
Abstract
Ischemic or mechanical injury to the optic nerve is an irreversible cause of vision loss, associated with limited regeneration and poor response to neuroprotective agents. The aim of this study was to assess the capacity of adult bone marrow cells to participate in retinal regeneration following the induction of anterior ischemic optic neuropathy (AION) and optic nerve crush (ONC) in a rodent model. The small-sized subset of cells isolated by elutriation and lineage depletion (Fr25lin−) was found to be negative for the neuroglial markers nestin and glial fibrillary acidic protein (GFAP). Syngeneic donor cells, identified by genomic marker in sex-mismatched transplants and green fluorescent protein, incorporated into the injured retina (AION and ONC) at a frequency of 0.35%–0.45% after intravenous infusion and 1.8%–2% after intravitreous implantation. Perivascular cells with astrocytic morphology expressing GFAP and vimentin were of the predominant lineage that engrafted after AION injury; 10%–18% of the donor cells incorporated in the retinal ganglion cell layer and expressed NeuN, Thy-1, neurofilament, and beta-tubulin III. The Fr25lin− cells displayed an excellent capacity to migrate to sites of tissue disruption and developed coordinated site-specific morphological and phenotypic neural and glial markers. In addition to cellular reconstitution of the injured retinal layers, these cells contributed to endothelial revascularization and apparently supported remodeling by secretion of insulin-like growth factor-1. These results suggest that elutriated autologous adult bone marrow-derived stem cells may serve as an accessible source for cellular reconstitution of the retina following injury.
Introduction
I
To overcome this problem, researchers are directing attention to stem cell therapy. Neuronal and retinal progenitors, induced pluripotent cells, and embryonic stem cells have been shown to incorporate into injured retina at variable frequencies [12,13]. The bone marrow is a potential accessible source of autologous cells for retinal cell therapy, without the disadvantage of the immune barrier inherent to allogeneic cell transplants. Studies have suggested that committed retinal progenitors residing in the bone marrow compartment can be mobilized to contribute to retinal repair [14,15], particularly endothelial precursors and hemangioblasts that support angiogenesis and neovascularization [16 –18]. Accordingly, murine SCA-1+lin−c-kit+ and human CD133+ hematopoietic precursors were able to regenerate retinal pigment epithelium and support revascularization [19 –22] for extended periods [23]. In all instances, acute injury was an essential prerequisite for cell incorporation in the retina [19 –23]. In proof-of-concept studies of the capacity of adult bone marrow cells to adopt neural and glial phenotypes in vivo [24 –27], researchers reported a distinct mechanism of cell contribution to tissue repair in addition to support of endogenous recovery from injury [16,28]. These data are consistent with incidental findings of neurons, astrocytes, and microglia of donor origin in the brain of recipients of sex-mismatched bone marrow transplants [29]. However, efforts to induce such differentiation in experimental models and in vitro have been largely inconclusive [30,31].
Bone marrow cells isolated by counterflow centrifugal elutriation and depleted of hematopoietic lineage markers have limited radioprotective activity but are endowed with indefinite hematopoietic reconstituting potential [32,33]. The cells differentiate into hepatocytes in vitro and in vivo [34] and into endocrine pancreas and endothelial cells in vivo [35]. In addition, single cells are able to regenerate multiple epithelial tissues [36]. The aim of the present study was to assess the capacity of this subset of bone marrow cells to participate in neogenesis of neural and glial lineages in murine models after induction of ischemic and mechanical retinal injury. These models are particularly suited to evaluation of cell therapies, because the injured retina can be uninterruptedly accessed through small and medium-sized blood vessels [8 –11].
Materials and Methods
Animal preparation and transplantation
Two mouse strains were purchased from Jackson Laboratories (Bar Harbor, ME): C57BL/6J (B6, CD45.2), B6.SJL-Ptprca Pepcb/BoyJ (CD45.1), and C57BL/6-TgN(ACTbEGFP)1Osb [green fluorescent protein (GFP), CD45.2]. The mice were maintained and handled in accordance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research and the NIH guidelines, and the protocols were approved by the institutional Animal Care Committee.
Injury models
The models were based on a series of preliminary studies intended to optimize several variables in the experimental approach. In all experiments, injury was induced in 1 eye with the contralateral eye serving as a control. The induction of AION was performed as previously described [2]. Fifty-three mice were anesthetized with ketamine 80 mg/kg (Fort Dodge Laboratories, Fort Dodge, IA) and xylazine 4 mg/kg (Veterinary Medicines Directorate, Surrey, UK). A plastic custom-designed fundus corneal contact lens was used for direct in vivo visualization of the retina and optic nerve head. AION was induced by intravenous injection of 40 μL of 2.5 mM Rose Bengal dye (Sigma, St. Louis, MO) followed by selective photoactivation above the optic disk at 532 nm with an argon laser beam at the following specifications: 200 μm spot size, 50 mW power, and constant duration (0.1 s) [2]. In all mice, the right eye was injured and the fellow (healthy) eye served as an internal control. The injury was re-evaluated by RGC count in tissue sections; a 35%–40% RGC loss was previously reported for this reproducible procedure [2,8]. ONC was induced by 3 repeated applications of a forceps for 7 s at ∼3.0 mm posterior to the eye globe [3]. Studies have shown that crush of the optic nerve leads to ∼70% RGC loss [3].
