
Abstract
Human embryonic stem cell-derived cardiomyocytes (hESC-CMs) provide the unique opportunity to study the very early development of the human heart. The aim of this study was to investigate the effect of calcium and beta-adrenergic stimulation on the contractile properties of early hESC-CMs. Beating clusters containing hESC-CMs were co-cultured in vitro with noncontractile slices of neonatal murine ventricles. After 5–7 days, when beating clusters had integrated morphologically into the damaged tissue, isometric force measurements were performed during spontaneous beating as well as during electrical field stimulation. Spontaneous beating stopped when extracellular calcium ([Ca2+]ec) was removed or after administration of the Ca2+ channel blocker nifedipine. During field stimulation at a constant rate, the developed force increased with incremental concentrations of [Ca2+]ec. During spontaneous beating, rising [Ca2+]ec increased beating rate and developed force up to a [Ca2+]ec of 2.5 mM. When [Ca2+]ec was increased further, spontaneous beating rate decreased, whereas the developed force continued to increase. The beta-adrenergic agonist isoproterenol induced a dose-dependent increase of the frequency of spontaneous beating; however, it did not significantly change the developed force during spontaneous contractions or during electrical stimulation at a constant rate. Force developed by early hESC-CMs depends on [Ca2+]ec and on the L-type Ca2+ channel. The lack of an inotropic reaction despite a pronounced chronotropic response after beta-adrenergic stimulation most likely indicates immaturity of the sarcoplasmic reticulum. For cell-replacement strategies, further maturation of cardiac cells has to be achieved either in vitro before or in vivo after transplantation.
Introduction
D
It is assumed that human ESC-derived cardiomyocytes (hESC-CMs) recapitulate early human development in vitro. Therefore, hESC-CMs provide a unique opportunity to study cardiac function during very early developmental stages [1].
In addition, hESC-CMs are already tested in preclinical studies as a cell source for regenerative medicine (see Refs. [2 –4] for review). Currently, it is not known which developmental stage of cardiomyocytes might be optimally suited for cell transplantation. While adult cardiomyocytes are characterized by a more advanced differentiation, early stage cardiomyocytes are characterized by a higher proliferation potential [5] and would be far easier to obtain in large numbers.
HESC-CMs are spontaneously active [6], and it was already shown that chronotropy of this spontaneous activity is hormonally modulated [7]. In contrast to hormonal modifications of the beating rate, up to now, hormonal modulation of the contractile force has only been measured indirectly using, for example, measurements of cell shortening [8 –10]. Optical methods estimate contractility indirectly by measuring shortening of cells or sarcomere length of isolated unloaded cells. Though there is some consistency in data from unloaded cell-contraction experiments, externally loaded force development and shortening are known to be related but not equivalent indices of contraction. Loaded force development remains the more relevant parameter [11]. To our knowledge, this is the first report on hormonal regulation of contraction that is not based on surrogate markers of force generation but on data obtained by isometric force measurements.
Previously, we have already demonstrated the existence of a length-tension relationship for loaded isometric contractions of early hESC-CMs [12]. Current knowledge on the mechanisms that determine contractile force during the very early cardiac development is still fragmentary [13 –15]. In the adult heart, hormonal signals, that is, primarily muscarinergic and beta-adrenergic signals transmitted by the autonomous nervous system, are known important regulators of contractility (see Ref. [16] for review). The goal of this study was to characterize the regulation of early hESC-CMs with regard to their probably most relevant function, the development of force.
The aim of this study was to extend our knowledge on the fundamental properties of early hESC-CMs and to delineate the effects of changes of the extracellular calcium ([Ca2+]ec) concentration, a blockade of the Ca2+ channel by nifedipine (NIF) and beta-adrenergic stimulation on the contractile properties of these cells.
