
Abstract
Recently, an additional type of pluripotent stem cell-line derived from mouse embryos has been established and termed epiblast stem cell (EpiSC), and is expected to be an important tool for studying the mechanisms of maintenance of pluripotency since they depend on basic fibroblast growth factor-MAPK and Activin A-Smad2/3 signaling to maintain pluripotency, unlike mouse embryonic stem cells (ESCs). Further, because of the similarities between mouse EpiSCs and human ESCs, EpiSCs are expected to be effective experimental models for human stem cell therapy. Recently, study for conversion from ESC state to EpiSC state or reversion from EpiSC state to ESC state has attracted interest since these techniques may lead to increasing the potential of pluripotent stem cells and our knowledge about their developmental status. In the present study, we find that a low oxygen concentration in culture environment accelerated, improved, and stabilized the EpiSC state of the converted cells from the ESC state using Oct4ΔPE-GFP transgenic ESCs. Induced EpiSCs (iEpiSCs) in hypoxia possess closer gene expression patterns to native EpiSCs, and bisulfite sequences for the promoter regions of Stella and Oct4 genes have elucidated that the iEpiSC gain EpiSC-specific methylation patterns in hypoxia. Our data provide evidence that oxygen concentration is an important factor for establishment of the EpiSC-specific state.
Introduction
M
Now a different type of pluripotent cells derived from mouse embryos has already been established, termed epiblast stem cells (EpiSCs) [10,11]. EpiSCs are pluripotent stem cells derived from epiblasts of postimplantation embryos; they depend on basic fibroblast growth factor (bFGF)-MAPK and Activin A-Smad2/3 signaling to maintain pluripotency and to perform self-renewal. Although EpiSCs can self-renew over a long-term and appear to be pluripotent by forming teratomas containing 3 embryonic germ layers in vivo, they rarely incorporate into the ICM and contribute to chimeric mice. The differentiation capability of EpiSCs indicates that EpiSCs are less pluripotent than mESCs and may reflect a developmental stage gap with mESCs and recipient embryos: the EpiSCs are derived from embryos at embryonic day 5.75 (E5.75) to E6.5, whereas the mESCs and recipient blastocysts are from E3.5 embryos. The most important part of the study on EpiSCs is the resemblance between mouse EpiSCs and conventional human ESCs (hESCs). Interestingly, although they are derived using blastocysts, conventional hESCs seem to correspond very closely to mouse EpiSCs with respect to many characteristics, including some gene expression, flat colony morphology, and signaling responses in self-renewal and differentiation. For example, both mouse EpiSCs/hESCs appear to be dependent on bFGF, MAPK, and TGFβ/Activin/Nodal pathway activity for self-renewal and differentiate rapidly when treated with MEK, FGFR, and/or ALK4/5/7 inhibitors [10 –14].
Understanding and elucidating mechanisms/factors enable us to switch these 2 pluripotent states, that is, the shuttle between mESC- and EpiSC state can lead to increasing the value of the hESCs. For example, it may provide a basis for generating more ideal types of mESC-like human pluripotent cells from conventional hESCs. Recently, converting ESCs to EpiSCs has been attempted in some groups. Zhang et al. obtained EpiSCs from mESCs via embryoid body formation, which partly mimic post-transplantation development [15]. Guo et al. demonstrated that the EpiSC state could be induced by culturing mESCs in the EpiSC condition, and EpiSCs derived from mouse epiblasts, called native EpiSCs (nEpiSCs) in this article, can be reverted to the ESC state by Klf4 over-expression [16]. The above studies illustrate that the pluripotent state of ESCs and EpiSCs are not fixed, but are influenced by the developmental and environmental context. However, the factors that affect the state-conversions were not fully elucidated.
Oxygen concentration in the environment is an important factor for definition of cell fate and development in embryos [17], and also in cultured cells and embryos [18,19]. Actually, the environment of the uterus is hypoxic in terms of development from pre- to postimplantation embryos [20]. Some reports have shown that reduced oxygen concentration is beneficial for the in vitro maintenance of hESCs in terms of decreasing the amount of spontaneous differentiation and supporting self-renewal [21,22]. On the other hand, it has also been reported that hypoxic conditions negatively affect mESC self-renewal by inhibiting LIFR expression [23]. These results suggest the possibility that the suitable oxygen concentration is different between the mESC state and mEpiSC state. To determine the possibility that the hypoxic condition may be a factor in promoting mESC-mEpiSC conversion, here we examined the conversion of mESC state to mEpiSC state in hypoxic condition and observed its effects for establishing EpiSC-specific features.
Materials and Methods
All procedures related to animal treating were approved by the Institutional Animal Care and Use Committee at Kinki University.
