
Abstract
Embryonic stem (ES) cells give rise to mesodermal progenitors that differentiate to hematopoietic and cardiovascular cells. The wnt signaling pathway plays multiple roles in cardiovascular development through a network of intracellular effectors. To monitor global changes in wnt signaling during ES cell differentiation, we generated independent ES cell lines carrying the luciferase gene under promoters that uniquely respond to specific wnt pathway branches. Our results show that successive, mutually exclusive waves of noncanonical and canonical wnt signaling precede mesoderm differentiation. Blocking the initial noncanonical JNK/AP-1 signaling with SP60125 aborts cardiovascular differentiation and promotes hematopoiesis, whereas interference with the subsequent peak of canonical wnt signaling using Dkk1 has the opposite effect. Dkk1 blockade triggers counter mechanisms that lead to delayed and extended activation of canonical wnt signaling and mesoderm differentiation that appear to favor the cardiomyocytic lineage at the expense of hematopoietic cells. The cardiomyocytic yield can be further enhanced by overexpression of Wnt11 leading to approximately 95-fold enrichment in contracting cells. Our results suggest that the initial noncanonical wnt signaling is necessary for subsequent activation of canonical signaling and that the latter operates under a regulatory loop which responds to suppression with hyperactivation of compensatory mechanisms. This model provides new insights on wnt signaling during ES cell differentiation and points to a method to induce cardiomyocytic differentiation without precise timing of wnt signaling manipulation. Taking into account the heterogeneity of pluripotent cells, these findings might present an advantage to enhance the cardiogenic potential of stem cells.
Introduction
E
Wnt (wingless-type MMTV integration site) signaling in mammals is mediated by the interaction of 19 distinct ligands with 10 Frizzled (Fzd) 7-transmembrane receptors [10]. Receptor activation regulates a battery of cellular processes including transcriptional activation of select target genes, proliferation, differentiation, migration, and polarity [11]. Wnt signaling has been traditionally divided into 3 branches, the canonical wnt branch, whose major downstream mediator is β-catenin, and 2 noncanonical branches, the first modulating the activity of Rho and JNK kinases, the second leading to increased intracellular levels of Ca2+ and subsequent activation of calcineurin and nuclear factor of activated T-cells (NF-AT) transcription factors [10].
An important component of wnt signaling is the existence of coreceptors such as members of the low-density lipoprotein-related proteins (LRP) that are required for canonical signaling, and secreted antagonists that interfere with the binding of wnt ligands to their receptors. Wnt antagonists are mainly of 2 types, those that allow binding of ligands to Frz receptors, but hinder the interaction between Frz and LRP partners such as members of the Dickkopf (Dkk) family, and, secreted frizzled-related proteins (sFRPs) that sequester wnt ligands in the extracellular space preventing binding to receptors [12]. Consequently, Dkk proteins block canonical signaling, which depends on LRPs coreceptors, and channel receptor activation to noncanonical wnt pathways. In contrast, sFRPs are expected to block all signaling branches.
Wnt proteins have pleiotropic effects on cardiac morphogenesis and cardiomyocyte differentiation [13 –15]. During development, canonical wnt signaling activation before mesoderm formation enhances cardiomyocytic differentiation. In contrast, canonical wnt signaling hinders cardiomyogenesis after gastrulation, thus suggesting a biphasic role in cardiac differentiation. Accordingly, Dkk1 has the opposite effects, that is, promoting cardiogenesis at late stages, but blocking it early on [16]. Manipulation of wnt signaling during ES cell differentiation can either hinder or enhance cardiac differentiation, often leading to apparently conflicting effects, thus suggesting that responses may vary depending on the stem cell line used, culture conditions, as well as the timing and duration of wnt signaling stimulation or inhibition [14,16 –23].
Our previous genomic analyses identified clusters of coordinately expressed genes during ES cell differentiation that include, in order of timing of expression, (1) genes which take part in the formation of the 3 embryonic germ layers during gastrulation, that is, Cerberus like 1 homolog, T-brachyury, and Wnt3; (2) molecular regulators of mesoderm development, including Bmp2, Bmp5, Vegfr2, and Foxf1; (3) genes linked to hematopoiesis such as Gata1 and hemoglobins; (4) transcripts involved in cardiac development, for example, Mef2c, Myl4, and cardiac Troponin T2; and (5) genes associated with neuronal development, such as Neurog1, Neurog2, and EphA5 [24]. To place changes in wnt signaling within the framework of this orchestrated differentiation process, we generated 3 independent ES cell lines, each carrying the luciferase gene under a promoter that responds to activation of a separate pathway branch activation, namely canonical/β-catenin, Ca2+/NF-AT, and JNK/AP-1.