Materials and Methods
Culture of hESC
Spontaneously beating cardiac clusters were generated from the hESC cell line H1 (Wicell; passage numbers between 19 and 63). This cell line was imported according to German law and after approval by the Robert Koch Institute, Germany (No. AZ 1710-79-1-4-2). Maintenance of pluripotency and differentiation was accomplished as previously described [17]. Briefly, H1 colonies were cultivated for ∼1 week on CF1 mouse feeder cell layers that have been inactivated by mitomycin C (Serva;
First, beating areas could be observed after 5 days of co-culturing with END-2 cells. HESCs were cultured in knockout Dulbecco's modified Eagle's medium (Invitrogen;
Preparation and simulated myocardial ischemia of ventricular slices
Experiments were carried out according to the Principles of Laboratory Animal care (NIH publication No. 86-23, revised 1985) as well as the guidelines of the animal welfare committee of the University of Cologne (approval no. 50.203.2-K04/06). Preparation and subsequent irreversible injury of ventricular slices from neonatal murine hearts was performed as previously described [12]. Briefly, neonatal SV129 mice were sacrificed at postnatal day 3–4. Ventricles were embedded in 4% low melting agarose (Roth) and sectioned with a vibratome (Leica Microsystems) in a short axis plane (thickness: 300 μm). The slices were kept in cold Ca2+-free Tyrode's solution during the whole procedure and continuously supplied with oxygen. To prevent Ca2+ overload, the slices were incubated for 30 min under low Ca2+ conditions (0.9 mM) before transfer into cold Iscove's medium and subsequent rewarming in an incubator (37°C; 5% CO2). After at least 1 h of recovery, the slices were placed in a custom-made hypoxia chamber to simulate myocardial infarction. The hypoxia chamber was filled with Tyrode's solution that contained 1.5 mM Ca2+ and an equimolar concentration of 2-deoxy-glucose instead of D-glucose to prevent glucose metabolism. Oxygen deprivation was obtained by bubbling with pure nitrogen. Complete loss of contractility and vitality was achieved after 20 h.
Co-culture
Beating hESC-derived clusters were mechanically dissected. A single beating cluster was placed partially beneath an avital ventricular slice, which served as matrix, in order to achieve direct contact. After a few days, hESC-derived clusters partially integrated into the matrix, resulting in the movement of the avital tissue slice. Every second to third day, Iscove's medium was changed. After 4–14 days of co-culture, the measurements were performed (Fig. 1A).
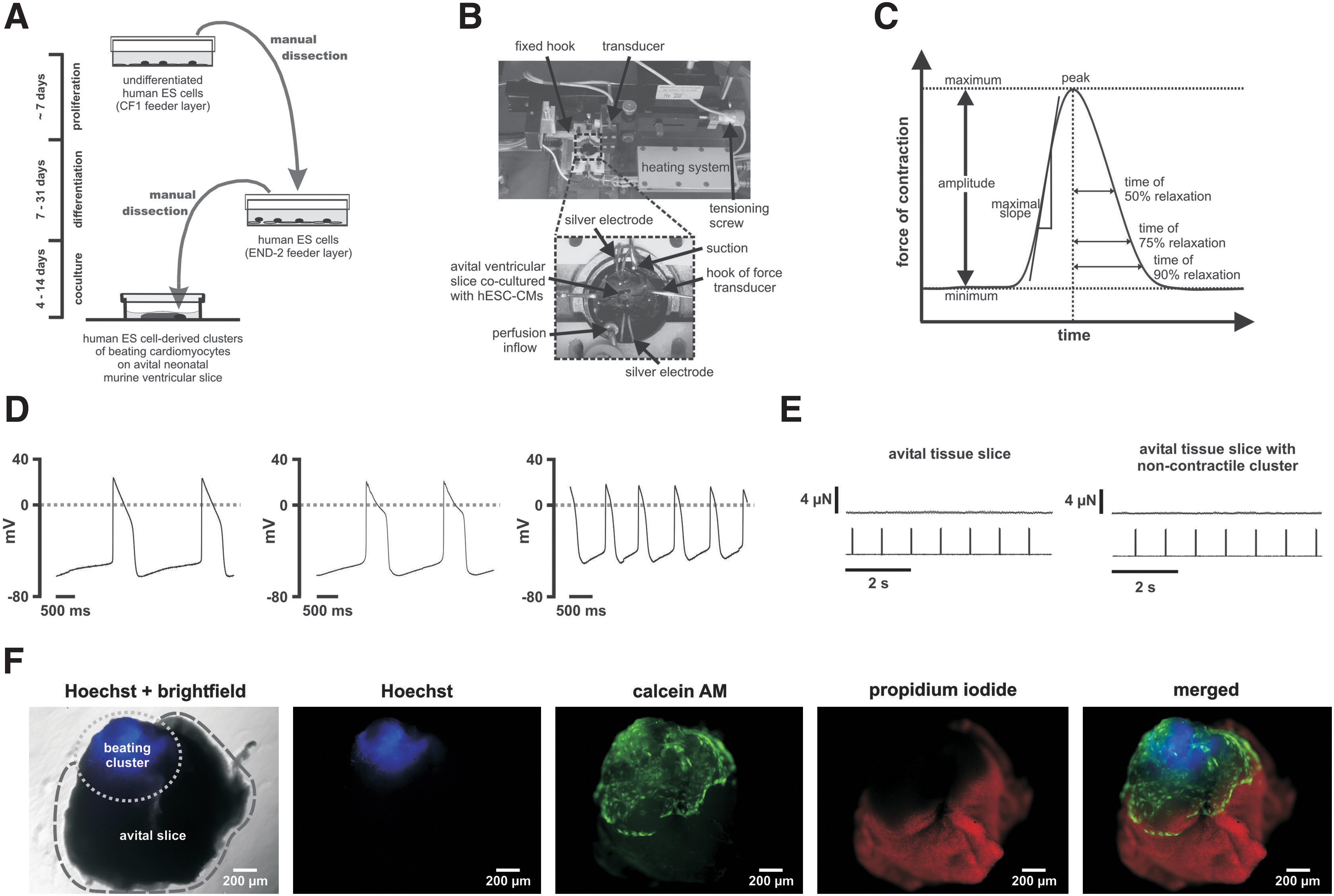
Co-culture.