Isolation of bone marrow cells
Nucleated bone marrow cells were harvested from C57BL/6 mice constitutively expressing GFP. Femurs and tibia were loaded onto a Beckman counterflow elutriation centrifuge (Fullerton, CA;
Stem cell administration
The elutriated Fr25lin− subset harvested from the donor mice was transplanted into the congenic C57BL/6 recipients using 2 routes of administration: 106 cells in 0.2 mL PBS infused intravenously and 2×105 cells in 3 μL PBS inoculated intravitreally (Table 1). Treatment was administered 24 h after injury induction on the basis of preliminary findings of better cell incorporation with this schedule than with treatment before or immediately after injury (Fig. 1A). Conditioning was eliminated for all transplantations, because our preliminary trials showed that sublethal irradiation (650 rad, 106 rad/min) using an X-ray irradiator (RadSource 2000, Suwanee, GA;
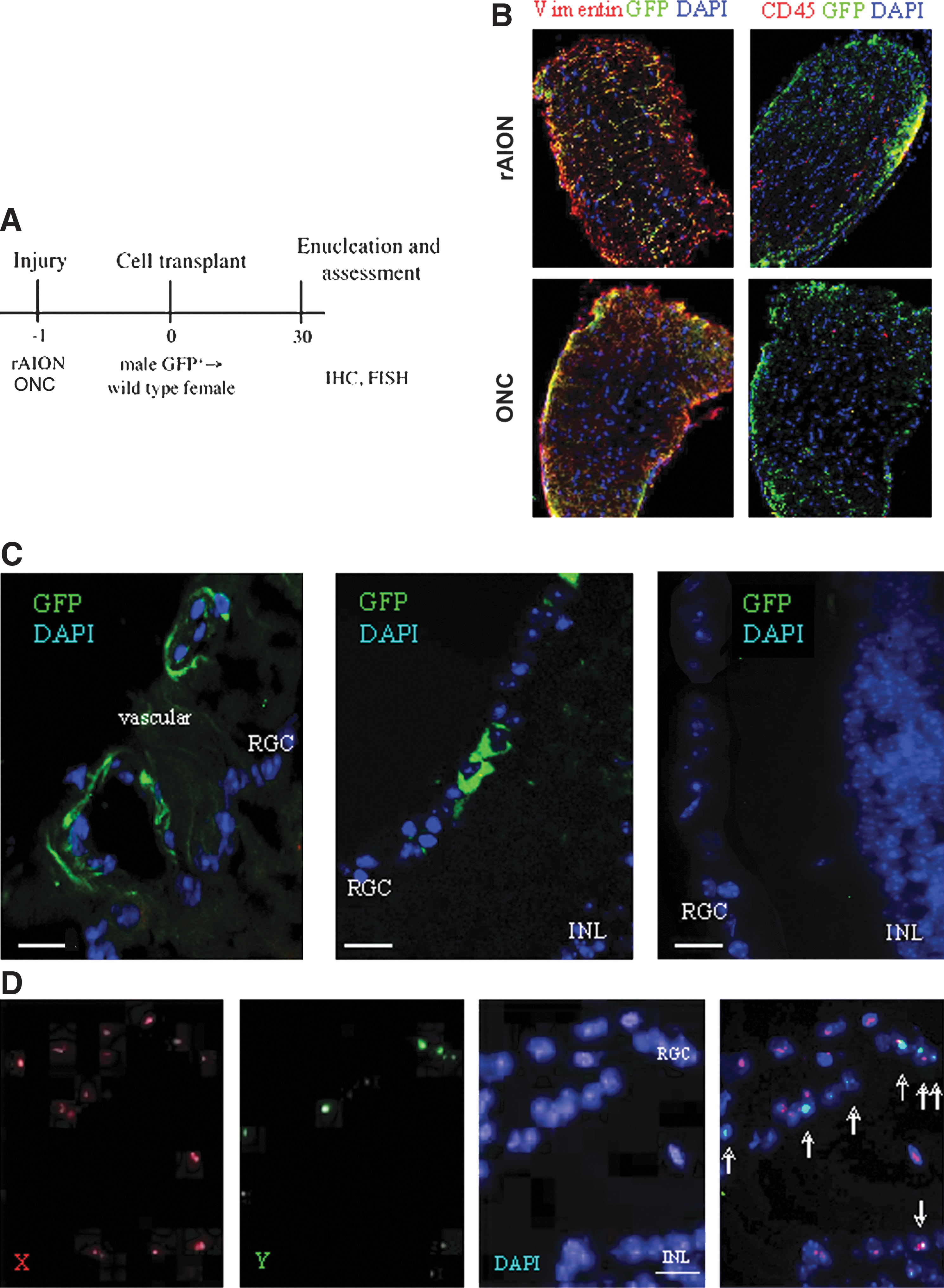
Incorporation of donor cells in the injured optic nerve and retina.
AION, anterior ischemic optic neuropathy; ONC, optic nerve crush; Fr25Lin−, fraction 25 lineage-negative; R/O, rotor off; IV, intravenous; IVT, intravitreal.
Histology and immunohistochemistry
Mice were sacrificed at 30 days after transplantation, and the tissues were analyzed by histology and immunohistochemistry. The eye bulb was enucleated, fixed in 4% paraformaldehyde overnight, and then transferred to 20% sucrose overnight at 4°C. Samples embedded in frozen OCT medium (Tissue-Tek, Sakura, Finerek, CA;
Fluorescence in situ hybridization
Grafted cell incorporation into the vascular and RGC layers was validated by fluorescence in situ hybridization (FISH) in male-to-female transplants [38]. In brief, FISH was performed in sections washed in 0.09% saline and 2×saline sodium citrate buffer (SSC) and digested by 0.5 mg/mL proteinase K. Sections were dehydrated at ascending ethanol concentrations (70%–100%) and air-dried. Thereafter, 10 μL of the probe solution labeled with PlatinumBright 495 and 550 (Poseidon FISH Probes, Amsterdam, The Netherlands;
Microscopy
Images were acquired with an Axioplan 2 fluorescence microscope (C. Zeiss, Göttingen, Germany;
Quantitative assessment of cell incorporation in the injured retina
To estimate the number of labeled cells in each eye, 6–15 nonoverlapping fields of view were assessed in every 10th slide, with each slide including 3 consecutive cryosections. The incorporation of transplanted cells was calculated by dividing the number of donor cells (GFP+) by the number of cells staining positive for tissue markers in the relevant layer. The total number of donor cells was calculated by multiplying the number of cells per section by the number of slides (10) and correcting for the spherical shape of the retina: Πr 2/4r 2=Π/4. The homing efficiency (in percent) was calculated by dividing the total number of GFP+ cells by the number of infused cells.