Establishment and maintenance of nEpiSCs
C57BL/6J strain mouse (CLEA Japan, Inc) and TgN(deGFP)18Imeg strain mouse (RBRC:00821, Oct4ΔPE-GFP) were provided by RIKEN BRC [24]. The epiblast fragments were isolated by tweezers from preimplantation embryos E5.75. Then, these fragments were cultured on mitomycin C-treated mouse embryonic fibroblasts (MEF) in EpiSC medium, which consisted of Dulbecco's modified Eagle's medium (DMEM)/F12 including 1× MEM nonessential amino acids, 0.1 mM β-mercaptoethanol, 200 mM l-glutamine, and 20% knockout SR (all were purchased from Invitrogen Corporation) supplemented with 6 ng/mL bFGF (Wako Pure Chemical Industries, Ltd.). After 4 days of culture, the EpiSC colonies were treated with 0.25% trypsin −0.04% EDTA in PBS(−) (trypsin-EDTA) and replated onto new MEF in EpiSC medium supplemented with 10 μM ROCK inhibitor Y27632 (Wako Pure Chemical Industries) to prevent dissociation-induced apoptosis. The medium was changed every day and passaged every 2–3 days. (Fig. S1; Supplementary Data are available online at
Conversion from ESC state to EpiSC state
For conversion of ESCs to EpiSCs, the Oct4ΔPE-GFP ES cells were cultured on MEF in an ESC medium, which consisted of Knockout DMEM including 1×MEM nonessential amino acids, 0.1 mM β-mercaptoethanol, 200 mM l-glutamine, and 20% knockout SR supplemented with 1,000 units/mL ESGRO (Millipore). After 2 days, these cells were treated with trypsin-EDTA solution for separating to single cells. These cells were then replated on MEFs in an EpiSC medium supplemented with ROCK inhibitor Y27632 at 37°C under an atmosphere of 5% CO2–20% O2 (normoxia) or 5% CO2–5% O2 (hypoxia). The ROCK inhibitor was only supplemented with culture medium on the first day when passaged. Characterizations of converted cells were performed after 3 times passages in each oxygen concentration.
The conversion method for EpiSC to ESC (reversion) refers to a previous report [25]. EpiSCs or induced EpiSCs (iEpiSCs) were trypsinized to single cells, and replated on MEF under ES medium supplemented with MEK inhibitor PD0325901, GSK3β inhibitor CHIR99021, and Tgfβ inhibitor A83-01 (all from Stemgent) for 2 weeks.
Immunofluorescent staining
Each cell was fixed in 10% formalin neutral buffer solution and permeabilized in 0.2% Triton-X (Sigma-Aldrich Corporation)/PBS(−) for 5 min. Then, dishes were blocked by incubation in 10% Block Ace (Dainippon Sumitomo Pharma)/PBS(−) for 1 h, and incubated with primary antibodies overnight at 4°C. The antibodies were anti-Oct4 (Santa Cruz Biotechnology; diluted at 1/100 by PBS(−) containing 10% Block Ace). For immunofluorescence of cell surface markers E-cadherin, N-cadherin, and SSEA1 (all from Santa Cruz Biotechnology), fixed cells were washed with PBS(−) and blocked by incubation in 10% Block Ace/ PBS(−) for 1 h. Then the primary antibodies diluted at 1/100 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace were added and incubated at 4°C for overnight. After that, the samples were washed twice with PBS(−) and reacted with rhodamine-conjugated anti-goat IgG antibody, FITC-conjugated anti-rabbit IgG antibody, or TexasRed-conjugated anti-mouse IgM antibody (all from Santa Cruz Biotechnology; diluted at 1/3,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace) for 1 h at room temperature. Samples were stained with DAPI (Vector Laboratories Ltd.) to counterstain the chromosomes before microscopic observation. For alkaline phosphatase staining, we used an alkaline phosphatase kit (Sigma-Aldrich) according to the instructions provided.
Producing of ubiquitously RFP-expressing ESCs and EpiSCs
For tracing analysis, we introduced a monomer RFP reporter gene to ESCs and EpiSCs. To create the pCAG-DsRed-IRES-Puro, the DsRed monomer sequences obtained from pDsRed-monomer-N1 vector (Clontech Laboratories, Inc.) was then cloned into the EcoRI/NotI site of the pCAG-IRES-Puro plasmid, which was kindly provided by Dr. Hirofumi Suemori. The cells were dissociated to single cells by Trypsin-EDTA treatment and resuspended in PBS(−) with the linearized pCAG-DsRed-monomer plasmid (mRFP). The cell suspension was then transferred to a Gene Pulser cuvette (0.4-cm gap), electroporated with Gene Pulser II (Bio-Rad Laboratories) at 200 V, 900 μF, and then plated on gelatin-coated dishes. After 24 h, 0.4 μg/mL Puromycin (Invitrogen) was added into culture media for drug selection.
Assessment of contribution into blastocyst formation by aggregation with zona-free embryos
Female ICR mice, 8 to 15 weeks old, were superovulated by intraperitoneal injection of 5 IU of pregnant mare serum gonadotropin (1,000 U; Sankyo Lifetech) followed 48 h later by 5 IU of human chorionic gonadotropin (Veterinary Puberogen, 1,500 U; Sankyo Lifetech). The females were then paired with ICR males of proven fertility. Pregnancy was confirmed by the presence of a copulated plug. Two-cell-stage embryos were flushed from the oviduct using M2 medium with 3 mg/mL of bovine serum albumin. Embryos were cultured in microdrops of mKSOM under mineral oil (Fisher Scientific) at 37°C under an atmosphere of 5% CO2 in air. After 24 h, the embryos that developed to the 4- to 8-cell stages were treated with Acid Tyrode's solution (Sigma-Aldrich) for removing the zona pellucida. For preparation of the cells, ESCs, EpiSCs, or iEpiSCs passaged 3 times in normoxic or hypoxic condition were treated with CTK solution [26] for 3 min and dissociated to approximately 10 cell clusters by gentle pipetting. Then, a cell clump and a zona-free embryo were transferred to the same well and cultured to the blastocyst stage.