Force measurements
Measurements of contractile force were obtained by mounting avital ventricular slices co-cultured with a single hESC-derived beating cluster onto the 2 custom-made J-shaped hooks (steel needles; outer diameter 0.22 mm; Hwato Suzhou Medical Appliance Factory). One hook was fixed to the setup assembly, while the second one, which was connected to an isometric force transducer (KG7A; range: 0–5 mN, resolution 0.2 μN, resonance frequency 250–300 Hz; Scientific Instruments), was movable via a micrometer screw to control the length of preparations. The signal was conducted to a bridge amplifier (BAM7C Scientific Instruments) followed by digital conversion via an A/D board (National Instruments;
Data and statistical analysis/force of contraction and beating frequency
The software Dasylab® (version 7.0; National Instruments) was used for signal recording, processing, and analysis. Amplitudes and frequencies were determined by averaging amplitudes and frequencies of one hundred single intervals of 3 s duration; (see Fig. 1C and Supplementary Fig. S1 for details; Supplementary Data are available online at
The following parameters were determined for twitch analysis: amplitude; maximum slopes; time from peak to 50% relaxation, 75% relaxation and 90% relaxation (Fig. 1C). For normalization, we used values at baseline condition for each of the individual experiments; for [Ca2+]ec experiments, values at [Ca2+]ec=2.5 mM served as reference. Twitch overlays were received by averaging single twitches for 5 min after stable conditions had been achieved. Amplitudes and frequencies reported in conjunction with the analysis of relaxation (i.e., Table 1) are based on the algorithm that is optimized for twitch analysis by automatically detecting and superimposing individual twitches (see Supplementary Fig. S1). Therefore, data in Table 1 are very similar but not identical to the amplitudes and frequencies that have been analyzed as just described.
Effect of [Ca2+]ec, NIF and ISO on relaxation: Parameters are normalized to values at 2.5 mM for [Ca2+]ec experiments and to baseline conditions for NIF and ISO experiments. Values are means±SEM.
n=9 (one preparation did not contract at 0.5 mM).
Indicates a significant difference as compared with values at 2.5 mM [Ca2+]ec (P<0.05).
Indicates a significant difference to baseline (P<0.05).
[Ca2+]ec, extracellular calcium; NIF, nifedipine; ISO, (-)-isoproterenol.
All data are expressed as means±SEM for the number of preparation indicated (n). Significance was determined using a paired or unpaired t-test where appropriate. P values<0.05 were considered statistically significant. All statistical analysis was performed in PASW 18.0 (SPSS Inc.).
Electrophysiological characteristics of hESC-CMs
Action potential (AP) recordings were performed on day 4 of co-culture with irreversibly injured ventricular tissue slices. Intracellular recordings of APs were performed as previously described [18] with sharp electrodes (30–50 MΩ, when filled with 3 M KCl) made of borosilicate glass capillaries (WPI; Sarasota). Briefly, for signal amplification, an SEC-10LX amplifier (npi electronic; Tamm) was used. Data were acquired with the Pulse software package (HEKA) and analyzed offline with the Mini Analysis program (Synaptosoft). All measurements were performed at 37°C in DMEM (concentrations of inorganic salts in mM: CaCl2 1.8, MgSO4 0.8, KCl 5.3, NaHCO3 44, NaCl 110, and NaH2PO4 0.9) without serum.
The majority of cells were embryonic ventricular-like (n=18; 64%), the other cells were embryonic atrial-like (n=10; 36%). APs (n=28) were characterized by a maximum diastolic potential of −49±2 mV, an amplitude of 59±3 mV, an AP duration at 50% of 258±21 ms, an AP duration at 90% of 359±23 ms, a 10%–90% rise time of 10.9±0.7 ms, and a maximum rate of the AP upstroke (dV/dtmax) of 10.1±1.1 V/s (Fig. 1D).
Control experiments
OGD-treated murine ventricular tissue slices alone and OGD-treated slices co-cultured with noncontractile H1 derived clusters developed no detectable force of contraction (Fig. 1E).
