
Abstract
Mesenchymal stem cells (MSCs) that are present in many adult tissues can generate new cells either continuously or in response to injury/cancer. An increasing number of studies demonstrated that MSCs have the ability to differentiate into cells of mesodermal origin and transdifferentiate into cells such as hepatocytes, neural cells. There has been growing interest in the application of MSCs to cancer therapy. The relationship between MSCs and cancer cells remains highly controversial. In this study, we analyzed the interaction of bone marrow-derived MSCs and cancer cells by cell–cell contact and transwell culture system. The flow cytometry and real-time polymerase chain reaction showed that after coculture of MSCs and cancer cells, MSCs displayed the hematopoietic cell markers such as CD34, CD45, and CD11b. The CD68, MRCI, and CSF1R were dramatically upregulated after coculture. The cytokine array showed that MSCs after coculture secreted monokines and chemokines much more than that of intact MSCs. The MSCs under tumor conditions were responsive to stimulation with lipopolysaccharide by cytokines release. The tumor-conditioned MSCs showed phagocytic ability and enhanced release of nitric oxide, which are the characteristics of macrophages. Calcium ion is an important intracellular messenger responsible for differentiation and gene expression regulations. The influx of Ca2+ into MSCs was obviously reduced after coculture. The blocking of calcium channel with verapamil obviously increased the expression of CD34, CD45, and CD11b, thus indicating that the diminished calcium ion influx is coupled with the hematopoietic differentiation of MSCs under tumor conditions. Taken together, in a cancer environment, MSCs could effectively differentiate into immune hematopoietic cells, precisely macrophages. Diminished transient influx of Ca2+ may mediate the hematopoietic differentiation of MSCs.
Introduction
M
A growing body of research has focused on the use of MSCs in cancer therapy. It has been demonstrated that MSCs can move to the tumor sites, and MSCs engineered to express antitumor factors could be an innovative choice as a cell-mediated gene therapy to inhibit tumor growth [6,7]. There are concerns that MSCs promote cancer growth or MSCs might transform to the tumorigenic cells, although this issue remains controversial and not well understood [8]. The relationship between the cancer cells and MSCs remains elusive. This complex dialog between MSCs and cancer cells is certainly critical for the outcome of tumor development. The differentiation potential of MSCs in a cancer microenvironment needs to be explored.
Several studies have suggested that phenotypically normal and functionally competent cells of hematopoietic system could be derived from embryonic stem (ES) cells, bone marrow hematopoietic stem cells (HSCs), or peripheral blood HSCs [1]. The hematopoietic differentiation of adipose tissue resident MSCs under differentiation media has also been recently reported [1]. Adipose tissue-derived stem cells were also reported as expressing angiogenic and hematopoietic factors [9]. Bone marrow-derived very small embryonic/epiblast-like stem cells were demonstrated as differentiating into the hematopoietic lineage after coculture over stromal cells [10]. Hematopoietic differentiation capacity of bone marrow-derived MSCs remains unknown.
Calcium ion was reported to regulate osteogenic differentiation and fate management of HSCs [11]. Ca2+ activates several myogenic transcription factors such as myogenin and myocyte enhancer factor-2 [12 –15], thus triggering the differentiation of myocytes. Ca2+ was reported as playing fundamental roles in the development of oligodendrocyte progenitor cells [16]. Here, we ask whether Ca2+ plays roles in the differentiation of MSCs under tumor conditions.
In this study, we found that bone marrow-derived MSCs are able to transdifferentiate into cells of hematopoietic lineage, more precisely macrophages, when cocultured with cancer cells. We further investigated the effect of calcium ion on the hematopoietic differentiation of MSCs. We concluded that the diminished Ca2+ influx was coupled with the hematopoietic differentiation of MSCs.
Materials and Methods
Cell culture
Human MSCs were obtained from bone marrow aspirates of ribs of patients undergoing thoracic surgery. The isolation and culture of MSCs were performed by using previously described methods [6]. Samples were from Second Xiangya Hospital, Central South University. The patients were informed about the sample collection and signed informed consent forms. Collections and use of tissue samples were approved by the ethical review committees of Second Xiangya Hospital. Human nasopharyngeal carcinoma cell line HK1, 5-8F, and 6-10B and human lung adenocarcinoma epithelial cell line A549 were grown in DMEM supplemented with 10% fetal calf serum at 37°C under 5% CO2 and 95% air. Coculture experiments were performed by a direct contact and transwell system (Corning HTS Transwell, 0.4 μm). The osteogenic, chondrogenic, and adipogenic differentiation were performed by using the differentiation media (Cyagen Bioscience, Inc.)
Flow cytometry
Flow cytometry was performed on bone marrow-derived MSCs stained for CD90, CD44, CD105, CD34, CD45, and CD11b. The following antibodies specific for human molecules were used: PC5-CD11b (Beckman Coulter), FITC-CD44 (Beckman Coulter), PC7-CD45 (Beckman Coulter), FITC-CD90 (Beckman Coulter), ECD-CD34 (Beckman Coulter), and PE-CD105 (eBioscience). Flow cytometry was performed on Moflo XDP (Beckman Coulter). The corresponding isotype control mAbs was from Beckman Coulter.