DNA methylation analysis
DNA methylation status of ESC, EpiSC, Normoxia-iEpiSC, and Hypoxia-iEpiSC was determined by the bisulfite sequencing method. Detailed protocol including polymerase chain reaction (PCR) conditions is described in our previous study [27]. The primer sequences were as follows (listed 5′ to 3′ in the order of forward primer, reverse primer): ATTTTGTGATTAGGGTTGGTTTAGAA and CCAAAACATCCTCTTCATCTTTCTTCT for the Stella gene, GTAAGAATTGAGGAGTGGTTTTAGAAATAA and ACCAAATATCCAACCATAAAAAAAATAAAC for the Oct4 gene.
Real time PCR analysis
Total RNA of samples were collected using a TRIzol reagent (Invitrogen) and reverse transcribed with the High Capacity cDNA reverse transcription kit (Applied Biosystems). Quantitative real-time PCR with total cDNA was performed using Perfect real-time SYBR green II (Takara Bio Inc). PCR amplifications were performed with the 7700 real-time PCR System (Applied Biosystems) at 95°C for 20 s followed by 40 cycles of 95°C for 5 s and 60°C for 30 s. To quantify the relative expression of each gene, the Ct (threshold cycle) values were normalized by β-actin (ΔCt=Ct target − Ct β-actin) and compared with a calibrator, using the “ΔΔCt method” (ΔΔCt=ΔCt sample − ΔCt calibrator). All values are means±SD of 3 experiments. Statistical significances were evaluated by Student's t-test with JMP software version 7.0.2 (SAS Institute). The primer sequences are described in Table 1.
Western blot analysis
All samples were homogenized in a sodium dodecyl sulfate (SDS) buffer (4% SDS, 125 mM Tris–glycine, 10% β-mercaptoethanol, and 2% bromophenol blue in 30% glycerol), and centrifuged at 10,000 rpm for 10 min at 4°C to remove debris. Aliquots were subjected to polyacrylamide gel electrophoresis in the presence of SDS (SDS/PAGE) followed by electrotransfer onto a polyvinylidene fluoride membrane (Hybond-P; Amersham Pharmacia Biotech). The blotted membranes were blocked overnight with Block Ace (Dainippon Sumitomo Pharma) and then proved with an objective primary antibody overnight at 4°C. Detection was performed with horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology) and ECL plus western blotting detection system (GE Healthcare UK Ltd.). The antibodies were anti-Oct4 goat IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/5,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace), anti-Nanog rat IgG polyclonal antibody (diluted at 1/3,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace; eBioscience, Inc), anti-Sox2 goat IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/5,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace), anti-Klf4 rabbit IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/5,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace), anti-LIFR rabbit IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/3,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace), anti-E-cadherin rabbit IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/3,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace), anti-N-cadherin rabbit IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/3,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace), anti-Actin goat IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/3,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace), anti-Stat3 mouse IgG monoclonal antibody (BD Bioscience; diluted at 1/3,000 by Immunoenhancer; Wako Pure Chemical Industries, Ltd.), anti-phopshorylated Stat3 rabbit IgG polyclonal antibody (Cell Signaling technology, Inc.; diluted at 1/3,000 by Immunoenhancer), and anti-Actin goat IgG polyclonal antibody (Santa Cruz Biotechnology; diluted at 1/3,000 by PBS(−) containing 0.1% Tween-20 and 10% Block Ace). The lumino-labeled membranes were analyzed by CCD-based chemiluminescent analyzer LAS4000 (GE Healthcare Japan). All values are means±SD of 3 experiments. Statistical significances were evaluated by Student's t-test.
Flow cytometry analysis
Normoxia- and Hypoxa-iEpiSCs at passages 1, 3, and 6 were dissociated into single cells by Trypsin-EDTA, blocked by incubation with FACS buffer (2% FCS and 10 mM HEPES in DMEM) for 15 min at 4°C and incubated with anti-SSEA1 antibody (diluted 1/100 by FACS buffer) for 1 h 30 min at 4°C. Then, the samples were washed twice with the FACS buffer and incubated in PE-conjugated anti-mouse IgM secondary antibody (Santa Cruz Biotechnology; diluted at 1/1,000 by FACS buffer) for 1 h. SSEA-1+/Oct4ΔPE-GFP+ and SSEA-1+/Oct4ΔPE-GFP− cell fractions were sorted with FACS Vantage SE (Becton Dickinson) and analyzed by flow cytometry with a FACS Caliber cytometer (Becton Dickinson). All values are means±SD of 3 experiments. Statistical significances were evaluated by Student's t-test.
Results
Observation of the conversion from ESC state to EpiSC state using Oct4ΔPE-GFP ESCs
Pluripotent cell marker gene Oct4 promoter activity is controlled by 2 enhancer regions: the proximal enhancer is silenced in the ICM and germ lineages, and the distal enhancer (DE) is silenced in the epiblasts. Oct4 promoter-lacking PE regions (Oct4ΔPE) become active only in ES cells and PGCs, but not in epiblasts; therefore, the converted cells to the epiblast state are observed by diminished GFP fluorescence [28]. Here we produced Oct4ΔPE-GFP ESCs, and first observed the spontaneous conversion from ESCs to EpiSCs in atmospheric (O2=20%) condition by transferring the ESCs to EpiSC-culture conditions containing bFGF and deprived LIF. By culturing in the EpiSC-condition, colony morphologies of these altered EpiSC-like flatter and GFP expression declined (Fig. 1A, B). To confirm that the GFP-negative cells contained in the flat colonies converted to EpiSC-like features, we sorted the GFP-negative stem cell-fraction using an antibody to pluripotent cell surface marker SSEA1 antibody to eliminate contamination of the differentiated cells and analyzed it after culture (Fig. 1C). SSEA1+/Oct4ΔPE-GFP+ fractions formed ESC-like dome-shaped colonies in LIF-added ES-culture condition (Fig. 1D). On the other hand, SSEA1+/Oct4ΔPE-GFP− negative fractions did not proliferate in ES-conditions and formed flat colonies in EpiSC-condition (Fig. 1D). Consistent with the original descriptions of EpiSCs, the iEpiSCs showed pluripotency markers OCT4, NANOG, and SOX2 (Fig. 1E) expression and specific gene expression, that is, lower expression of Klf4, Piwi2, and Stra8, and higher expression of the postimplantation markers Eomes and Lefty1 than ESCs (Fig. 1F).