Live/dead-cell viability staining
A Live/Dead-Cell viability staining was performed (see Supplementary Materials and Methods) to demonstrate the viability of the H1-derived beating clusters and the severe damage of the ventricular tissue slice (Fig. 1F).
Chemicals were of high purity and were purchased from Sigma-Aldrich GmbH unless otherwise stated.
Results
Force-frequency relationship
Experiments that determine the force-frequency relationship were limited by the spontaneous beating and the restricted capture of the preparations during stimulation at higher frequencies. Given these constraints, we had no indication of a negative or positive force-frequency relationship and could not detect any significant potentiation when the stimulation frequency was increased or decreased (Fig. 2).
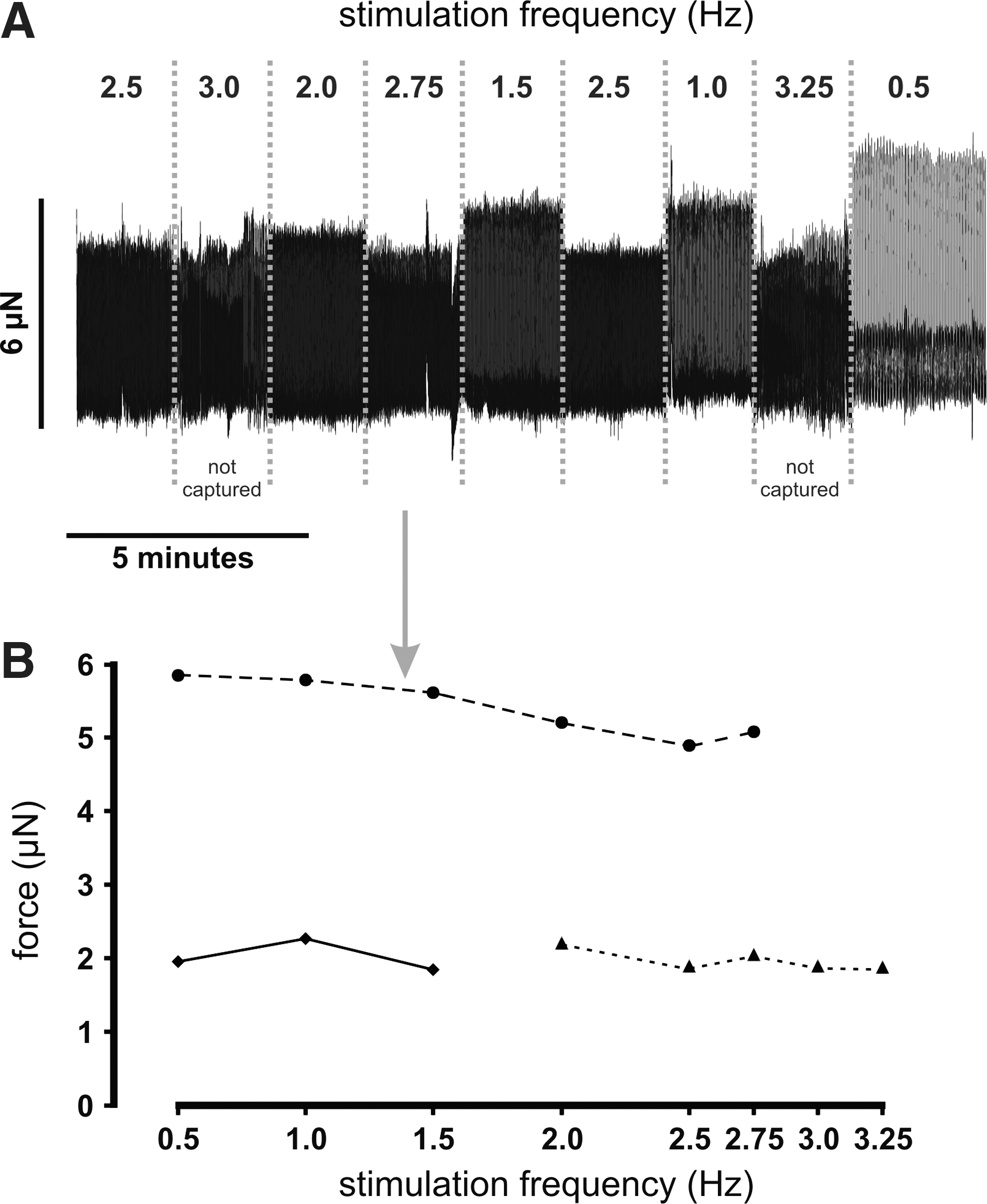
Force-frequency relationship.