Our results show that noncanonical wnt signaling is activated at the onset of ES cell differentiation before the formation of cells representing the 3 embryonic germ layers. Noncanonical activity is followed by a strong transient induction of canonical wnt signaling that coincides with expression of gastrulation markers and extends to the early differentiation stages of mesodermal progenitor cells. We found that blockade of the initial noncanonical JNK/AP-1 activity hinders differentiation of endothelial cells and cardiomyocytes and favors hematopoiesis. In contrast, antagonism of canonical wnt signaling by Dkk1 has the opposite effect enhancing expression of cardiomyocytic and endothelial cell markers at the expense of hematopoiesis specific genes. Although Dkk1 initially blocks canonical wnt activity, it eventually leads to a delayed and prolonged counter activation of canonical wnt signaling that shifts the time window marked by the appearance of mesodermal progenitor cells. Thus, in certain pluripotent ES cell lines, it is possible to stimulate cardiovascular cells through constant antagonism of canonical wnt signaling by Dkk1.
Materials and Methods
ES cell culture and differentiation
CGR8 ES cells were grown in GMEM medium (Sigma-Aldrich;
RNA preparation and quantitative real-time–polymerase chain reaction analysis
Total RNA was extracted from ES cells by using the RNeasy kit (Qiagen;
Fluorescence-activated cell sorting
EBs created using the T-Brachyury-GFP and Nkx2.5-GFP cell lines [5,6] were washed in 1× PBS and dissociated with 0.25% Trypsin for 5 min at 37°C and dispersed to single cells by gentle pipetting. Cells were centrifuged for 3.5 min at 3,500 rpm, resuspended in 500 μL 1× PBS containing 1:1,000 dilution of propidium iodide for viability staining, and analyzed by flow cytometry. For fluorescence-activated cell sorting (FACS) analysis of day 14 EBs, we used 0.2 mg/mL Collagenase type 2 (Worthington;
Luciferase assays
ES cells undergoing differentiation in hanging drops were washed with 1× PBS, spun and dissolved in 100 μL 1× reporter lysis buffer (Promega;
Immunofluorescence
CGR8 cells were cultured and differentiated as just described. On day 4, 5 EBs/well were transferred to chambered slides. On day 6 for β-catenin, and day 14 for α-actinin, cells were washed with 1× PBS and fixed with 4% paraformaldehyde for 15 min at room temperature, washed thrice with 1× PBS for 5 min each, permeabilized with 0.2% Triton X-100 in 1× PBS for 30 min at room temperature, and washed again thrice with 1× PBS. Samples were blocked with 1 mg/mL bovine serum albumin (BSA) in 1× PBS for 30 min at room temperature followed by primary antibody incubation overnight at 4°C. Next day, cells were washed thrice with 1× PBS, incubated with secondary antibodies for 1 h at room temperature, and washed with 1× PBS. Slides were then sealed with mounting medium (Vector Laboratories;
Generation of stable ES cell lines
We used the Lipofectamine 2000 reagent (Invitrogen) to transfect CGR8 cells and generate stable clones with (1) the Myh6–Wnt11 plasmid that was created by digesting the Myh6 (a-MHC) gene promoter-polyA hGH cloning vector (kindly provided by Dr. J. Robbins [28]) with SalI and HindIII restriction enzymes. The Wnt11 full-length open reading frame fragment was obtained by PCR using CGR8 differentiation day 12 cDNA as template and primers with attached SalI and HindIII sites as follows: 5′-GT
Statistical analysis
Error bars represent average±SE. Statistical analyses were performed by using Student's t-test.
Results
Wnt signaling pathway activity during differentiation of mouse ES cells
To monitor the activity of the various branches of the wnt signaling pathway, we generated stable ES cell clones of the germline-competent CGR8 line [29] carrying the reporter luciferase gene under the control of either tandem repeats of the TCF/LEF binding sites that respond to canonical/β-catenin signaling, tandem repeats of the TPA response elements that are activated by JNK/AP-1, or tandem repeats of the NF-AT consensus binding site that is induced by mobilized NF-AT factors. Engineered CGR8 cells with 1 of the 3 indicator constructs were then dissociated and allowed to form EBs in hanging-drop suspensions until day 4. At this stage, EBs were attached to gelatin-coated plates for further differentiation. Treatment regimes were refreshed every second culture day.
The results show that undifferentiated ES cells have low, but detectable levels of canonical/β-catenin and moderate levels of JNK/AP-1 activity (Fig. 1A). At the onset of differentiation, there is an immediate drop in canonical wnt activity that mirrors a sharp increase in noncanonical/JNK/AP-1 signaling (Fig. 1A). After these initial changes in wnt signaling, at day 3 of differentiation, there is a steep and transient peak in canonical wnt signaling accompanied by a parallel decrease in the JNK/AP-1 branch (Fig. 1A). After day 4, measurable levels of Luciferase activity in both indicator lines decline to close-to-background levels, possibly reflecting the plurality of codifferentiating cell lineages and, thus, the small percentage of cells with active wnt signaling at late differentiation time points.

Activity of wnt pathways during mouse ES cell differentiation. CGR8 cells carrying the luciferase gene under the control of various wnt signaling branches were allowed to differentiate in hanging drops.