Induction of EpiSC state from Oct4ΔPE-GFP ESCs by transfer in EpiSC-culture medium.
Hypoxia accelerate the ESC-EpiSC conversion
Then, we transferred the Oct4ΔPE-GFP ESCs to an EpiSC-condition in 2 different oxygen concentration conditions (normoxia=atmospheric 20% O2 concentration, hypoxia=5% O2 concentration). Oct4ΔPE-GFP expression declined rapidly in hypoxic condition and the conversion was also observed in the ES-condition (Fig. 2A). When evaluating the efficacy of the conversion in hypoxic environment by calculating the remaining rate of Oct4ΔPE-GFP+ cells per SSEA1+ cells, the culture in hypoxia accelerated ESC-EpiSC conversion (Fig. 2B) compared with the normoxic condition. The iEpiSCs in hypoxic condition (hypoxia-iEpiSC) displayed the marker profile of EpiSCs clearly: Oct4 expression, reduced expression of Nanog, Sox2, and Klf4, and diminishing ICM-specific marker expression Stra8 and Piwi2 (Fig. 3A, B). Further, complete reduction of E-cadherin, which is a major marker of mESC and ICM was observed, alternatively switched to N-cadherin expression, which shows that epiblast-specific features were detected (Fig. 3B, C). The alteration of the cadherin expression was not complete in normoxia-iEpiSCs, it might be a specific feature of hypoxia-iEpiSCs. When epigenetic statuses of the promoter regions of Oct4 and Stella were observed and compared among ESCs, nEpiSCs, hypoxia-EpiSCs, and normoxia-EpiSCs, hypoxia-EpiSCs clearly had the closest pattern to nEpiSCs (Fig. 3D). Then, to determine the developmental properties of hypoxia-iEpiSCs in vivo, we observed the contribution to blastocyst formation after morula aggregation. We found that after morula aggregation, hypoxia-iEpiSCs could not integrate to ICM cells in blastocysts. Consistent with this, no contribution was detectable in fetuses or pups after embryo transfer (Fig. 4).
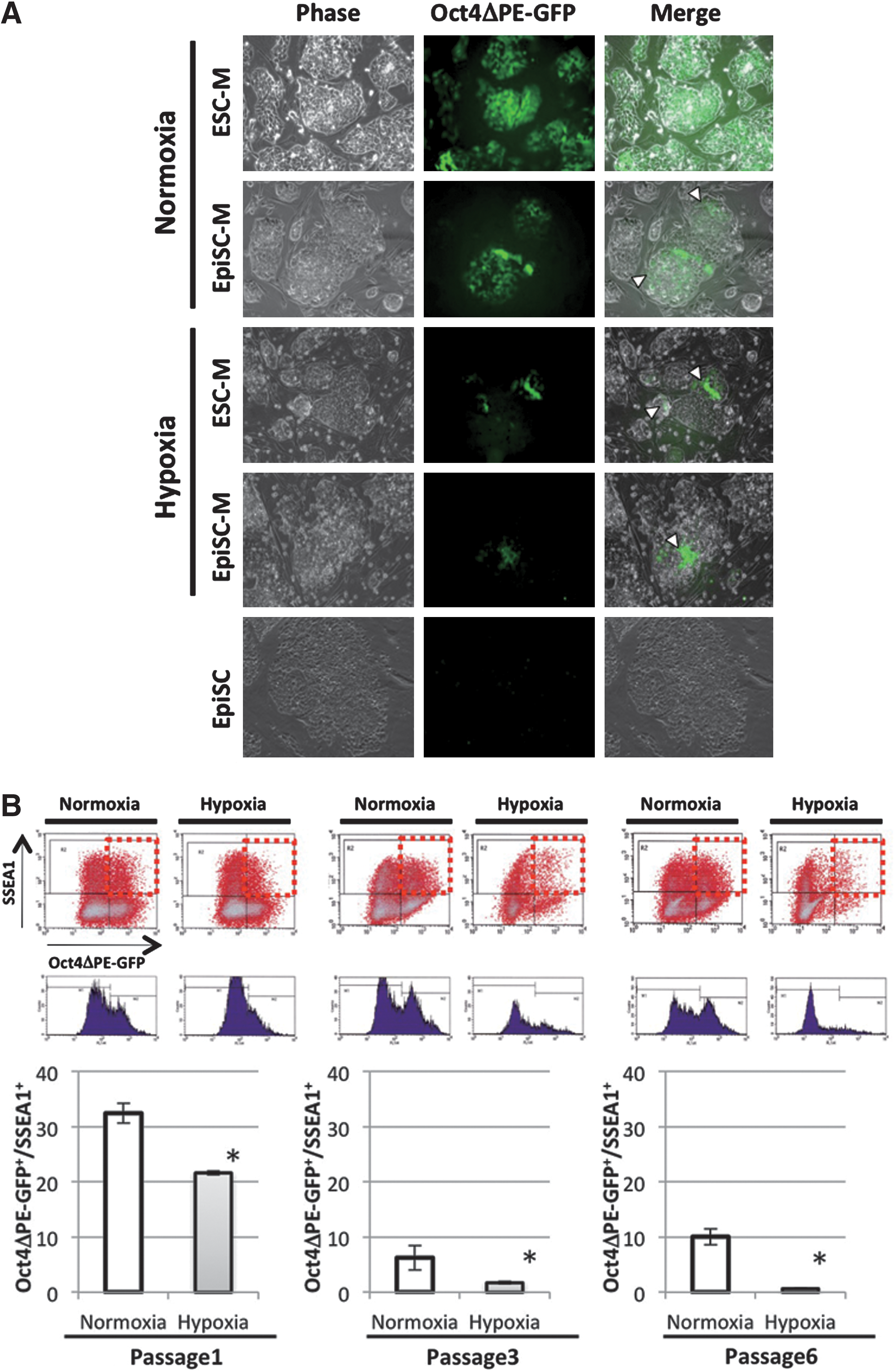
Hypoxia accelerates ESC- to EpiSC state conversion.