Effects of [Ca2+]ec
When exposed to a nominal Ca2+-free Tyrode's solution, no active force generation could be detected in any preparation. Force of contraction increased with increasing [Ca2+]ec in spontaneously beating preparations (0.5 mM: 1.0±0.3 μN, 2.5 mM: 2.0±0.4 μN, 4.5 mM: 2.3±0.5 μN, 6.5 mM: 2.6±0.6 μN, 8.5 mM: 2.8±0.6 μN, n=10) (Fig. 3A, C) and in those stimulated with 1.0±0.1 Hz (0.5 mM: 1.7±1.0 μN, 2.5 mM: 2.7±1.2 μN, 4.5 mM: 3.1±1.2 μN, 6.5 mM: 3.2±1.2 μN, 8.5 mM: 3.2±1.2 μN, n=5) (Fig. 3B, D). In 2 preparations, we tested even a higher concentration of [Ca2+]ec (up to 22.5 mM) and could demonstrate a further increase of the developed force to 149% and 161% of the force measured at [Ca2+]ec of 8.5 mM. The spontaneous beating rate was 0.8±0.1 Hz at [Ca2+]ec=0.5 mM, increased to a maximum of 1.4±0.2 Hz at [Ca2+]ec=2.5 mM, and decreased when [Ca2+]ec was increased further (4.5 mM: 1.3±0.2 Hz, 6.5 mM: 1.2±0.2 Hz, 8.5 mM: 1.0±0.2 Hz, n=10) (Fig. 3C).
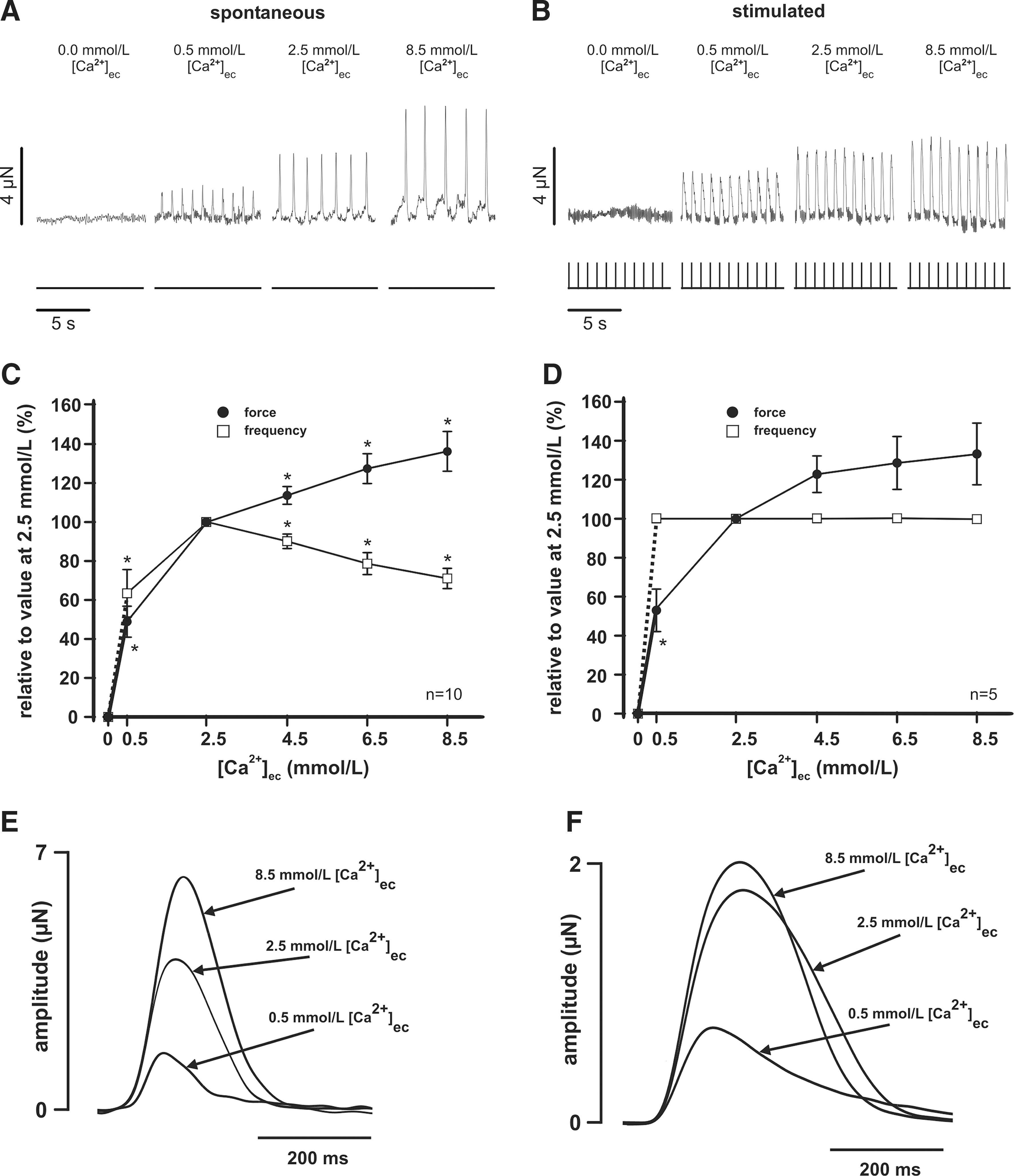
Extracellular calcium ([Ca2+]ec).