Polymerase chain reaction
Real-time quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was used to study the expressions of nestin, GFAP, insulin-like growth factor-1 (IGF-1), fibroblast growth factor-2 (FGF-2), transforming growth factor-β (TGF-β), and tumor necrosis factor-α (TNF-α). Elutriated fractions were frozen in liquid nitrogen. Total RNA was extracted with TRIzol (Invitrogen), followed by reverse transcription into cDNA using random hexamers (Amersham Biosciences, Cardiff, UK;
Statistics
All data are expressed as mean±standard error. The results were statistically analyzed with SPSS for Windows, version 15.0.1 (SPSS, Inc., Chicago, IL). Two-way ANOVA with repeated measures was used to analyze within subject factor type (route of injection, differentiation). A P value of less than 0.05 was considered significant. Because of the relatively small size of some groups, we applied the nonparametric Mann–Whitney test for 2 independent samples.
Results
Incorporation of donor cells in injured retina
Without cell transplantation, both AION and ONC led to cell loss in the retinal ganglion layer, of approximately 30% and 70%, respectively (Fig. 1).
Both AION and ONC caused massive gliosis but only modest inflammation (Fig. 1B). FISH study validated the predominant incorporation of the elutriated Fr25lin− subset into the anterior vascular and RGC layers in the male-to-female transplants (Fig. 1D). In the particular area of the retina presented in Fig. 1D, the X chromosome was detected in 85% of the RGC nuclei; 6 cells (30%) displayed the male X-Y genotype, in addition to cells incorporated in the inner nuclear layer (GFP+ cells; Fig. 1C, left and middle). The control eyes showed virtually no cell incorporation in the absence of injury (Fig. 1C, right).
Differentiation of Fr25lin− cells in the injured retina
The subset of elutriated Fr25lin− cells used in this study expressed the panhematopoietic marker CD45 at variable levels; however, the majority of cells that incorporated in the RGC layer were CD45 negative (Figs. 2A and Table 1). On lineage analysis, some of the GFP+ donor cells located in the vasculature anterior to the RGC layer had an endothelial morphology and expressed CD31, whereas other cells cuffed the blood vessels and expressed vimentin (Fig. 2B) or GFAP (Fig. 2C).
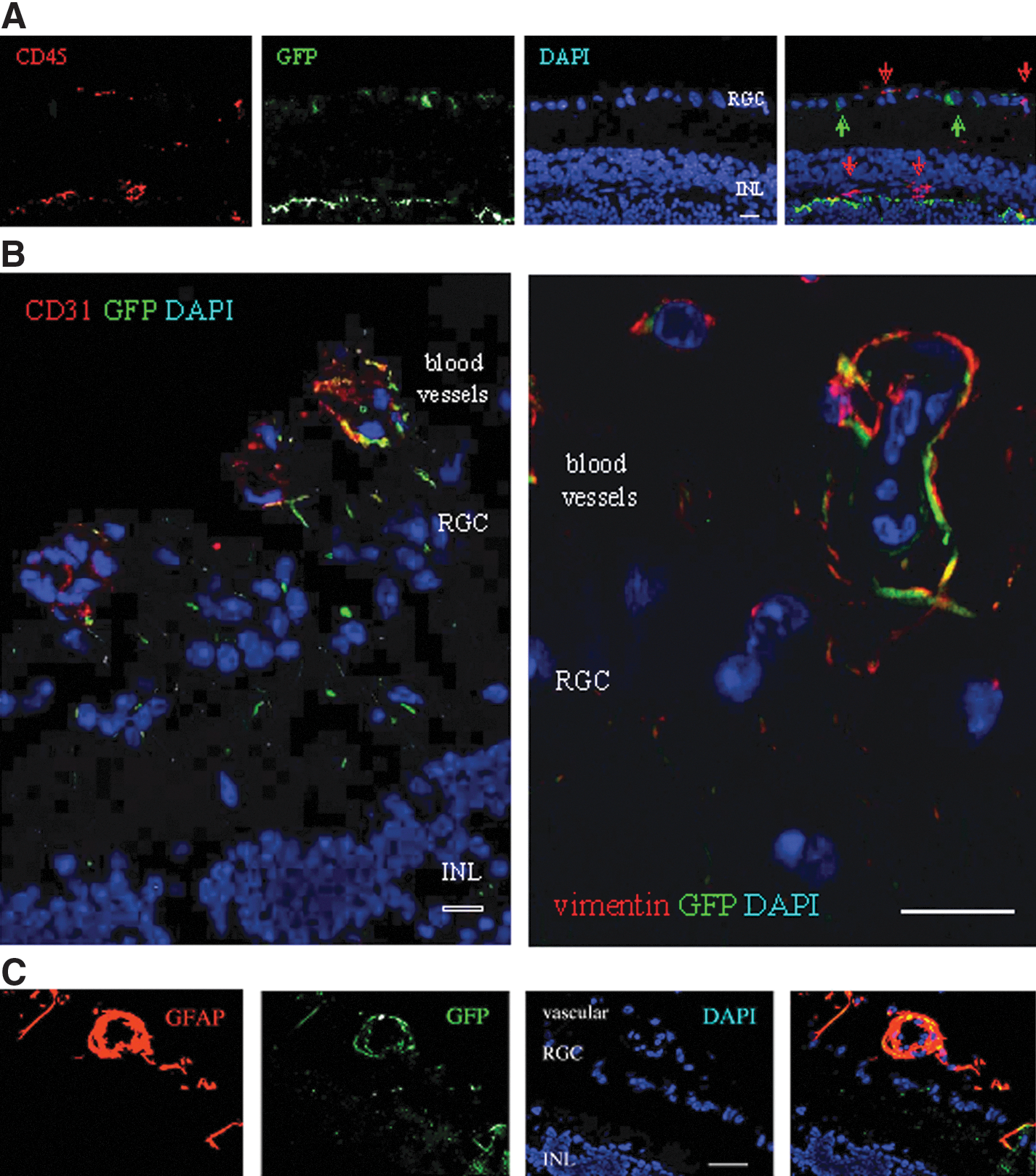
Immune, vascular, and glial differentiation.