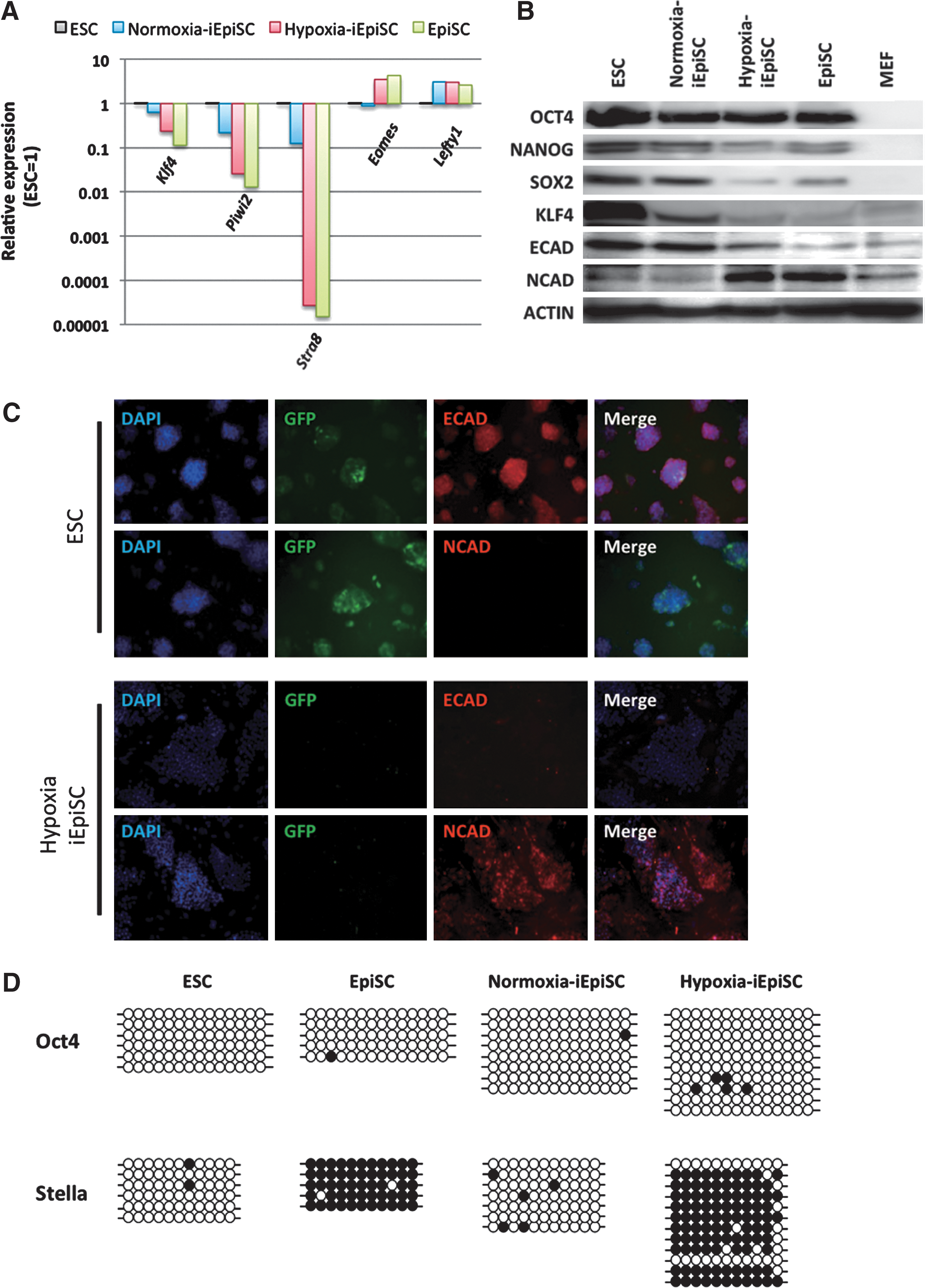
Detailed characterization of the iEpiSCs passaged 3 times in normoxic or hypoxic conditions.