NF-AT-driven Luciferase activity (not shown) followed, in general, the pattern of JNK/AP-1, but was considerably lower by 3 orders of magnitude compared with canonical wnt signaling maxima and 2 orders of magnitude compared with JNK/AP-1, thus suggesting that JNK/AP-1 activation is the predominant noncanonical branch during these stages. Although such differences might reflect the relative strengths of the artificial reporter constructs and not necessarily endogenous levels of activation, because of the low readings of NF-AT-driven Luciferase activity, we focused further analyses on the canonical/β-catenin and noncanonical/JNK/AP-1 pathways.
To better visualize the activity of JNK/AP-1 and canonical/β-catenin signaling in relation to the ES cell differentiation processes, we plotted the expression of their respective targets together with the expression profiles of gene markers for various lineages using the FunGenES database (
Briefly, our results show sequential, mutually exclusive waves of JNK/AP-1 and canonical/β-catenin wnt signaling up to day 4 of ES cell differentiation with canonical/β-catenin after JNK/AP-1 activity. High noncanonical/JNK/AP-1 activity in the first 2 days of ES cell differentiation precedes the appearance of cells representative of ectoderm, mesoderm, and endoderm, whereas canonical wnt activity overlaps with the emergence of progenitor cells for the 3 germ layers and early mesoderm differentiation.
Interference with distinct wnt signaling branches at their corresponding peak windows alters the differentiation of mesodermal lineages in opposite ways
To test how interference with the various wnt pathway branches affects ES cell differentiation, we applied the JNK inhibitor SP60125 at days 0–2 and the canonical wnt antagonist Dkk1 at days 2–4 that correspond to peak stages of JNK/AP-1 and canonical wnt signaling, respectively. SP60125 abolished JNK phosphorylation in differentiating ES cells as expected, thus indicating a blockade of the JNK/AP-1 pathway (not shown).
SP60125 application at 0–2 results in strong suppression (20-fold) of cardiogenic and endothelial differentiation assessed at day 10, whereas hematopoiesis- and endoderm-specific genes are induced by 3.5 and 2-fold, respectively (Fig. 2A). In contrast, treatment with Dkk1 during the peak of canonical wnt activity between days 2 and 4 induces cardiac (17-fold) and endothelial (2-fold) genes and suppresses hematopoietic (2-fold) and endodermal markers (5-fold, Fig. 2A). There is also a modest 1.5-fold induction in neuronal genes such as tubulin β3 in Dkk1-treated EBs and no major changes in RNA samples from EBs treated with SP60125. Thus, it appears that interference with the initial JNK/AP-1 or the subsequent canonical wnt activity has opposite effects on the differentiation of mesodermal and endodermal lineages.
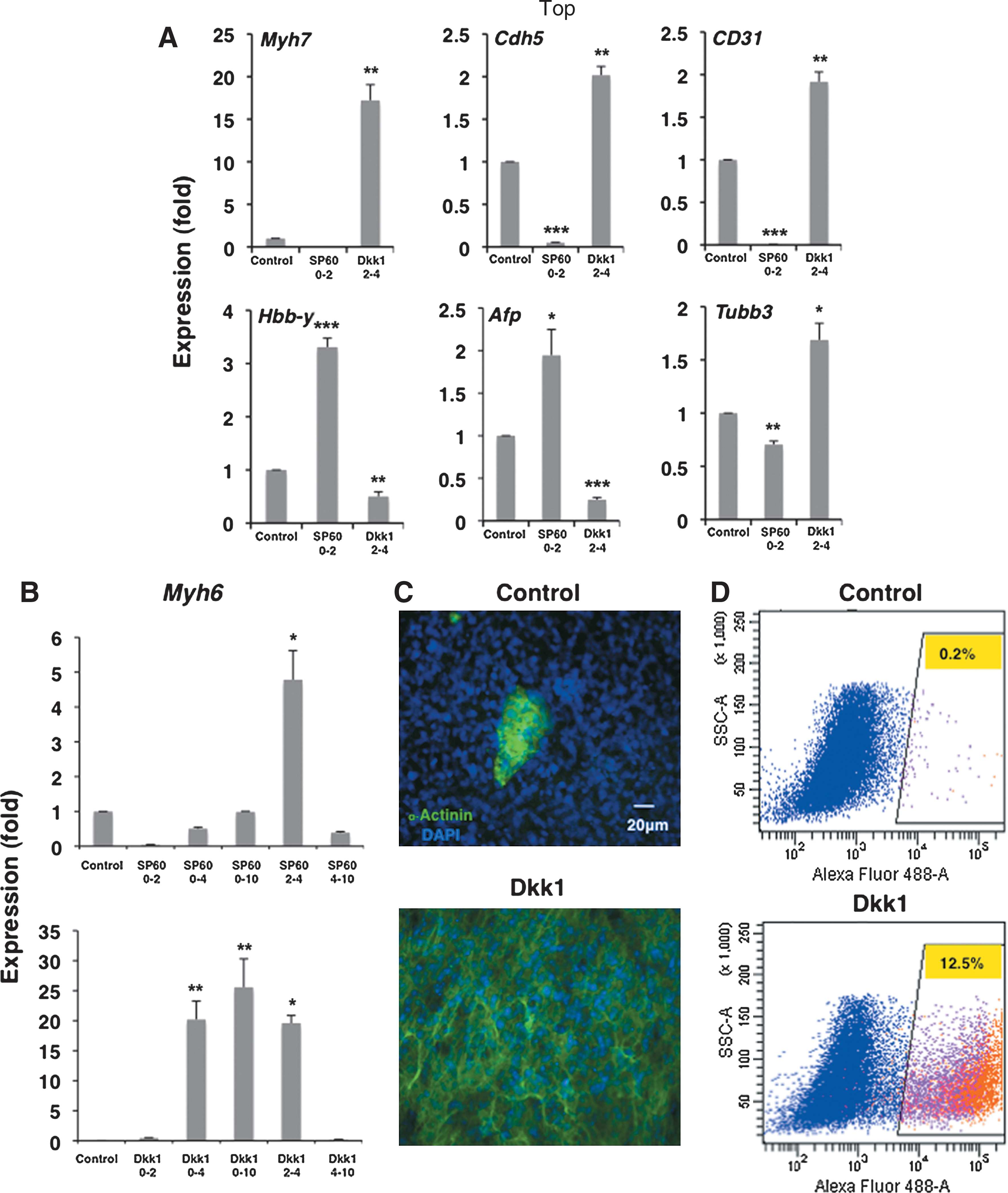
Interference with wnt signaling branches at specific time windows affects the differentiation of multiple lineages. Differentiating CGR8 ES cells were treated with the JNK inhibitor SP60125 or the canonical wnt antagonist Dkk1 at various time windows as indicated.