Lentiviral vectors and MSC transfection
The lentiviruses were created by using the ViraPower™ Lentiviral Expression System (Invitrogen). The coding sequence of green fluorescent protein (GFP), red fluorescent protein (RFP) was subcloned into Lenti6/V5-D-TOPO. The detailed processes were previously described [6].
Cytokine assay
The cytokines released from cells with and without stimulation of lipopolysaccharide (LPS) (1 μg/mL) were assessed by using the RayBio Human Cytokine Antibody Array 5 (RayBio Tech., Inc.). Seven days after coculture, the MSCs and HK1 cells, tumor-conditioned medium were removed, and cells were cultured in fresh medium for an additional 3 days. The collected media were applied on cytokine membranes. The chemiluminescent signal was detected by using a CCD imaging system (Alpha Innotek FluoChem). The resulting images were analyzed by using Quantity One Software to measure the expression of various cytokines. An uncultured media aliquot was used as a negative control sample. A positive control was used to normalize the results from the different membranes being compared. By comparing the signal intensities, relative expression levels of cytokines were made.
Phagocytosis assay
The phagocytotic ability of cells was determined by incubating the cells with Escherichia coli (K-12 strain) BioParticles®, Alexa Fluor® 488 conjugate (Invitrogen). Cells were thoroughly rinsed with PBS and imaged under a fluorescence microscope (Canon). Flow cytometry was also used, and the cells were excited with an argon lazer at a wavelength 488 nm and detected at 530 nm by using flow cytometry (Moflo XDP; Beckman Coulter).
Measurement of nitrite levels (Griess test)
NO• generated as a result of MSCs differentiation was determined by measurement of accumulated nitrite in the cell culture medium and cell lysate. One hundred microliters of the medium or cell lysate was taken from each culture well, mixed with 100 μL Griess reagent (50 μL of Griess A and 50 μL of Griess B, Beyotime, Inc.), incubated at room temperature for 10 min, and the absorbance at 540 nm was determined. Fresh tissue culture medium served as the blank, and solutions of NaNO2 served as standards.
Measurement of Ca2+
Measurement of cell Ca2+ was performed by flow cytometry. Briefly, Ca2+ was measured with Fluo-3 AM (Beyotime, Inc.). Fluorescence in cells was excited with an argon lazer at wavelength 488 nm and detected at 530 nm by using flow cytometry (Moflo XDP; Beckman Coulter).
Immunofluorescence
Cell were blocked and permeabilized in 2% bovine serum albumin (BSA)/0.2% Triton X-100 in antibody buffer (150 mM NaCl, 50 mM Tris base, 2% BSA, 100 mM L-Lysine, 0.04% Na azide, and pH 7.4) for 60 min at room temperature. Primary antibodies against human molecules ECD-CD34, PC7-CD45, or PC5-CD11b (Beckman Coulter) were diluted in antibody buffer (1:2,000). Cells were incubated in primary antibody solution overnight at 4°C, rinsed thrice in PBS, and then mounted in mounting medium with DAPI. Cells were imaged under a fluorescence microscope (Canon).
Real-time reverse transcription–polymerase chain reaction
Real-time polymerase chain reaction (PCR) was performed by using SYBR Green Master Mix. The primers were synthesized as showed in Table 1. Real-time PCR was performed on the BIO-RAD IQTM5 Multicolor Real-Time PCR detection System (Bio-Rad). The data were analyzed by using iQ5 software. The relative gene expression was quantified based on the threshold cycle value and normalized to the housekeep gene GAPDH. The data are representative of the means of 3 experiments. Student's t-test was applied when 2 or more values were compared. P<0.05 was considered a significant difference.
Results
Isolation and characterization of MSCs
Bone marrow-derived MSCs were isolated from ribs of patients undergoing thoracic surgery. Immunophenotyping confirmed that the bone marrow-derived MSCs displayed mesenchymal markers such as CD90+, CD44+, CD105+, CD34−, CD45−, and CD11b−, thus indicating the absence of hematopoietic progenitor cells (Fig. 1A). The capability of differentiating into adipogenic, osteogenic, and chondrogenic lineages in vitro was displayed in Fig. 1B. The capacity of differentiation showed the enhanced alkaline phosphatase activity, osteocalcin production, proteoglycan production, and mineral deposition.

Phenotypic analysis of bone marrow-derived mesenchymal stem cells (MSCs).