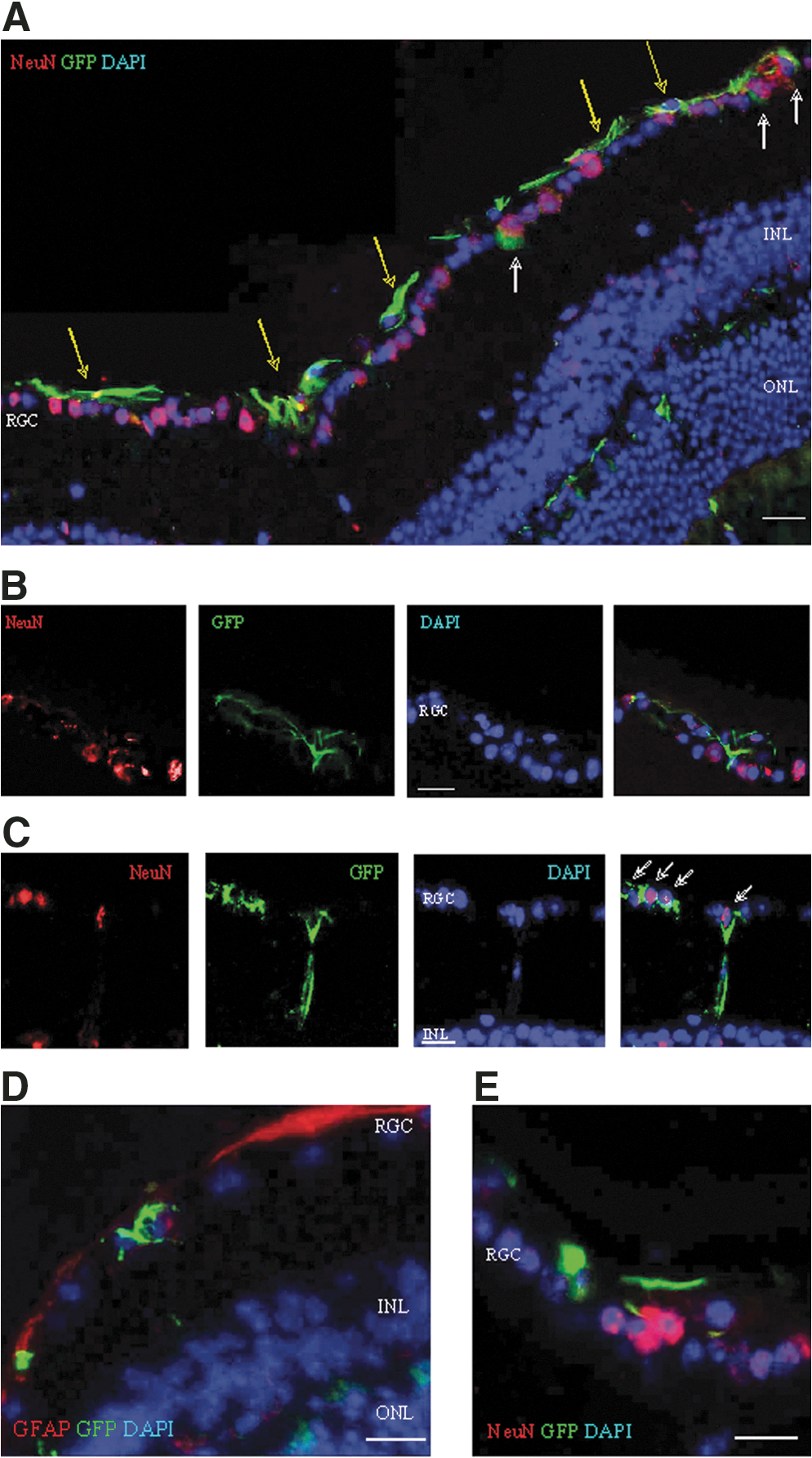
Because astrocytic processes surround ganglion cells, the morphologic differentiation of the donor cells to astrocytes made it difficult to positively identify the cells engrafted in the RGC layer. Microscopic analysis at high magnification dissociated the cells coexpressing cytosolic GFP and vimentin from GFP+ cells in the RGC, which displayed a different morphology and lacked astrocytic markers (white arrows in Fig. 3A). Superposition of pseudocolored fluorescence layers of green GFP and red vimentin colored the cytosol yellow (yellow arrows in Fig, 3A). Reciprocally, the GFP+ cells with the characteristic astrocytic morphology in the anterior vasculature were negative for NeuN (Fig. 3B); however, NeuN was detected in the nuclei of GFP+ cells incorporated in the RGC layer (Fig. 3C). The capacity of RGC-engrafted donor cells to upregulate neuronal markers was confirmed by the expression of nuclear Thy-1 (Fig. 3D) and cytosolic beta-tubulin III (Fig. 3E) and neurofilament (Fig. 3F).

Expression of neuronal markers in cells incorporated in the RGC layer.
Effect of route of administration on quantitative cell incorporation in the injured retina
Quantification of the cell engraftment in the injured retina at 1 month after transplantation revealed significant differences by route of administration (Table 2). In both models of injury, engraftment was consistently better when the cells were inoculated into the vitreous body (∼1.9%) rather than infused intravenously (∼0.4%) (Table 2). In both models, engraftment was restricted to the injured retina.
Values are mean±SD.