Taken together, our results suggest that the original JNK activity is necessary for properly carrying out subsequent differentiation steps, as inhibition with SP60125 practically abolishes myocardial and endothelial gene expression favoring instead hematopoietic and endodermal differentiation. Conversely, Dkk1 interference suppresses hematopoietic and endodermal lineages and induces endothelial and cardiomyocyte-specific gene markers.
Continuous suppression of wnt signaling by Dkk1 promotes cardiomyocyte differentiation
To test how interference with the JNK/AP-1 and canonical/β-catenin wnt signaling affects cardiomyocytic differentiation throughout the differentiation process, we applied SP60125 or Dkk1 between 0–2, 2–4, 4–10, and 0–10 days of differentiation.
The results show that although JNK blockade at its peak suppresses cardiomyocyte differentiation, application of SP60125 during the surge of canonical wnt activity at days 2–4 induces expression levels of cardiac-specific genes by 5-fold (Fig. 2B). The positive effect of JNK inhibition at days 2–4 appears to override the negative effect on days 0–2, as addition at days 0–4 and 0–10 has less of an impact on cardiogenesis (Fig. 2B).
In contrast to JNK inhibition that shows negative and positive effects on cardiomyocytic differentiation consistent with the corresponding endogenous high and low JNK/AP-1 activities, respectively, the results of Dkk1 application follow a more complex pattern. For example, addition at days 0–2 and after day 4 does not influence cardiac differentiation. Conversely, addition of Dkk1 during the peak of canonical activity between days 2 and 4 strongly induces cardiogenesis, and the same outcome is observed in the remaining 2 treatments that straddle this time point, namely Dkk1 addition between days 0–4 and 0–10 (Fig. 2B). Actually, continuous Dkk1 presence during ES cell differentiation produces the highest induction of Myh6 on day 10 of differentiation at approximately 30-fold, as measured by quantitative real-time–polymerase chain reaction (qRT-PCR) analysis.
To correlate the induction of the cardiac-specific Myh6 gene at the RNA level to the yield of cardiomyocytes, we stained control untreated and Dkk1-treated EBs with an antibody against the sarcomeric protein α-actinin. IF analysis shows a strong increase in the number of α-actinin-positive cells with sarcomeric structures that coincide with contracting areas, thus suggesting that they represent cardiomyocytes (Fig. 2C). To quantify the increase of α-actinin-positive cardiomyocytes further, control and Dkk1-treated EBs were dissociated, stained with the α-actinin antibodies, and analyzed by FACS. The results demonstrate that the percentage of α-actinin-positive cells increases from approximately 0.2% in control cultures to 12.5% with Dkk1 treatment, an increase of approximately 60-fold, in line with the molecular data that show approximately 35-fold induction in Myh6 expression and 25-fold in Myh7 (Fig. 2A, B, D).