MSCs cocultured with cancer cells displayed hematopoietic differentiation
Along with the emerging cancer gene therapy approaches of using mesenchymal stem cells, there are concerns that MSCs might promote tumor growth and metastasis or transform into tumorigenic cells. Elucidating the interaction between MSCs and cancer cells should be an important research focus. Here, we cocultured MSCs with epithelial cancer cell lines HK1, 5-8F, 6-10B, and A549. For the direct contact coculture, cancer cells and MSCs cells were respectively infected with lentivirus-expressing GFP and RFP. The morphology of MSCs dramatically changed after coculture from spindle- to round-shaped cells and then formed multinucleated giant cells or long pseudopodia shape (Fig. 2A). The morphology changes were observed in both direct contact culture and the transwell culture system. The flow cytometry analysis for the transwell cocultured MSCs showed an obvious increase in the size compared with the control MSCs (Fig. 2B). Control culture showed no morphologic alteration.
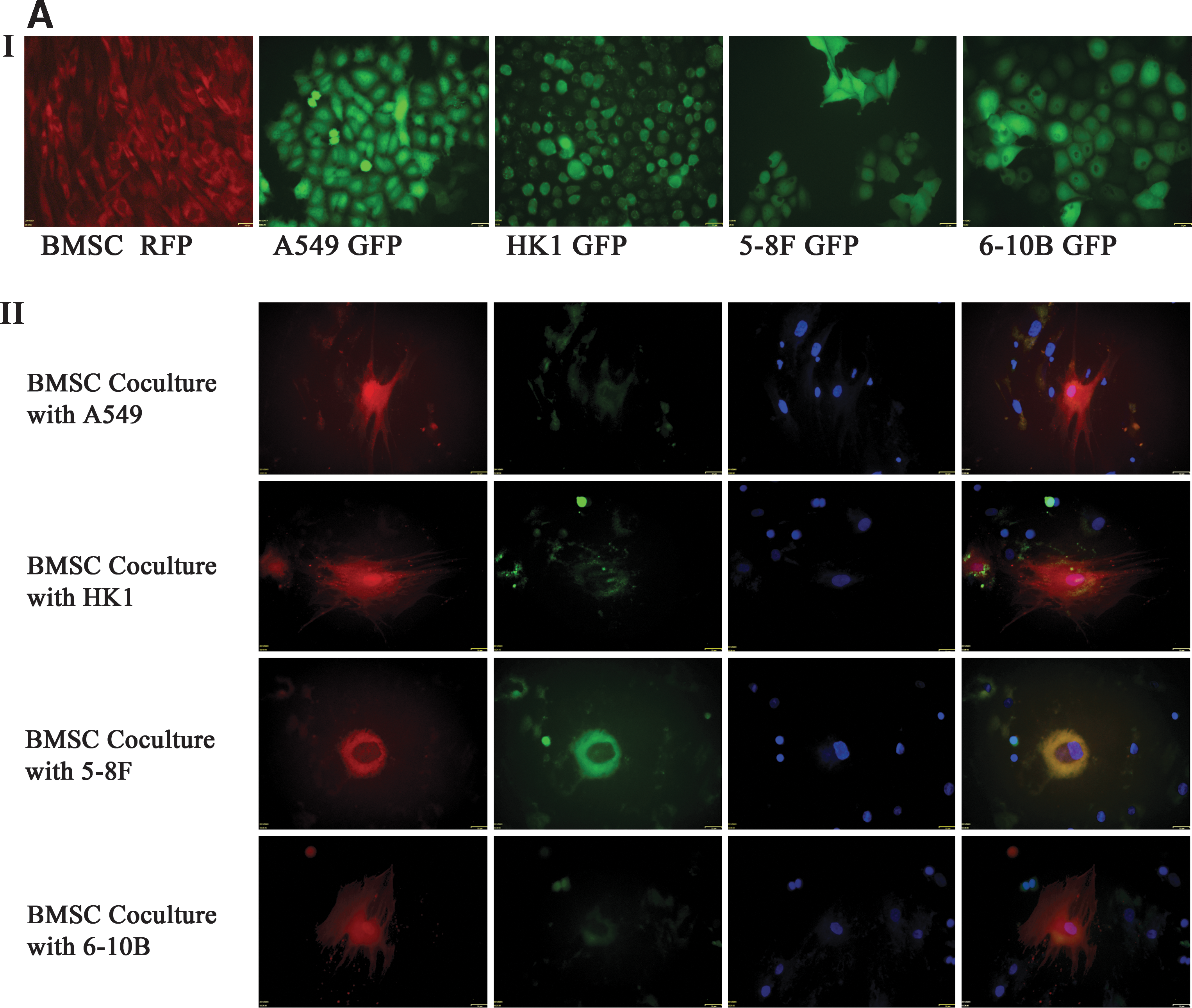
MSCs after coculture with cancer cells displayed hematopoietic differentiation.