Twitch morphology was altered by [Ca2+]ec during spontaneous beating (Fig. 3E) as well as during stimulated contractions (Fig. 3F). Increasing [Ca2+]ec raised the maximal slope of the increase during spontaneous contractions (0.5 mM: 23±3 μN/s, 2.5 mM: 37±7 μN/s, 8.5 mM: 50±10 μN/s; P<0.05) and during electrical stimulation (0.5 mM: 29±12 μN/s, 2.5 mM: 47±19 μN/s, 8.5 mM: 60±23 μN/s) (see Table 1).
In summary, an increase of [Ca2+]ec exhibited a distinct positive inotropic response, and [Ca2+]ec modulated the frequency of spontaneous beating with a maximum at a near-physiological [Ca2+]ec. In contrast, the effect of [Ca2+]ec on relaxation remained equivocal.
Role of Ca2+ channels
To assess the importance of the L-type Ca2+ channel, we administered NIF. After 20 min of baseline recording, NIF was added with increasing concentrations (0.1, 1.0, and 10 μM, if applicable for 20 min each) until contractions were no longer detectable. All experiments were performed in Tyrode's solution. Three out of 5 preparations stopped at a concentration of 1 μM NIF, spontaneous beating abrogated in the remaining experiments at 10 μM NIF. During baseline condition, spontaneous beating was observed at a rate of 1.3±0.3 Hz, the mean developed contraction force amounted to 5.0±1.8 μN. After application of 0.1 μM NIF, frequency increased slightly to 1.3±0.3 Hz (103%±6%), force of contraction decreased to 4.9±1.8 μN (96%±2%), and after application of 1 μM NIF, mean frequency was 1.2±0.3 Hz (97%±9%), and force was 3.9±1.6 μN (69%±10%) (n=5).
Reproducibly, the decrease in the force of contraction was continuous, while the beating rate remained almost stable until beating stopped, frequently advanced by a transitory increase of the beating rate during the last 1–2 min before cessation (Fig. 4A). This observation is further supported by an analysis of the last 10 min before cessation of spontaneous activity (Fig. 4B). These results stress the divergent effects of NIF on the beating rate in contrast to its influence on the development of force. The largest discrepancy between the diminishing force of contraction and the frequency could be observed 1–2 min before cessation of contractile activity when contraction force was halved while frequency was still completely unchanged (Fig. 4B).

Ca2+ channel blocker.
A very similar behavior was observed in 3 preparations that were treated with NIF during electrical field stimulation at a constant rate of 1.5±0.4 Hz (Fig. 4C). Application of NIF resulted in a decrease of the developed force after application of 1 μM NIF (1.8±0.4 μN: 66%±18%) compared with baseline conditions (force: 2.8±1.2 μN). However, the developed force increased after 0.1 μM NIF in one preparation before 1.0 μM NIF abrogated force development. Despite field stimulation, preparations showed a decrease in frequency during the last 2–3 min before stimulated beating ceased in contrast to the behavior or spontaneously beating preparations (1.5±0.5 Hz: 88%±12%, Fig. 4C).
Twitch morphology was altered by NIF during spontaneous beating (Fig. 4D) as well as during stimulated contractions (Fig. 4E). NIF did not significantly reduce the maximal slope of the increase during spontaneous contractions (baseline: 82±32 μN/s, 0.1 μM: 80±31 μN/s and 1.0 μM: 66±29 μN/s) and during electrical field stimulation (baseline: 38±21 μN/s, 0.1 μM: 37±17 μN/s and 1.0 μM: 22±6 μN/s). However, the maximal slope of the decrease was not significantly steeper in spontaneously beating preparations (baseline: 28±11 μN/s, 0.1 μM: 26±10 μN/s and 1.0 μM: 26±10 μN/s) and in stimulated beating preparations (baseline: 20±13 μN/s, 0.1 μM: 20±12 μN/s and 1.0 μM: 14±6 μN/s). Twitches became narrower, because relaxation times shortened (see Table 1). To affirm that the effect is due to a blockade of the L-type Ca2+ channel, we performed additional experiments combining 2 calcium antagonists [NIF and verapamil (VRP)], both of which act predominantly on the L-type channel at concentrations where the unspecific effects of these drugs are negligible. Similar to experiments with NIF alone, the combination of VRP and NIF decreased the beating rate (baseline: 2.0±0.5 Hz [100%], 0.1 μM NIF+0.1 μM VRP: 1.8±0.6 Hz [83%±15%], 0.5 μM NIF+0.5 μM VRP: 0.4±0.4 Hz [16%±16%]; n=3, P<0.05) to a less extent than the developed force (baseline: 2.4±0.6 μN [100%], 0.1 μM NIF+0.1 μM VRP: 1.2±0.2 μN [33%±13%], 0.5 μM NIF+0.5 μM VRP: 0.9±0.2 μN [10%±8%]; n=3, P<0.05) (Fig. 4F).