Continuous exposure of differentiating ES cells to Dkk1 expands the original time window of noncanonical activity, but triggers delayed canonical wnt activation
Since canonical wnt signaling is required for both mesoderm specification and expansion of cardiac progenitor cells, the enrichment of cardiomyocytes in differentiating ES cells cultured under Dkk1 was apparently contradictory to the expected outcome. To further investigate the molecular basis of this phenomenon, we tested wnt pathway activities by using the CGR8 Luciferase indicator ES cell lines in the continuous presence of Dkk1 during the differentiation process. The results show that addition of Dkk1 extends the window of JNK/AP-1 for approximately 2 days, from days 0–2 to 0–4 of differentiation, but noncanonical signaling decreases thereafter (Fig. 3A). Moreover, the inclusion of Dkk1 initially reduces canonical wnt activity, but canonical wnt activity springs back at later stages even in the presence of Dkk1. Briefly, continuous Dkk1 application does not eliminate the pattern of JNK/AP-1 followed by canonical/β-catenin sequence seen in control differentiating ES cells. Instead, it creates a second window of JNK/AP-1 activity which is followed by a delayed peak of canonical/β-catenin signaling.
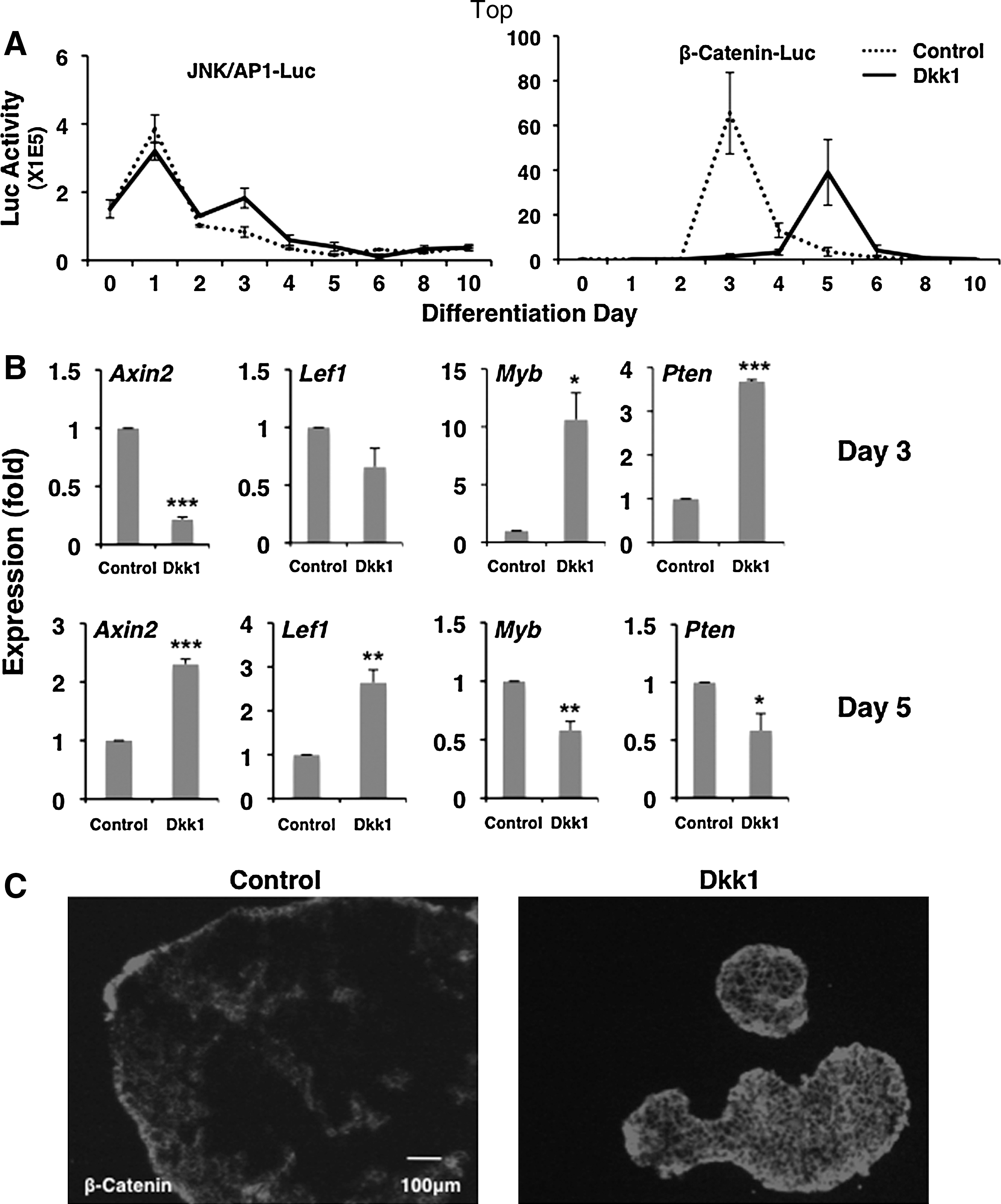
Continuous exposure of differentiating ES cells to Dkk1 expands the time window of noncanonical activity, but triggers delayed canonical wnt activation. CGR8 ES cells were cultured with or without continuous exposure to Dkk1 and analyzed for wnt pathway activity.