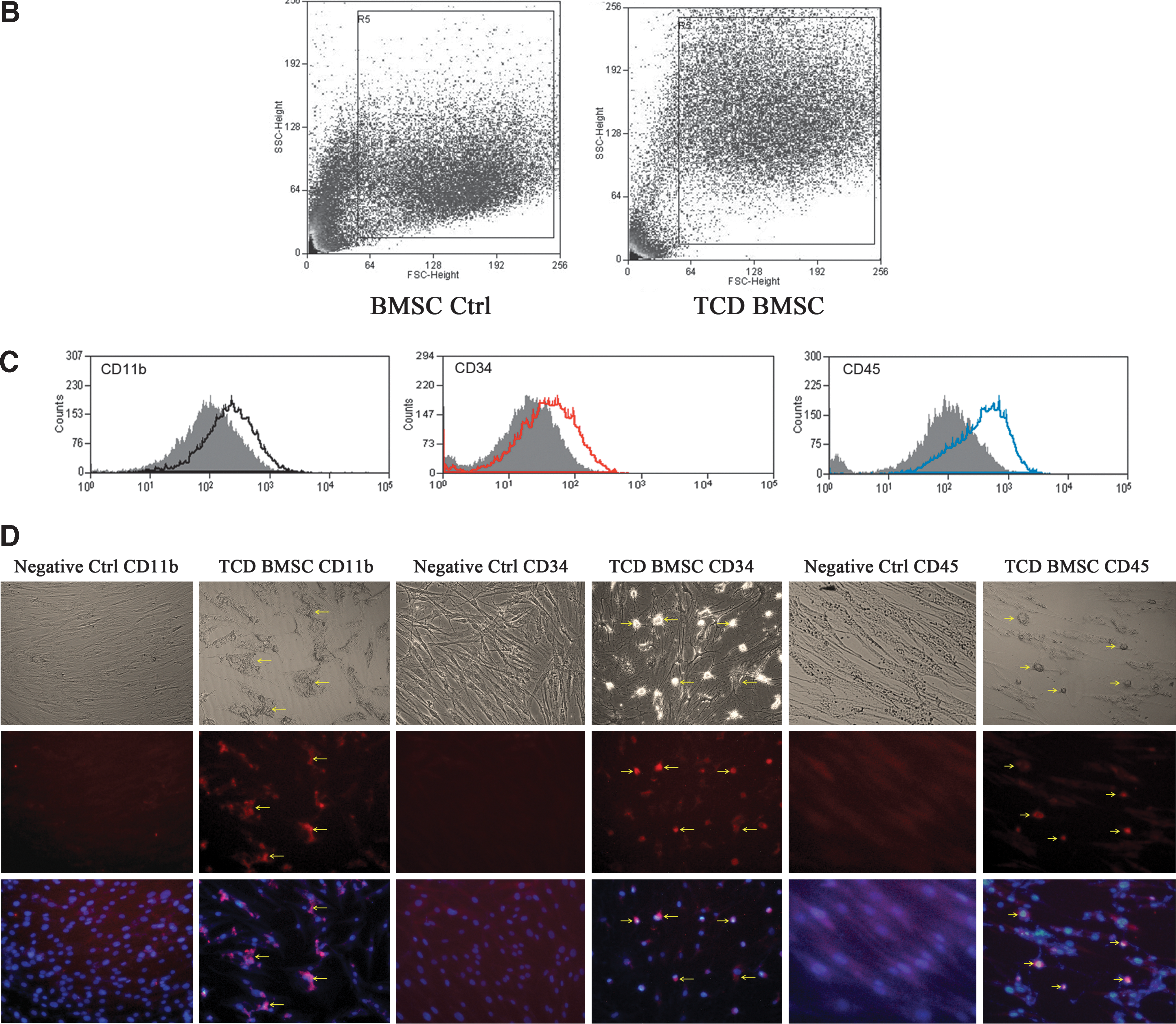
Next, we analyzed the hematopoietic markers on the surface of MSCs by flow cytometry. The MSCs after coculture with cancer cells displayed hematopoietic markers such as CD11b, CD34, and CD45 (Fig. 2C). The MSCs were stained for the expression of hematopoietic markers. Immunofluorescence indicated the expression of hematopoietic lineage specific antigens CD11b, CD34, and CD45 (Fig. 2D).
Since the morphology of MSCs after coculture changed from spindle to multinucleated giant cells, we asked whether these differentiated cells were precisely macrophages. To answer the question, we performed real-time reverse transcription–PCR to investigate the expression of macrophage marker molecules CD68, MRC1, and CSF1R. Gene expression analysis revealed that MSCs after coculture with cancer cells expressed mature monocyte/macrophage markers CD68, CSF1R, and MRC1 (Fig. 3A). Meanwhile, the transcription factor signature of pluripotent stem cells OCT3/4, SOX2, and Nanog were dramatically downregulated (Fig. 3A). These results indicated that in the cancer environment, bone marrow-derived MSCs showed hematopoietic differentiation.