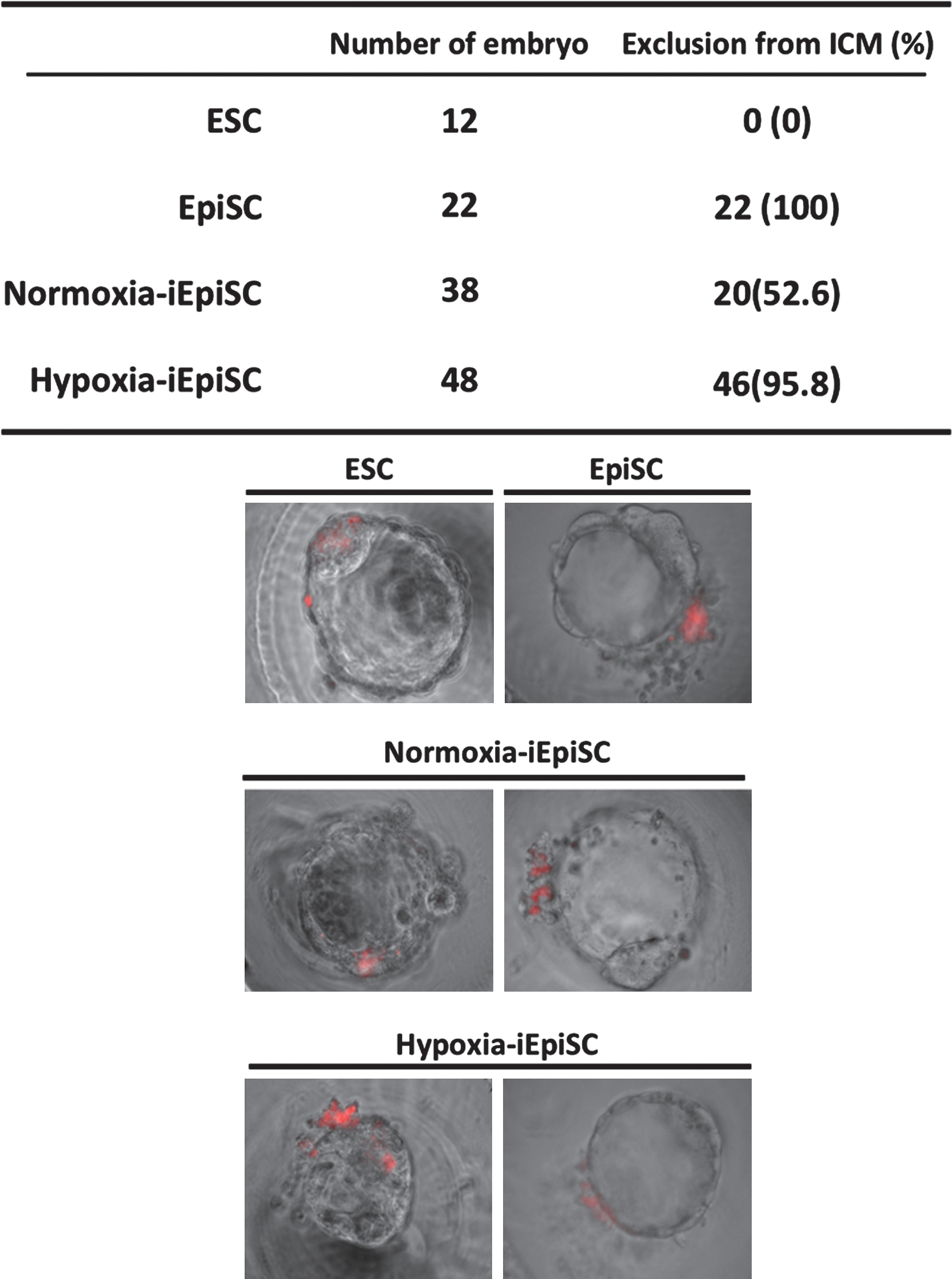
Observation of the contribution properties into blastocyst formation. In control experiments, ESCs were contributed into ICM in all cases, but EpiSCs were excluded from blastocoel. Consistent with the results of the native-EpiSCs, hypoxia-iEpiSCs did not contribute to ICM in almost cases. ICM, inner cell mass.
Hypoxia positively affects EpiSC state conversion than the condition in which LIF-Stat3 signaling was inhibited
It has been described that the repression of LIF receptor gene expression is one of the hypoxic effects in pluripotent cells. Therefore, we next examined the effect of hypoxia on the conversion to EpiSC state by comparing with the condition using JAK1 inhibitor (JAKi) that blocks LIF-Stat3. Consistent with a previous study, LIFR and its down-stream phosphorylated Stat3 (Y705) were repressed in the converting cells in hypoxic condition (Fig. 5A). Hypoxia-related LIF-Stat3 inhibition occurred also in the LIF-added ESC culture condition. In the cells cultured in the JAKi-supplemented culture condition, the phosphorylation of Stat3 was completely inhibited (Fig. 5B). Interestingly, the iEpiSCs obtained from JAKi-added culture conditions (JAKi-iEpiSC) remained ICM specific gene expression such as Klf4, Piwi2, and Stra8 (Fig. 5C). Finally, we examined the stability of the gained EpiSC state in each condition. Oct4ΔPE-GFP+ ESC-like colonies were recovered easily from normoxic-iEpiSCs and JAKi-iEpiSC (∼15% in one typical experiment) only by transferring them into the LIF-added ESC-condition (Fig. 6A, B). On the other hand, this spontaneous reversion was never observed in hypoxia-iEpiSCs. It showed that the ES-EpiSCs conversion occurring in the hypoxic condition was more stable than in normoxic conditions. When forced conversion condition by combination of ALK, MAPK, and GSK3β inhibiters, which has already been reported to enable the conversion of nEpiSCs to an ESC state, was adopted, hypoxia-iEpiSCs exhibited a wave of differentiation and cell death, few Oct4ΔPE-GFP+ colonies emerged after 2 weeks (Fig. 6B) as a result of this conversion.