Effect of nature of injury on donor cell differentiation
A similar quantitative cell engraftment suggested that the rate-limiting step of incorporation in the retina is the number of Fr25lin− cells that are responsive to the injury signals (Fig. 4). Cells inoculated into the vitreous body showed a clear tendency to concentrate along the internal layers of the injured retina (Fig. 4B). Following incorporation in the retina, the Fr25lin− cells adopted layer-specific phenotypes at different frequencies (Fig. 4A): predominantly glial differentiation under ischemic conditions (AION, Fig. 4D) as opposed to more pronounced expression of the neuronal markers in the ONC model (Fig. 4C). In addition, mechanical injury (ONC) enhanced the differentiation of the GFP+ donor cells to CD45+ immunohematopoietic phenotypes (Fig. 4E).
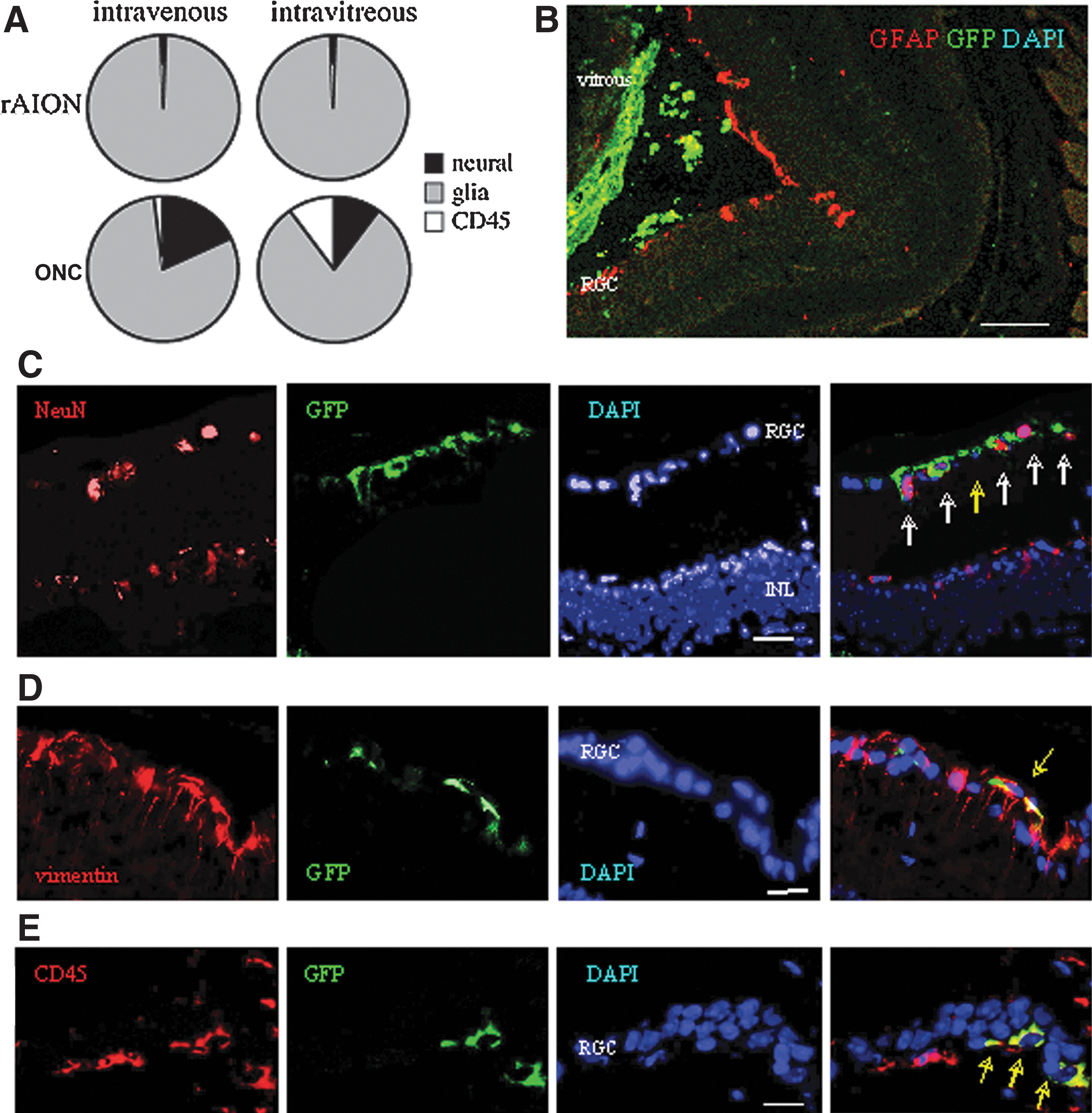
ONC injury enhances neural and immune differentiation.
Absence of neuroglial progenitors in grafted Fr25lin− cells
Earlier findings of the differential incorporation of Fr25lin− cells under various experimental conditions raised the possibility that bone marrow does not contain committed neuroglial precursors that respond to particular injury signals [14,15] (Fig. 5). To determine whether the Fr25lin− subset was primed to neuroglial lineages, we measured mRNA encoding GFAP and nestin. The results showed that the expression of both markers was low in whole bone marrow cells and decreased further by elutriation and lineage depletion (Fig. 5A).