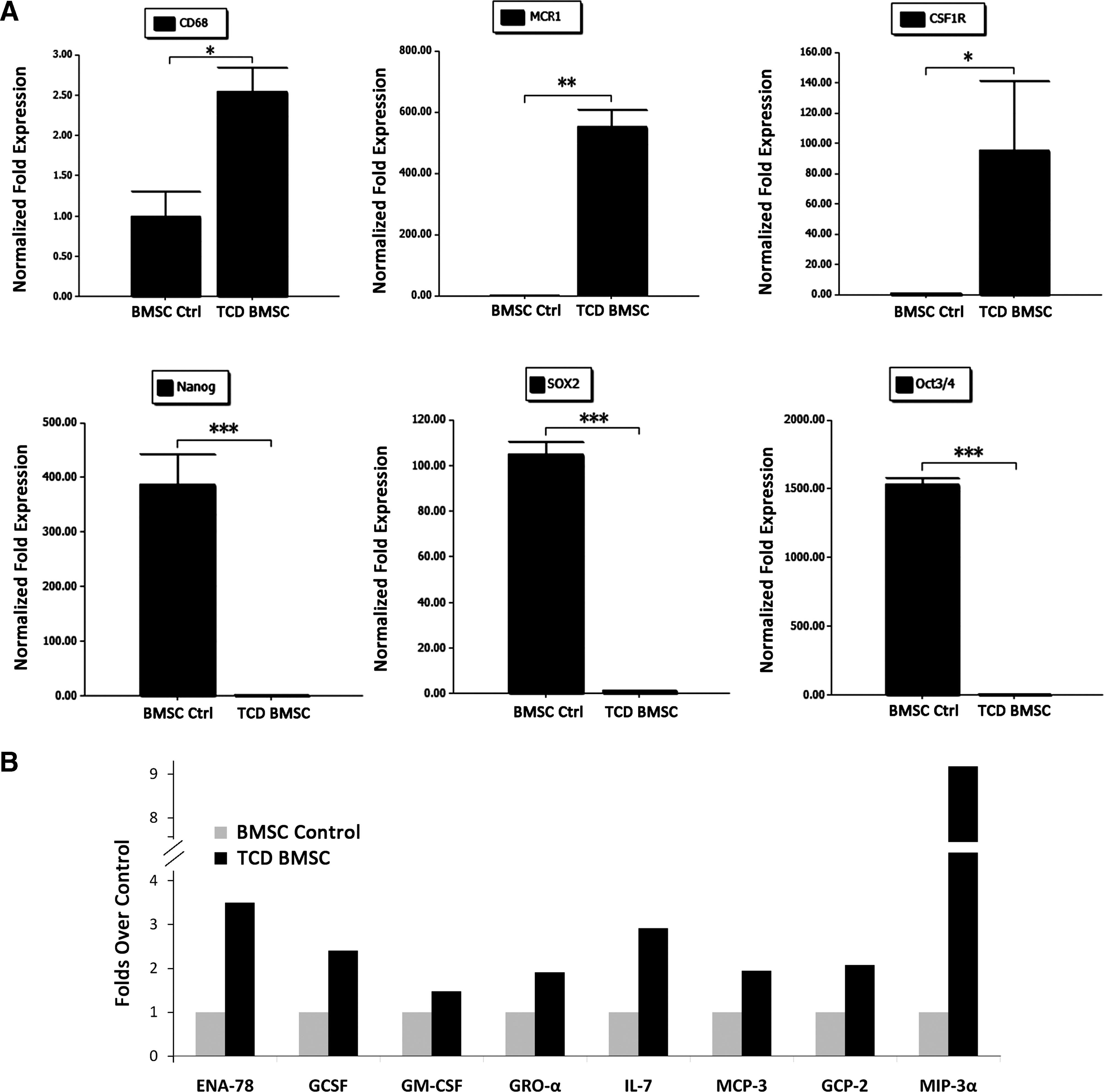
Tumor-conditioned differentiated MSCs cells were precisely macrophages.