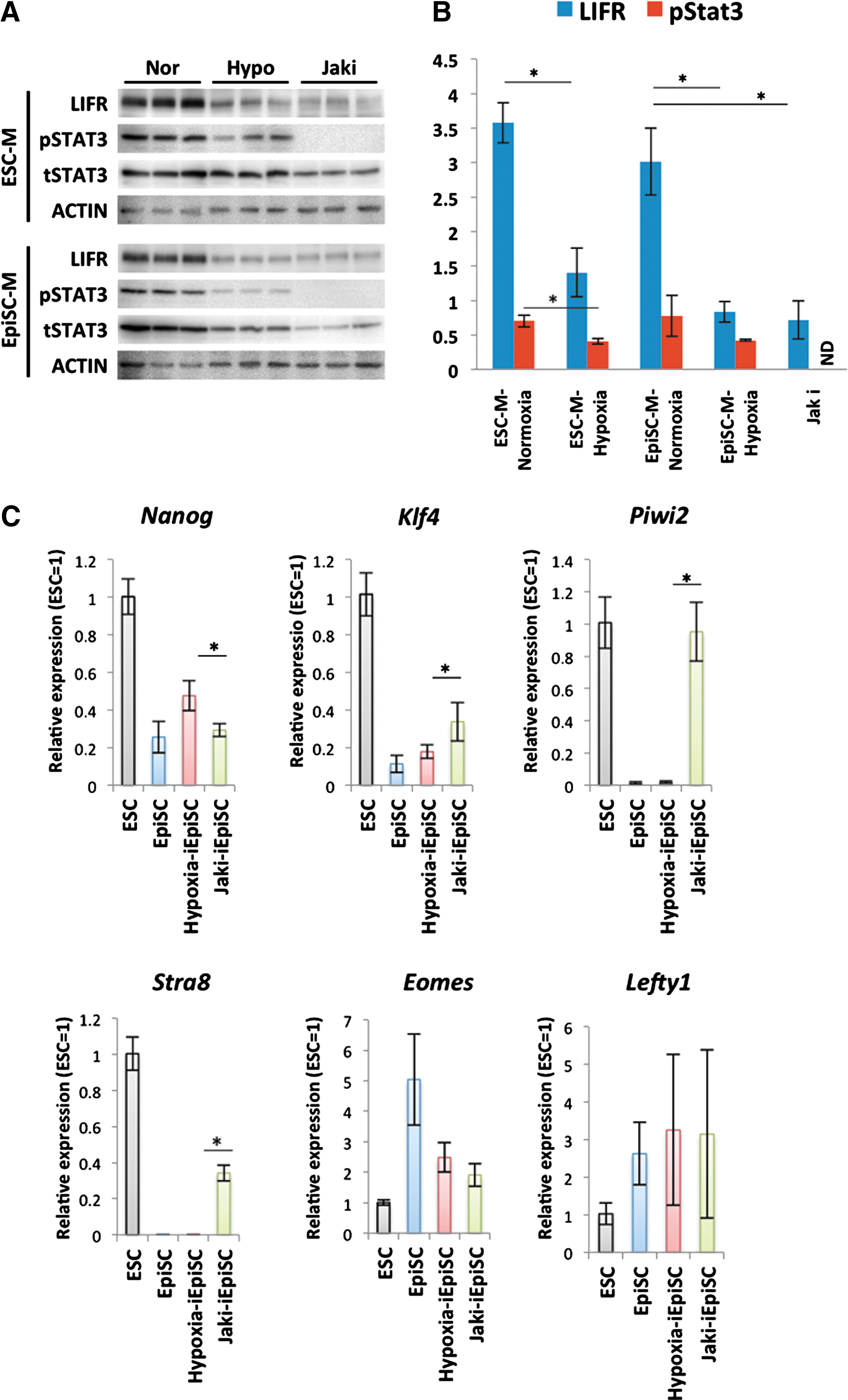
Comparison between the hypoxia-iEpiSCs and JAKi-iEpiSCs.

Spontaneous or forced reversion from EpiSC state to ESC state. Arrowheads show the Oct4ΔPE-GFP positive cells.
Discussion
Oxygen concentration is an important factor for tissue formation, differentiation, and function [19]. Here we examined the effect of hypoxic treatment on conversion from a mESC state to an EpiSC state. When the ESCs were cultured in LIF-/bFGF+ condition, some parts of the cells lost Oct4ΔPE-GFP expression, which represents DE activity-derived Oct4 expression [28]. The converted iEpiSCs were obtained as SSEA-1+/Oct4ΔPE-GFP− fraction: these cells expressed epiblast markers, whereas expression of ICM-specific marker genes declined. The iEpiSCs induced in normoxic condition (normoxia-iEpiSCs) represented some EpiSC-specific gene expression, but they easily reverted to an ESC state distinguished by dome-shaped colonies and DE activity-derived Oct4 expression that is shown by Oct4ΔPE-GFP expression. On the other hand, the iEpiSCs induced in the hypoxic condition (hypoxia-iEpiSCs) possessed more similar characteristics to epiblast-derived EpiSCs (nEpiSCs) than normoxia-iEpiSCs in terms of the quantity of marker gene expression and pattern of cadherin expression. These results clearly show that a hypoxic environment has a positive effect on establishing an EpiSC state. Interestingly, the hypoxic condition also affected the epigenetic status in the converted cells. At least the promoter region of Stella, which is a mESC state marker, was highly methylated only when the conversion was performed in the hypoxic condition. Further, the EpiSC state of hypoxia-iEpiSCs was more stable than normoxia-iEpiSC; no spontaneous reversion was observed if it was transferred in the ESC-culture condition. In other words, the effects of hypoxia did not only accelerate and promote ESC-EpiSC conversion, but improved and stabilized the EpiSC state.