To test whether canonical wnt activity changes in the indicator lines extend to typical gene targets of the 2 wnt branches, we isolated RNA samples from the corresponding differentiation stages. qRT-PCR analysis showed that transcripts of known canonical wnt gene targets such as Axin2 and Lef1 are downregulated at day 3 in Dkk1-treated cultures, but are induced at day 5 of differentiation compared to untreated cells (Fig. 3B). Correspondingly, the expression of JNK/AP-1 pathway associated genes such as Myb and Pten show higher expression levels at day 3 Dkk1-treated cells than controls. The delayed activation of canonical wnt signaling also leads to increased levels of cytoplasmic and membrane-associated β-catenin as seen in IF staining of day 6 EBs (Fig. 3C). Thus, it appears that persistent challenge of canonical wnt signaling with Dkk1 leads to activation of regulatory loops that overcome the original suppression. The effect of this reaction counteracts Dkk1 antagonism, thus leading to net activation of canonical wnt signaling at later differentiation stages.
Exposure of differentiating ES cells to Dkk1 delays and expands mesoderm differentiation
As described in Fig. 1, canonical wnt activation during ES cell differentiation coincides with induction of transcripts that have been implicated in the gastrulation phase of development which is represented in day 3 EBs. To investigate how time displacement of canonical wnt signaling affects this stage, we isolated and tested RNA samples for genes, the expression of which follows the peak of canonical wnt signaling. As seen in Fig. 4A, expression of T-brachyury, Cerberus homolog-like 1, Wnt3, Wnt5a, and Wnt 8a is suppressed in the presence of Dkk1 at day 3. However, in all cases, expression strongly recovers after a delay of 1–2 days.

Continuous exposure of differentiating ES cells to Dkk1 delays and expands mesoderm differentiation. Differentiating CGR8 and T-brachyury-GFP ES cells were continuously treated with Dkk1 and analyzed by qRT-PCR and FACS analysis, respectively.
To test the effect of Dkk1 treatment on T-brachyury+ cells, we used the T-brachyury-GFP ES cell line [6]. Control, untreated T-brachyury GFP cells follow a similar, but slower differentiation program than CGR8 cells, with T-brachyury+ cells appearing at day 5 of differentiation instead of day 3 in CGR8 cells. Nevertheless, continuous exposure of T-brachyury-GFP ES cells to Dkk1 also relocated the peak of T-brachyury+ cells by 2 days, from day 5 to 7 (Fig. 4B). Thus, canonical wnt antagonism with Dkk1 temporarily suspends, but does not abolish the emergence of T-brachyury+ cells in 2 independently derived ES cell lines with distinct paces of differentiation and peaks of mesodermal gene induction.
Continuous exposure of differentiating ES cells to Dkk1 induces cardiogenic progenitor cells
Previous studies have shown 2 waves of mesodermal progenitor cells, an early wave that predominantly gives rise to hematopoietic and endothelial cells, and a second wave that yields cardiomyocytes, as well as endothelial and smooth muscle cells [30,31]. Moreover, it was also shown that cardiogenic progenitor cell expansion depends on canonical wnt signaling [18]. The fact that Dkk1 antagonism leads to delayed induction of canonical wnt signaling during cardiogenic progenitor cell specification and growth raised the possibility that the observed cardiogenic stimulation is due to the expansion of cardiac progenitor cells.
In agreement with this idea, we found that Dkk1 treatment causes a delayed and prolonged induction in the expression levels of cardiac progenitor cell (CPC) markers such as Nkx2.5, Isl1, Myocardin, and Mesp1 (Fig. 5A). Further, we found a 5–6-fold enrichment of Nkx2.5+ cells at day 6 of differentiation using the Nkx2.5-EGFP ES cell line [5] (Fig. 5B). A subset of early cardiac markers such as Isl1 and Mesp1 show a transient induction that follows the pattern of canonical wnt signaling activity, whereas Myocardin and Nkx2.5 show long-term persistent high expression throughout the tested time points (Fig. 5A). Although the significance of this split is unknown, it might reflect the persistence of select early cardiac-specific genes in cardiomyocytes. In addition, although the Mesp1 and Isl1 levels are lower in Dkk1-treated cells, their expression windows are prolonged compared with control, untreated differentiating cells. Taken together, our results suggest that the delayed canonical wnt activation, because of the presence of Dkk1, increases differentiation of CPCs.

Continuous exposure of differentiating ES cells to Dkk1 induces cardiogenic progenitor cells. Differentiating CGR8 ES cells were treated with Dkk1 and analyzed by qRT-PCR and FACS analysis.