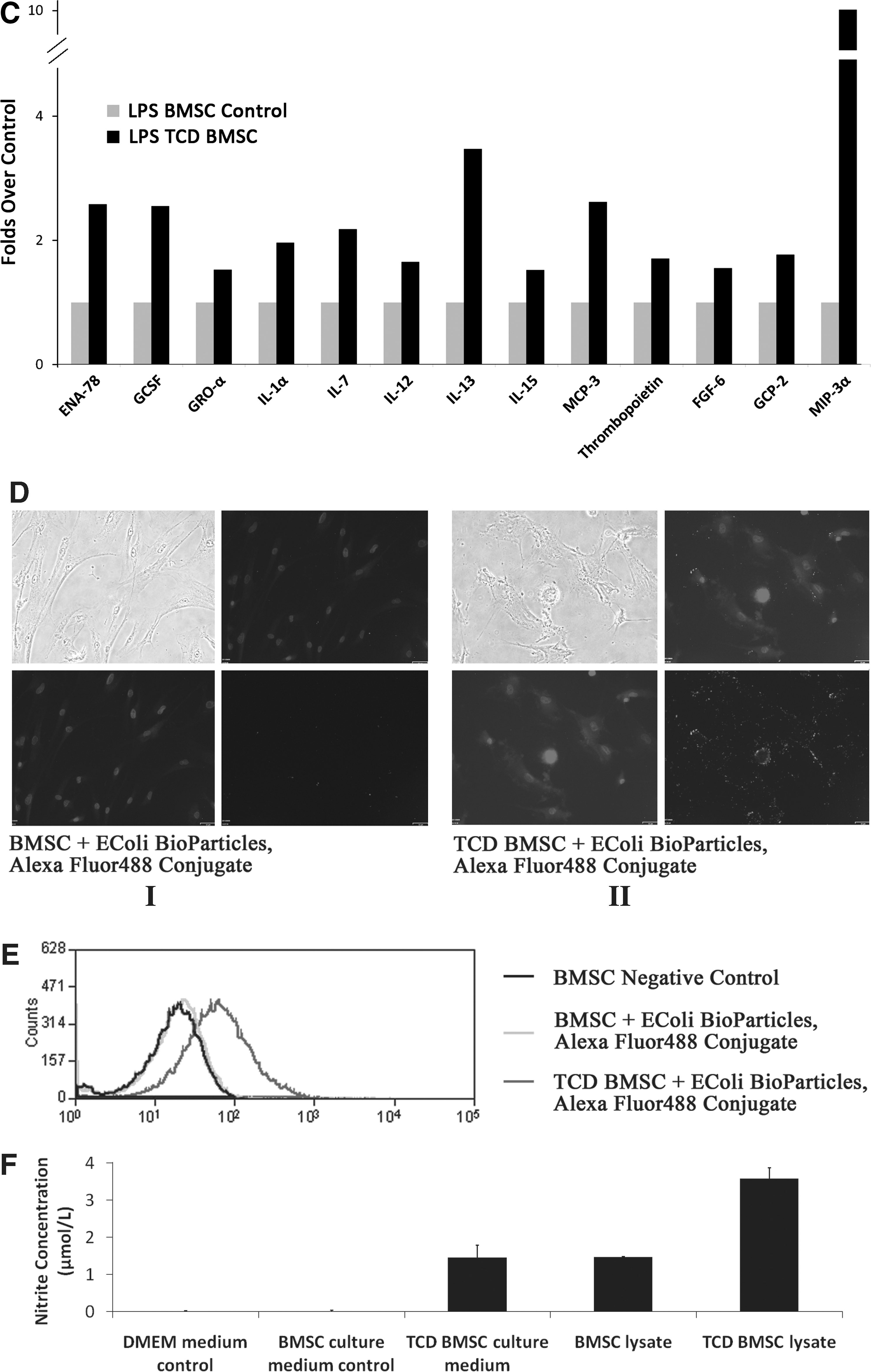
As secretory cells, monocytes and macrophages are vital to the regulation of immune responses and the development of inflammation; they produce a wide array of monokines and regulatory factors such as interleukin-1 (IL-1). In this study, the MSCs after coculture with cancer cells were also assessed for the secretion of monokines and regulatory factors. The supernatants were collected from the tumor-conditioned differentiated MSCs. A considerable upregulation of monokines secreted by mature monocyte/macrophages such as ENA-78 (CXCL5, 3.49-folds), GCSF (2.4-folds), GM-CSF (1.48-folds), GRO-a (CXCL1, 1.9-folds), IL-7 (2.9-folds), MCP-3 (CCL7, 1.9-folds), GCP-2 (CXCL6, 2.07-folds), and MIP-3a (CCL20, 9-folds) were observed in the tumor-conditioned differentiated MSCs compared with undifferentiated MSCs (Fig. 3B). Macrophages play a key role in the inflammatory response and release cytokines in response to stimulator LPS. We further investigate the secretion of cytokines of MSCs on stimulation with LPS. Compared with undifferentiated MSCs, tumor-conditioned differentiated MSCs after stimulation of LPS (1 μg/mL) resulted in a dramatic upregulation of ENA-78 (2.58-folds), GCSF (2.55-folds), GRO-a (1.53-folds), I-309 (1.54-folds), IL-1α (1.96-folds), IL-7 (2.18-folds), IL-12 (1.66-folds), IL-13 (3.5-folds), IL-15 (1.52-folds), MCP-3 (2.62-folds), thrombopoietin (1.71-folds), FGF-6 (1.55-folds), GCP-2 (1.77-folds), and MIP-3a (10-folds) (Fig. 3C).
In addition to the production of cytokines or chemokines, an important macrophage function in response to infection is phagocytosis of bacteria or other particles, and killing of bacterial or other particles [17]. The phagocytic capability of tumor-conditioned differentiated MSCs was evaluated by incubating cells with Alexa Fluor® 488 conjugate E. coli (K-12 strain) BioParticles. The fluorescent microscopy showed the E. coli particles phagocytosed by differentiated MSCs. The control MSCs were unable to phagocytose the E. coli particles (Fig. 3D). The phagocytosis of differentiated MSCs was confirmed by flow cytometry. As shown in Fig. 3E, tumor-conditioned MSCs showed obvious Alexa Fluor® 488 positive.