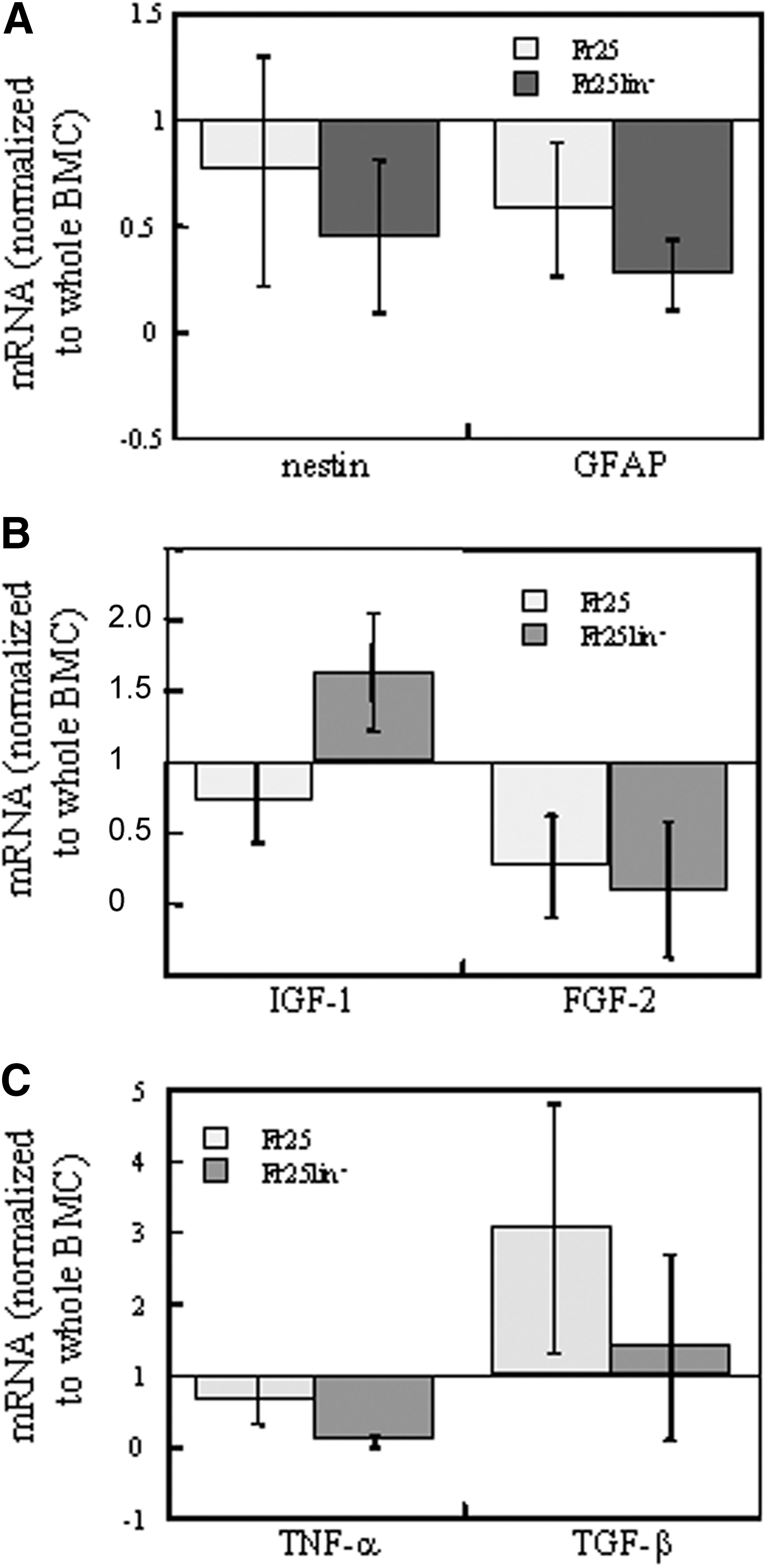
Expression of lineage markers and chemokines in grafted cells. Real-time polymerase chain reaction measurements in elutriated (Fr25) and lineage-depleted fractions (Fr25lin−) are normalized against mRNA expression in whole bone marrow cells. Data represent 3 independent mRNA extracts:
Supportive effects of the grafted cells
In addition to their neural and glial neogenesis ability, donor Fr25lin− cells may also support endogenous cell reconstitution through modulation of the microenvironment in the injured retina (Figs. 5 and 6). Our analysis of several factors that may play an active role [40] yielded an enhanced expression of IGF-1 by the donor cells (Fig. 5B). The Fr25lin− donor cells did not participate in retinal remodeling through secretion of FGF-2, TNF-α, or TGF-β (Fig. 5B, C).
Cell-to-cell interactions.
The inability to perform a functional assessment of the impact of grafted cells is an intrinsic drawback of experimental retinal injury. However, we found indirect evidence of donor cell participation in postinjury stabilization of the retinal architecture. (1) There was a strong correlation between the lineage-selective morphology of the differentiated donor cells and the corresponding phenotypic markers specific to the layer in which they engraft. (2) The differentiated donor cells filled gaps and contributed to structural confluence (Fig. 6A, C). In particular, cells located at sites of vascular disruption expressed astrocytic markers (Fig. 6D) and extended processes to the ganglion cell (Fig. 6C, E). (3) In addition to the astrocytes of GFP+ donor origin that support the RGC layer (Fig. 6B), the donor GFP+NeuN+ cells that were aligned with ganglion cells displayed a pyramidal shape characteristic of young neurons and extended dendritic processes toward bipolar cells and the inner nuclear layer (Fig. 6C).
Discussion
The present study was performed in murine models of ischemic or mechanical retinal injury using small-sized stem cells derived from adult bone marrow. The Fr25lin− cells displayed an excellent capacity to detect the sites of injury in the retina and to assume the corresponding morphology and phenotype. The different patterns of differentiation of the donor cells in the retina corresponded to the specific structures in which the cells were located. Such fine-tuned patterns of integration reflect the high responsiveness of the primitive Fr25lin− cells to diverse signals that direct their migration, engraftment, and differentiation.
Quantitatively, the route of administration was the primary factor affecting engraftment. Previous studies have shown that the majority of intravenously injected bone marrow cells are filtered in the liver and lungs and only ∼10% of the progenitors home to their physiological site of residence [41,42]. Accordingly, in our preliminary study, irradiation decreased quantitative cell engraftment in the retina, presumably by facilitating uptake by the bone marrow, which competes with the injured tissue for primitive cells. We found that intravitreal inoculation resulted in better incorporation of the donor cells (∼2%) than intravenous infusion (∼0.4%), suggesting that the inoculated cells responded better to recruitment signals. At the same time, the high acuity of donor cell retention and engraftment at sites of disrupted tissue structure in both experimental models, even in the presence of the injury-induced disruption to the blood–brain barrier [43], points to a specific chemotactic response also of the cells in the peripheral circulation.