Successive noncanonical/canonical wnt pathway activation is necessary for subsequent canonical wnt signaling and cardiac differentiation
As just described, JNK/AP-1 activity comes before canonical wnt signaling (Fig. 2A). Dkk1 application creates a second peak of JNK/AP-1 activity that subsides when the delayed canonical wnt activation takes place (Fig. 3A). To test whether this second peak of JNK/AP-1 pathway activation that precedes canonical activity in Dkk1-treated EBs is also important for the delayed canonical wnt activation and cardiomyocyte differentiation, we treated Dkk1-exposed EBs with SP60125 at days 2–4. As shown in Fig. 6, SP60125 treatment during the time window of the second, Dkk1-induced JNK/AP-1 peak attenuates the induction of mesodermal markers (T-brachyury, Wnt3, and Wnt8a), markers for cardiogenic differentiation (Mesp1, Myocardin, and Nkx2.5), and the subsequent appearance of cardiomyocyte-specific genes Myh6 and Myh7. Conversely, application of SP60125 at days 0–2 has minor effects on the cardiogenic differentiation of Dkk1-treated cells. These results suggest that the JNK/AP-1 activity immediately before canonical wnt signaling plays a role in the subsequent mesodermal and cardiogenic differentiation.

Successive noncanonical/canonical wnt pathway activation is necessary for subsequent canonical wnt signaling and cardiac differentiation. RNA analysis of differentiating CGR8 ES cells treated with Dkk1 and SP60125. Addition of SP60125 on days 2–4 just before the delayed canonical wnt activity in Dkk1-treated cells reduces cardiac marker gene expression at day 10 of differentiation. qRT-PCR values are the mean±SE of 3 separate experiments. *P<0.05; **P<0.005; ***P<0.0005; # P=0.06 compared with controls.
Dkk1 induces Wnt11 that further promotes cardiogenic differentiation
The results just described show that persistent antagonism by Dkk1 leads to a number of permanent gene expression changes such as sustained Nkx2.5 and Myocardin expression beyond the typical time windows in control ES cells. However, the fact that Nkx2.5+ CPCs are expanded by 5-fold, but cardiomyocytes by nearly 60-fold, suggested that additional changes, taking place concurrently or subsequent to CPC expansion, further enhance cardiomyocytic differentiation. Among the wnt factors that are induced by Dkk1 is Wnt11, which is known to stimulate the growth and differentiation of cardiomyocytes during development and ES cell differentiation [15] (Fig. 7A).

Dkk1 induces Wnt11 that further expands cardiogenic differentiation. CGR8 cells, transfected with Myh6-DsRed-Nuc and with or without Myh6-Wnt11, were treated with Dkk1 and analyzed for expression of cardiac genes and by fluorescence microscopy.
To test the effects of higher Wnt11 expression in conjunction with Dkk1 application on cardiomyocytic differentiation, we cloned the Wnt11 cDNA under the cardiac-specific Myh6 promoter [28] and generated stable CGR8 ES cell lines. Myh6-Wnt11 ES cells were also engineered with the nuclear DsRed fluorescent protein gene (DsRed-Nuc) under the same Myh6 promoter. The results show that Wnt11 increases Myh6 gene expression by 23-fold compared with 33-fold by Dkk1 treatment alone. Combinatorial treatment of Myh6-Wnt11 ES cells with Dkk1 throughout differentiation augments Myh6 expression levels by 95-fold (Fig. 7B). The increase in Myh6 gene expression corresponds to higher yields of red-fluorescence-labeled cardiac cells (Fig. 7C). These results suggest that combinatorial hyperactivation of canonical wnt signaling and enhancement of cardiogenic mesoderm can be further stimulated by Wnt11 to enrich cultures of contracting cardiomyocytes during ES cell differentiation.
Discussion
Pluripotent ES cells offer a practically unlimited source of cardiomyocytes to overcome some of the experimental obstacles associated with adult cells [4 –7,25]. The possibility to generate ES-like induced pluripotent stem (iPS) cells from adult somatic tissues opens further opportunities to isolate, analyze, and genetically modify cardiomyocytes from human patients [32 –34]. However, these approaches necessitate the ability to obtain sufficient numbers of cardiac cells for both experimental analyses and clinical applications [9,35,36]. Methods to achieve this goal have been developed and depend on timely manipulation of signaling pathways, favorable extracellular matrix conditions, hypoxia, or management of EB size [37 –39].
However, it becomes increasingly evident that individual ES cell lines have variable differentiation capacities in culture or show a wide range of differentiation timetables [34,40]. Therefore, lineage-specific differentiation regimes that depend on precise manipulation of particular steps would need to be customized for individual cell lines based on landmark differentiation points. A case in point is the contribution of the wnt signaling pathway in cardiomyocytic differentiation. Inactivation of various wnt morphogens or pathway modulators shows that they regulate multiple stages of cardiogenesis [41 –44]. Studies in zebrafish suggested a biphasic requirement that was also seen in differentiating ES cells [16]. However, it appears that the role of various wnt signaling branches is more complex and dynamic. For example, it was shown that canonical Wnt signaling, mediated by Wnt3a, is required for formation of endodermal and mesodermal cells [45,46]. After this step, canonical Wnt/β-catenin needs to be tempered by specific wnt ligand antagonists such as sFRP-1 and sFRP-5 in order for cardiac specification to proceed [47]. Noncanonical signaling initiated by Wnt5a and Wnt11 promotes this stage. Once CPCs become specified, then canonical wnt/β-catenin signaling driven by Wnt2b is again required to stimulate proliferation and expansion of CPCs [18,19]. Finally, a blockade of canonical and concomitant activation of noncanonical wnt signaling mainly by Wnt11 pushes differentiation of CPCs to cardiomyocytes [15,16]. During embryonic development and ES cell differentiation, these events take place within a narrow time window, thus making it difficult to time with precision proper manipulation of wnt signaling at corresponding peaks and valleys of activity.
