomyogenesis and angiogenesis. In this study, we observed that the preinduced ELSCs express cTnT and Cx43 after transplantation. Especially, Cx43 expression was observed at conjunction of Y chromosome-positive cells and recipient cardiomyocytes The result indicates that the transplanted cells can differentiate into cardiomyocytes and form gap connection with recipient cardiomyocytes. This is an important structural basis for synchronizing contraction of transplanted cells with recipient myocardium. On the other hand, angiogenesis is important for improving cardiac function. It may be evaluated by a number of CD31+ microvessels [42,43]. In our experiment, Y chromosome-positive cells expressing CD31 are observed in peri-infarcted and infarcted regions. After transplantation of the preinduced ELSCs, Y chromosome-positive cells expressing CD31 distribute on the wall of microvessels. The result implies that the transplanted cells may differentiate into endothelial cells and directly participate in angiogenesis. Cardiomyogenesis and angiogenesis in the infracted myocardium may be performed by differentiation of the transplanted cells toward cardiomyocytes and endothelial cells and paracrine effect of the cells.
In summary, rat marrow-derived ELSCs represent characteristics of ESCs and have no risk of tumorigenicity. The cells may differentiate into cardiomyocytes and endothelial cells after induction with cytokines. Transplantation with the preinduced ELSCs improves cardiac function after MI by promoting cardiomyogenesis and angiogenesis. ELSCs may be applied as optimal seed cells for regenerative medicine. Due to biological dominances, ELSCs from rats are more suitable for investigating cellular and molecular mechanisms in regenerative medicine. Recently, effectiveness of improvement of cardiac functions in MI after stem cell transplantation is still not desirable. Serious problems are that survival and differentiation of the transplanted cells in the local environment are poor. Transplantation with ideal stem cells is beneficial for survival and differentiation of the cells in myocardium repair.
Footnotes
Acknowledgments
The authors appreciate the team of Department of Laboratory Animal Science (Shanghai Medical School of Fudan University, Shanghai, China) for establishment of MI models. This study was supported by grants from Natural Science Foundation of China 30470883 and 30971674 and the Research Fund for the Doctoral Program of Higher Education of China 200802460044.
Author Disclosure Statement
No competing financial interests exist.