Injury is absolutely essential to elicit migration to and incorporation in the retina. In our study, the grafted cells were virtually absent in the unaffected (contralateral) eye. This has been reported for other types of bone marrow-derived cells and neural progenitors as well [19 –21,31,44 –46]. Further, the nature of the injury affected the location of cell engraftment and the phenotype, thereby dictating the fate of the differentiating cells.
The AION model causes selective damage to the RGC layer, the most internal layer of the retina. The other layers of the sensory retina, as well as the external nonsensory RPE layer, which supports the photoreceptors, remain intact. The dominant astrocyte differentiation of the donor cells at sites of vascular deficiency in the internal retinal layer was common to both models, but the differentiation of the nonglial (neural and inflammatory) lineages was more prevalent following mechanical injury. These findings are supported by an earlier study wherein primary neuronal damage caused by ONC led to retrograde degeneration and necrotic or apoptotic death [3] and substantially promoted neuronal differentiation in 10%–18% of the engrafted donor cells. In contrast, ischemia led to predominantly glial neogenesis. Others observed that in both ischemic and mechanical injury models, disruption of the blood–brain barrier abolished the eye's immune privilege [47], but only mechanical injury was favorable to induction of CD45 immunohematopoietic cells. The marked variability in morphology and phenotype in the present study, together with the absence of hematopoietic cell engraftment in the absence of conditioning, suggest that the engrafted donor cells adopted a lineage-specific differentiation in response to local cues in situ. Even if a few progenitors reach the bone marrow, functional engraftment of Fr25lin− cells usually appears later than 1 month post-transplantation [32,33,37], when the present analysis was performed.
In a recent study in a retinal laser burn model, Singh et al. [48] delivered Lin-negative bone marrow cells intravenously and intravitreously. The intravenous route was found to be superior in terms of cell mobilization. However, their study differed from ours in several important aspects. (1) Singh et al. [48] used a whole bone marrow cell lineage marked with chemical labeling (carboxyfluorescein diacetate succinimydal ester), whereas we used elutriated small fraction (Fr25) lineage-depleted cells of bone marrow extracted from transgenic GFP+ mice. (2) Singh et al. [48] applied a laser injury (burn) to the retina, which destroyed both the photoreceptors and the RPE, the most external layers of the retina. In contrast, we selectively damaged the RGC by inducing AION or ONC, which affected only the internal neuronal layer. The near location of the injured layer and vascular choroids relative to the distance of these layers from the vitreous may explain the better transplantation outcome for the intravenous route in the study by Singh et al. [48]. (3) Singh et al. [48] injected the cells immediately after induction of injury, whereas we injected them 1 day after injury, during the maximal point of damage (based on our preliminary data). (4) Singh et al. [48] injected significantly fewer and less selective cells, 50,000 and 100,000 cells intravitreously, with no incorporation at the higher dose because of clumping. Intravenously, they injected 100,000 cells compared with 1×106 in our study, with better results. This finding may be related to the different types of injury. Overall, the different layers of the eye injured may account for the discrepant results.
The bone marrow is a heterogeneous compartment that hosts multiple cell types and precursors. The manner in which it contributes to revascularization is still controversial. Some researchers attributed this contribution to the hemangioblasts and endothelial progenitors committed to angiogenic differentiation that reside in the bone marrow [16 –21,49,50]. This led to the assumption that committed retinal progenitors from the marrow compartment can be recruited to replenish the retinal pigment epithelium layer [14,15]. Alternatively, the adult bone marrow may contain subsets of pluripotent cells that can respond to multiple signals and adopt versatile differentiation traits. In the present study, in line with previous reports that elutriated Fr25 cells do not express the hemangioblast and hematopoietic stem cell markers CD31, CD34, SCA-1, and c-kit [51 –53], we found no evidence of nestin or GFAP expression in these cells. Thus, it is unlikely that vascular, glial, and neural conversion can be attributed to tissue progenitors sequestered in the bone marrow, even if the commitment of precursors is limited to priming at the transcriptional level [54,55]. Nestin is a promiscuous marker of ectodermal and gastrointestinal precursors and mature lineages [56,57]. However, it frequently accompanies GFAP, an early marker of dual neural and glial differentiation of astrocytes and neural progenitors [58,59], during astrocyte differentiation [60]. Further, we recently reported that endocrine differentiation of Fr25lin− cells is not restricted to the expression of progenitor markers SCA-1, c-kit, and Thy1 [38]. This finding is consistent with the minor advantage of prospectively isolating SCA-1+ and Thy-1+ in bone marrow cells for the in vitro generation of nestin-positive neurospheres [61]. Although the neuroglial and hematopoietic markers are considered to be involved in neural differentiation of mesenchymal stroma cells [62,63], it remains to be determined whether the small-sized subset of elutriated cells contains sufficient numbers of mesenchymal stromal cells to account for the observed engraftment in the injured retina. Nevertheless, evidence points to a differential capacity of various bone marrow-derived cell types to repair the retinal pigment epithelium, reduce photoreceptor degeneration, induce angiogenesis under various experimental conditions, and mediate choroidal revascularization following laser-induced injury affecting the outer retinal layers and Bruch's membrane [14 –22,31,46,49,50].