So then, what factor stabilizes the EpiSC state in hypoxia? In a hypoxic environment, hypoxia-inducing transcription factor families hypoxia inducible factors (HIFs) were stabilized and participated in regulation of various gene expression. Jeong et al. reported that LIF-Stat3-dependent ESC self-renewal is inhibited in a hypoxic condition because stabilized HIF1 represses LIFR expression [23]. It has already been reported that inhibition of JAK, which is a down-stream molecule of LIF-Stat3 signaling, improves pluripotency and specific features of mouse EpiSCs [11]. Therefore, at first, we thought it was most plausible that the positive effect of hypoxia on the EpiSC state might have resulted by simply inhibiting the LIF-Stat3 signaling pathway. However, given the fact that the epiblast state of the hypoxia-iEpiSCs was more stable than iEpiSCs produced by JAK inhibition, other mechanisms promoting the EpiSC state might exist. Until recently, some researchers described the positive effect of hypoxia on hESCs. Ezashi et al. reported that hypoxic (3%–5% O2) cultures reduce spontaneous differentiation of hESC cultures and it could contribute to remaining their pluripotent state [21]. Forristal et al. also suggested that the hypoxic condition has a positive effect on hESC pluripotency [29]. According to their description, HIF 2, which is also a transcription factor stabilized by hypoxic condition, upregulates Oct4, Nanog, and Sox2 expression and leads to the enhancement of pluripotency. Moreover, some descriptions of other types of cells also suggest the mechanisms of the effect connected to hypoxia on the pluripotent-related signaling cascade. For example, Zhang et al. showed that hypoxic condition increases HIF1 stabilization and leads to smad3 activation in human umbilical vein endothelial cells [30]. Recently, Chen et al. actually demonstrated that hypoxia induced activation of smad2/3 signaling cascade also occurred in hESCs and it enhances their self-renewal [22]. Further, HIF2 activation brings cell cycle progression and proliferation via β-catenin phosphorylation in the human renal cancer cell line [31]. The inhibition of GSK3β and activation of β-catenin lead to promotion of both mESC and hESC self-renewal via enhancement of Nanog expression is well known [32]. On the other hand, it has already been clearly demonstrated that smad1/5 inactivation and smad2/3 activation strongly enhance hESC pluripotency, whereas they prevent mESC self-renewal [12]. Taken together, although hypoxia promotes the common molecule for mESC-, mEpiSC-, and hESC pluripotency such as Oct4, Nanog, Sox2, and β-catenin, the mESC state cannot be maintained by inhibition of vital signaling cascades such as LIF-Stat3 or Smad1/5. Promotion of the conversion and stabilization of the EpiSC state observed in this study may have resulted from the correlation of the above signaling cascades and selective enhancement of mEpiSC-type pluripotency.
It is considered that determination of the factors related to ESC-EpiSC conversion provides valuable information not only for improving culture systems of pluripotent stem cells but also to elucidate its developmental origins and detailed mechanisms for formation of the specific features of each state. From this perspective, the present results that the hypoxia enhanced and stabilized epiblast state may have important meanings. Oxygen concentrations in reproductive tracts have been measured in a previous study [20]. Oviduct includes O2 at over 8.7%, whereas the environment in uterus is very hypoxic; there is only 1.5%–2% O2. The epiblast in the embryonic disk is located within the uterine wall, so that the speculative physiological environment of EpiSCs may originally be hypoxic. As described above, some hypoxia-specific transcription factors such as HIF1 are stabilized in low O2 condition and function to regulate expression of many important genes for tissue development. It was described that the HIF1 stabilization is initiated around 5% O2 concentrations [33], and actually a high amount of HIF1 accumulation was observed also in the present hypoxic condition (data not shown). In other words, at least the gene expression environment of the present study may successfully simulate the culture condition of uterus. And as a result of this, in hypoxic conditions, ESCs convert EpiSC state more naturally. Some previous reports have described the reactions of hESCs to hypoxia. It clearly reflects their adaptability in a hypoxic environment, and may also strengthen the idea that hESCs possess characteristics of postimplanted embryonic cells and are developmentally close to EpiSCs.
In conclusion, the present study provides evidence that oxygen concentration is an important factor for the promotion and stabilization of the EpiSC state. Determination of the factors and signal cascade enhancing the conversion to EpiSC state or increasing EpiSC state stabilities in future studies are essential in correctly understanding the developmental stages and properties of pluripotent stem cells, and to develop a method which enables ESC to shuttle to an EpiSC state, and EpiSC to shuttle to an ESC state; there must be a more ideal model system for studying human development. These conversion–reversion systems may also contribute to the expansion of the availability of human pluripotent stem cells.
Footnotes
Acknowledgments
We gratefully acknowledge Dr. Hirofumi Suemori, Institute for Frontier Medical Science, Kyoto University, for kindly providing pCAG-IRES-Puro plasmid. We also thank Ms. Naomi Backes Kamimura and Ms. Julia Walhelm-Kimura, Department of Biology-Oriented Science and Technology, Kinki University, for English editing.
Author Disclosure Statement
The authors have nothing to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.