Beta-adrenergic stimulation
To observe the beta-adrenergic effect, we administered the beta-adrenergic agonist (-)-isoproterenol (ISO) (Fig. 5). It was added with increasing concentrations (10−9, 10−8, 10−7, and 10−6 M). All experiments were performed in Tyrode's solution. During baseline conditions, spontaneous beating was observed at a rate of 1.0±0.1 Hz, the mean developed force amounted to 3.7±0.4 μN. ISO induced a significant dose-dependent increase of the frequency (Fig. 5C), with a maximum of 1.6±0.1 Hz (162%±14%) at 10−6 M. In contrast to the effect of the frequency, ISO did not change the developed force significantly (10−9 M: 3.7±0.4 μN, 10−8 M: 3.8±0.4 μN, 10−7 M: 3.8±0.4 μN, 10−6 M: 3.7±0.4 μN, n=11). During electrical stimulation at a constant rate (1.9±0.3 Hz, n=4), we also observed that the developed force was almost unchanged (Fig. 5D). ISO increased the force of contraction only to a maximum of 3.0±0.4 μN (108%±5%) after 10−6 M (baseline: 2.8±0.4 μN).

Beta-adrenergic stimulation.
Twitch morphology was altered by ISO during spontaneous beating (Fig. 5E) as well as during stimulated contractions (Fig. 5F). ISO did not change the maximal slope of the increase during spontaneous contractions after 1 μM ISO (baseline: 60±8 μN/s, 10−6 M: 60±9 μN/s). Furthermore, ISO did not significantly augment the maximal slope of increase after application of the same dose in spontaneously beating preparations nor was there an alteration during electrical field stimulation (baseline: 39±5 μN/s, 10−6 M: 45±5 μN/s). The maximal slope of the decrease was significantly changed neither in spontaneously beating preparations (baseline: 31±4 μN/s, 10−6 M: 28±8 μN/s) nor in stimulated beating preparations (baseline: 22±4 μN/s, 10−6 M: 25±3 μN/s). Twitches became narrower, because relaxation times decreased during spontaneous and stimulated beating (see Table 1).
Discussion
Since proof-of-principle experiments already unambiguously confirmed differentiation into cardiomyocytes, this study aimed at characterizing the principal qualities of hESC-CMs. Clinically relevant problems are caused not only by systolic but also by diastolic dysfunction and dysregulation [19,20]. Therefore, we determined the basic characteristics of cardiac cells: regulation of inotropy, relaxation, and lusitropy by isometric force measurements of early cardiomyocytes.
We could demonstrate the importance of [Ca2+]ec and the L-type Ca2+ channel for the developed force. In contrast to the situation in the adult human heart, we could not find a significant positive inotropic effect of beta-adrenergic stimulation by ISO in early hESC-CMs despite a definite positive chronotropic effect.
Previous studies showed that external Ca2+ was not required for the activity of murine ES cells early in the pathway to cardiac differentiation [21]. In contrast, in our study, contraction of hESC-CMs was clearly dependent on [Ca2+]ec. Though we tested extremely high [Ca2+]ec only in 2 experiments (see Ref. [22] for comparison), the highest [Ca2+]ec was required to achieve the maximal force of contraction. In addition, our data support the predominant importance of the L-type Ca2+ channel for the contraction of early hESC-CMs. Blockade of this channel by NIF or NIF plus VRP resulted in a decline of the developed force even more than reduction of the frequency of spontaneous activity.
These observations support previous reports demonstrating that the embryonic and fetal heart is primarily dependent on trans-sarcolemmal Ca2+ influx, rather than Ca2+ released from the sarcoplasmic reticulum (SR) [23]. Relaxation times did not change consistently during modification of [Ca2+]ec, while NIF shortened relaxation times, which is in line with a shortening of the AP duration in hESC-CMs [6] (Table 1). However, it should be kept in mind that the force decline during relaxation might not only be regulated by a decline in the intracellular calcium concentration ([Ca2+]ic) but also depends on myofilament properties [24].