On the basis of the present findings, we suggest that a putative subset of pluripotent cells, such as the one used in this study, migrates to various layers of the retina and, in response to specific local signals of injury, differentiates into endothelium (CD31), immune cells (CD45), astrocytes (GFAP or vimentin), or neural cells (NeuN, Thy-1, beta-tubulin III). This interpretation complements earlier findings that these cells regenerate the endocrine pancreas, liver, and multiple epithelial tissues [34 –36] and that the various subsets of adult bone marrow cells adopt neural and glial phenotypes in vivo [24 –27]. However, there is no evidence that tissue-committed progenitors residing in the bone marrow navigate to specific sites and develop according to an imprinted (genetic and epigenetic) program. True differentiation is exemplified by the CD45-negative phenotype of the retina-engrafted cells and the normal diploid genotype. Rare events of fusion between stem cells or the myeloid progeny of hematopoietic progenitors and injured tissue might result in lateral transfer of specific markers without true differentiation [64]. An overzealous speculation that fusion in culture negates developmental plasticity [65] of bone marrow cells that differentiate into glia and neurons [24 –27] is not substantiated by careful genotypic examination of diploid phenotypes of neural progenitors converting into endothelium [66] and of Fr25lin− cells converting into hepatocytes [34] and insulin-producing cells [35]. The capacity of myeloid lineages to fuse, phagocytose, and engulf apoptotic and necrotic cells is irrelevant to the observed phenomena in the absence of CD45 expression in the hybrids [67]. The data reported here are likely to be an underestimate of cell engraftment in the retina [38], considering that only ∼50% of grafted cells express GFP [68] and the expression of this reporter protein is markedly downregulated during neuronal differentiation [69].
Locating donor cells, especially long after transplantation, is technically difficult. Although the retina is considered an immunologically privileged site [70], researchers have reported GFP rejection in rabbits early after viral transfection of retinal cells [71] or after subretinal administration of lentiviral vectors [72]. The rejection was a humoral immune response and not a cellular reaction [73,74]. There was no rejection when the vector was injected without any transgene, indicating that it was not due to the viral vector [73]. In the present study, the donor cells were derived from GFP-transgeneic mice and were not virally transfected. We did not find signs of GFP rejection in the eye, in accordance with numerous other investigators who used GFP cells from transgenic GFP donors [74 –77]. The cells were injected either intravenously or intravitreously, but not into the subretinal space, and they survived the circulation and reached the injured retina to incorporate into it and differentiate. The lack of rejection may be explained by species differences, the small number of cells injected, or the weaker expression of the transgenic proteins. To overcome the labeling difficulties in the above techniques, we also identified donor cells by other methods: 42.1 and 42.2 and FISH for X-Y mismatch.
One of the major drawbacks of neural models of regeneration is the difficult functional assessment of the organ. Unlike models of diabetes and hepatotoxicity [34,35], we do not know to what extent the observed contribution of the grafted cells improves organ function. Preliminary studies indicated better axonal transport following transplantation, but it is unlikely that axonal processes of donor cells adopting neural phenotypes in the RGC were responsible for such distant transport. Rather, it may primarily rely on collateral support of endogenous repair through vascularization and secretion of neuroprotective agents [40]. We assessed the possibility that the grafted cells further contribute to tissue remodeling after injury by local secretion of neuroprotective agents. We detected increased levels of mRNA encoding IGF-1 [78,79], but no increased expression of TGF-β [80,81], FGF-2 [82 –84], or the neurotrophic factor TNF-α [85 –87].
In summary, a small-sized subset of adult bone marrow cells contributes to retinal remodeling after ischemic and mechanical injury by both direct contribution to cellular regeneration and support of endogenous repair. The major direct cellular contribution is gliosis, which is the dominant inductive process within the physiological mode of repair of the retina as a neural tissue. Precursors of the various lineages are not present in the grafted cells, which develop phenotypes corresponding to the site of engraftment. The correlation among stem cell location, morphology, phenotype, and cooperation with endogenous cells in stabilization of retinal architecture is strong evidence that those with flexible developmental potential receive site-specific inductive signals. A second mechanism is support of retinal remodeling through angiogenesis in the immunomodulation and secretion of IGF-1. Monitoring of retinal function will determine the significance and potential implementation of adult autologous bone marrow-derived cells in ischemic eye injury.
Footnotes
Acknowledgments
The authors thank Dr. Saul Sharkis and Dr. Michael Collector for the outstanding support, discussion, and conceptual contribution to this study. Funding was provided by generous grants from the Leah and Edward M. Frankel Trust for bone marrow transplantation, the Zanvyl and Isabelle Krieger Fund, Israel Academy of Sciences and Humanities (ISF, NGC, 1371/08), the Eldor-Metzner Clinician Scientist Award, Chief Scientist, Israel Ministry of Health (NGC, 3-3741), and the Lirot Foundation (NGC, 3-4538) and a pilot grant from the North American Neuro-Ophthalmology Society and by Tel Aviv University: The Adams Super Center for Brain Research (2004–2005, 2005–2007, NGC); the Clair and Ameddee Maratier Fund to prevent blindness (2004–2005; 2010–2011; NGC, 2008 BRAL); Walter Friendliest and Herman Shudder Fund (NGC 2006–2008); The Lions (BRAL 2010) Mazritzky Fund (TS 2010).
Author Disclosure Statement
The authors declare that no competing interests exist. The results were presented in part at the Association for Research in Vision and Ophthalmology, Fort Lauderdale, FL (ARVO), 2006, 2007; Meeting of the Israel Society for Eye and Vision Research, Neve Ilan, Israel (ISVER), 2006, 2007, 2008, 2009, 2010; Meeting of the North American Neuro-Ophthalmology Society (NANOS), Tucson, AZ, 2006, 2010; Meeting of the International Society for Stem Cell Research (ISSCR), Philadelphia, PA, 2008; the Second International Stem Cell Meeting, 2008, 2009, Tel Aviv, Israel (BIOMED); Tel Aviv Research Day, Tel Aviv, 2008, 2010; Rabin Medical Center Research Day 2006, Petach Tikva, Isreal; the Ninth Michaelson Symposium 2007, Baltimore, MD; Meeting of the Macular Society, 2007, London, United Kingdom; the Second Young Investigators Stem Cell Meeting 2009, Bar Ilan University, Ramat Gan, Israel; and ILANIT/FISEB Federation of the Israel Societies for Experimental Biology Meeting, Eilat, Isarel, February 2011.