The effector functions of macrophages include the ability to produce cytotoxic compounds such as nitric oxide (NO). The production of NO by tumor-conditioned MSCs was determined by Griess test. The medium of tumor-conditioned differentiated MSCs and the cell lysate were assayed for nitrite levels. Nitrite concentrations in the medium from control MSCs were undetectable. The nitrite levels in the tumor-conditioned differentiated MSCs were shown in Fig. 3F. The differentiated MSCs medium showed an obvious increase of nitrite concentration. The nitrite concentration in the control MSCs lysate was 1.46 μm. The differentiated MSCs lysate showed more than 2 times increase. Tumor-conditioned MSCs produced a large amount of NO, thus indicating that the tumor-conditioned MSCs showed the macrophages characterization.
Calcium flux coupled with MSCs differentiation
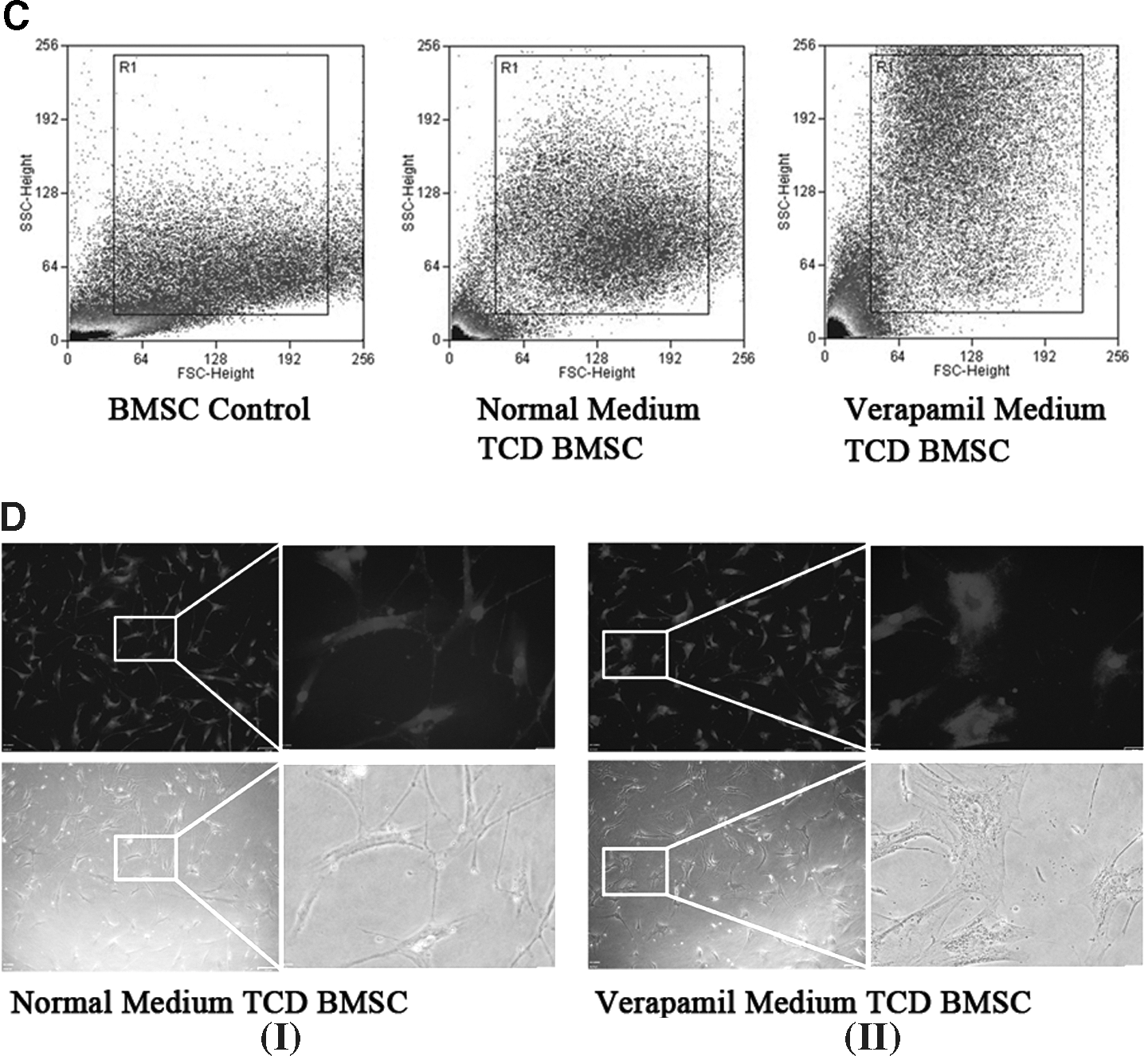
Calcium functions as a ubiquitous intracellular messenger. To examine the role of calcium ion in the tumor-conditioned MSCs differentiation, we performed flow cytometry to explore the intracellular calcium. Primary cultured MSCs were loaded with Ca2+ indicator dye Fura-2. A high concentration of calcium ion was found in the MSCs. The coculture caused a reduction of calcium ion influx to MSCs, which significantly alters the intracellular calcium concentration (Fig. 4A). We further treated cells with verapamil, which is a calcium ion influx inhibitor. We found that verapamil caused the MSCs to display more hematopoietic molecule positive cells and changed phenotype (Fig. 4B, D). The flow cytometry showed that more cells were distributed at the higher region along the y-axis, thus indicating the obvious increase in the cell size (Fig. 4C). As shown in Fig. 4B, after coculture with cancer cells, the percentage of CD34, CD45, and CD11b positive cells was more in the verapamil treatment group than in that of the control. This indicated that diminished calcium ion influx was coupled with the hematopoietic differentiation of MSCs. The exact roles of calcium ion in the differentiation need to be further explored.