Astonishingly, there was no positive inotropic response to the nonspecific beta-adrenergic agonist ISO. The typical response to ISO is thought to involve mainly the cAMP-induced phosphorylation of L-type Ca2+ channels (which increases sarcolemmal Ca2+ entry) [25] and of phospholamban (which leads to increased SR Ca2+ uptake and loading) and of ryanodine receptor (RyR) [which increases calcium release of the SR during Ca2+-induced Ca2+ release (CICR)] (see Ref. [26] for review). Thus, though recently, cardiac troponin I (cTnI) phosphorylation has also been considered important in the positive inotropic response to beta-adrenergic stimulation [27], the action of beta-adrenergic agonists largely relies on mechanisms related to the SR. The lack of an inotropic reaction despite a pronounced chronotropic response after beta-adrenergic stimulation is most likely explained by a still rudimentary function of the SR in the early hESC-CM. Previous studies already described almost absent effects of modulators of SR function in hESC-CMs [28]. These findings are supported by earlier reports that could not detect a t-tubular system [29 –31] at all. Only recently, reports showed the development of t-tubuli after long-term culture (40 days) of hESC-CMs and even the formation of an M-line [13]. In vivo studies have shown that T-tubules frequently develop even after birth [32,33]. Thus, our data confirm and significantly extend these observations, stressing that the development of a functional SR with a substantial CICR occurs quite late during cardiac development.
In contrast, despite a missing positive inotropic effect, ISO elicited a definite positive chronotropic response. This positive chronotropy has been previously described and is primarily mediated via a beta1 adrenoceptor-mediated activation of the cAMP-dependent protein kinase A (PKA) [34]. One possible target for PKA that might explain the increased rate of spontaneous activity is the L-type Ca2+ channel, where PKA increases the opening probability [34].
Local subsarcolemmal ryanodine receptor-mediated Ca2+ releases via RyRs that produces an inward current via activation of Na+/Ca2+ exchange has also been identified as a further mechanism during diastolic depolarization in rabbit sinoatrial nodal pacemaker cells [35].
ISO shortened relaxation times in spontaneous beating preparations. This is most likely due to the β-adrenergic stimulation of PKA-mediated phosphorylation of cTnI and phospholamban via cAMP. Even if the relative contribution of these 2 phosphoproteins is still a matter of debate [19,20,36], these 2 factors are currently thought to be the most important regulators of relaxation. A phosphorylation of the thin filament component cTnI down-regulates its sensitivity to calcium, resulting in a faster inactivation [37,38]. Phospholamban phosphorylation increases the activity of the sarco-endoplasmic reticulum calcium ATPase. As a consequence, calcium is shifted faster from the cytosol into the SR [37].
In conclusion, contractility of early hESC-CMs is already largely regulated by [Ca2+]ec and L-type Ca2+ channels. However, though beta-adrenergic stimulation induced a pronounced increase of the spontaneous beating frequency, a significant positive inotropic effect was still missing. It is most likely explained by the immaturity of the SR and emphasizes the need to understand the mechanisms involved in the development of the SR.
However, there are some limitations of our study: Though the environment is derived from ventricular tissue and, therefore, might constitute a reasonable matrix, it is avital and, therefore, not physiological. In addition, beating areas still contain an inhomogeneous mixture of cell types [8]. Even if embryonic ventricular-like cells dominate, there remain a substantial number of embryonic atrial-like cells. Culture conditions (e.g., END-2 cells) are known to influence the composition of the beating areas and the degree of maturation substantially [39]; differences between cell lines have also been described [40]. Therefore, one should be cautious when generalizing our results.
Nevertheless, these observations stress the importance of a functional assessment of the cardiac myocytes suggested for replacement strategies. If early hESC-CMs are used for cell replacement therapies, further experiments will have to prove in vivo differentiation or introduce new methods of in vitro maturation, especially regarding the development of a functional SR.
Footnotes
Acknowledgments
The authors thank Christine Mummery and Robert Passier (Hubrecht Laboratory, Utrecht, Netherlands) for providing END-2 cells and support with the culture of hESC-CMs, Cornelia Böttinger for supplying them with hESC-CMs, and Annette Köster and Christoph Backhausen for their skilful technical assistance. They also thank Suzanne Wood for the secretarial assistance and the electronic and mechanic workshop, especially Frank Stassen and Harald Metzner for the technical support.
This work was supported by the Koeln Fortune Program/Faculty of Medicine, University of Cologne (grant: 157/2003) (F.P.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.