Our data show that changes in wnt signaling take place immediately, at the onset of ES cell differentiation, starting with a downregulation of canonical signaling and a surge in JNK/AP-1 activity which lasts around 48 h in CGR8 cells. After this stage, there is a decrease in JNK/AP-1 activity followed by a sharp increase in canonical activity that is also maintained for approximately 48 h. The wnt signaling waves appear to mirror each other with canonical wnt activity maxima corresponding to minima of noncanonical activity and vice versa. The preceding noncanonical activity is necessary for the subsequent induction of canonical activity, as blocking JNK/AP-1 attenuates the ensuing canonical signaling.
The sequential arrangement of noncanonical/canonical signaling is required for proper differentiation of ES cells. Although interference with either branch at its maximum activity time window does not abolish ES cell differentiation, it results in opposite effects on lineage development. Specifically, JNK/AP-1 blockade reduces cardiogenic and endothelial differentiation with a concurrent increase in hematopoiesis and endoderm differentiation. In contrast, interference with canonical signaling with Dkk1 suppresses hematopoietic and endodermal lineages and induces endothelial and cardiomyocyte-specific gene markers. The last outcome, although consistent with our results, is in contrast to results obtained in another ES cell line showing that continuous application of Dkk1 blocks cardiogenesis [45]. It is likely that individual ES cell lines respond differently to various manipulations of specific signaling cascades. Consistent with this possibility, besides reacting in distinct ways to Dkk1 treatment, the 2 ES cell lines also respond differently to the BMP antagonist Noggin [45,48,49].
At the molecular level, JNK/AP-1 blockade moderates, but does not abolish canonical wnt signaling during mesoderm differentiation. Conversely, Dkk1 abolishes canonical wnt activity during the original mesoderm differentiation stages, but leads to increased canonical signaling during subsequent steps. These changes in canonical wnt activity lead to corresponding variations in T-brachyury expression as well as other markers that are involved in the generation of cells representative of the 3 embryonic germ layers. It has been postulated that 2 waves of T-brachyury expressing cells arise during ES cell differentiation, the first giving rise to blood and endothelial cells and the second to cardiomyocytes, smooth muscle, and endothelial cells. Therefore, it is likely that the delayed activation of canonical wnt signaling favors the expansion of the second wave of cardiogenic T-brachyury cells at the expense of the first.
Our results also show that both branches of wnt signaling are wired to counteract inhibition, as both JNK/AP-1 and canonical/β-catenin inhibition lead to subsequent activation of compensatory mechanisms that restore JNK/AP-1 and canonical/β-catenin signaling. Therefore, depending on their timing and duration, inhibitory signals might lead to apparently conflicting data regarding the effects of wnt signaling on ES cell differentiation to cardiovascular lineages. In this light, it appears that a critical determining factor is the time window of canonical wnt activity that coincides with T-brachyury induction and other gastrulation stage genes. Since different ES cell lines display slower or faster paces of differentiation with variable T-brachyury expression, it is likely that similar timings of treatment regimes could lead to opposite outcomes in individual cell lines. As a result, it is possible that both inhibition and activation of canonical wnt signaling have eventually the same outcome.
These results might be relevant for the adult situation as well where both inhibition and stimulation of canonical wnt signaling have been shown to have beneficial effects [36,50 –55]. It is possible that blocking wnt signaling in adult cardiac cells also triggers regulatory loops that lead to the opposite of predicted effects [56].
In conclusion, our results suggest an interconnection between canonical and noncanonical pathway branches, the existence of strong regulatory loops that counteract interference leading to unexpected outcomes in signaling directions, and that continuous antagonism of canonical wnt signaling creates a counter reaction increasing the number of cardiogenic progenitor cells and the expression of wnt ligands such as Wnt11, which favor cardiogenesis. Combinatorial Dkk1/Wnt11 application further enhances cardiogenic output. These findings could enhance future exploitation of ES cells and iPS cells to study the molecular and cellular bases of cardiovascular diseases and to develop strategies for myocardial regeneration.
Footnotes
Acknowledgments
The authors would like to thank the Vanderbilt FACS Core for expert assistance in cell analysis. This work was supported by NIH grants HL083958 and HL100398 to A.K.H., and Institutional Support from Vanderbilt Medical Center.
Author Disclosure Statement
No competing financial interests exist
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.