Calcium flux coupled with MSCs differentiation.
Discussion
Myeloid and lymphoid lineages are often derived from HSCs. The hematopoietic potential of ES cell and iPS were also reported [1,18,19]. MSCs confirmed the mesenchymal lineage differentiation. In this study, we showed that adherent MSCs cocultured with cancer cells resulted in a generation of cells that exhibit morphologic and functionally distinct hematopoietic characteristics. The morphology of MSCs dramatically changed from spindle- to round-shaped cells and then formed multinucleated giant cells or long pseudopodia shape. Intact MSCs expressed positive mesenchymal markers and none of those associated with the hematopoietic phenotype (CD44+, CD90+, CD105+, CD45−, CD34−, and CD11b−). The negative hematopoietic specific surface marker verified no contamination of HSCs. The MSCs cocultured with cancer cells displayed hematopoietic specific surface marker CD34, CD45, and CD11b. The immunofluorescence confirmed that CD34, CD45, and CD11b were expressed in the round cells. The monocyte/macrophage markers CD68, CD11b, MRC1, and CSF1R were obviously upregulated in MSCs after coculture with cancer cells. The morphology change and the positive macrophage markers indicated that MSCs differentiated precisely into macrophages. The cytokine array assay showed that MSCs after coculture with cancer cells secreted monokines such as ENA-78 (CXCL5), GCSF, GM-CSF, GRO-a (CXCL1),IL-7, MCP-3 (CCL7), GCP-2 (CXCL6), and MIP-3a (CCL20), thus indicating that the tumor-conditioned MSCs displayed hematopoietic differentiation and may also participate in generation of hematopoietic cells [20,21].
The tumor-conditioned MSCs may definitely influence the fate of tumor cells located nearby. MSCs as stromal cells were reported as secreting cytokines to promote cancer cells growth and angiogenesis [22,23]. The immunosuppressive properties of MSCs were also described. MSCs not only escape immune responses but also suppress inflammation [24]. The differentiated MSCs displayed different immunological modulation properties [25,26]. The presence of mature immune cells have been described, including macrophages and lymphocytes that have been largely involved in the inflammatory process linked to cancer. The cytokines secreted on stimulation of LPS showed the macrophages immune responsiveness of tumor-conditioned MSCs. It was shown in the study that the undifferentiated MSCs response on LPS stimulation was much lower than that of the tumor-conditioned MSCs. The phagocytotic ability and production of NO also confirmed that tumor-conditioned MSCs are characteristics of macrophages. In a cancer environment, MSCs can function as effector cells during innate and adaptive immune responses and contribute to elevation of proinflammatory cytokines levels. MSCs were conversely reported to induce T-cell immune response as antigen presenting cells [27]. The immunological roles of MSCs in tumors need to be further investigated.
Several earlier studies have reported that Ca2+ responses appear to diminish with maturation of oligodendrocytes from progenitors to mature cells in culture [16,28]. In this study, we examined Ca2+ influx in MSCs. After flow cytometry analysis, Ca2+ influx was diminished when cocultured with cancer cells. The Ca2+ obviously entered selectively into the cancer cells (data not shown). Blocking Ca2+ influx by verapamil clearly promotes the hematopoietic differentiation of MSCs. The following events after the reduced Ca2+ influx need to be further explored.
Summary
The current work demonstrated that interaction of cancer cells and MSCs promotes the hematopoietic differentiation of MSCs. The diminished Ca2+ influx may play a fundamental role in the differentiation.
Footnotes
Acknowledgments
This work was supported by National Natural Science Foundation, China (Project No. 30971497), Innovation Subject Foundation for Doctoral Student, Hunan Province, China (Project No. CX2010B039), and Hunan Natural Science Foundation (10JJ7003).
Author Disclosure Statement
There are no potential conflicts of interest to be declared.