
Abstract
Human embryonic stem cells (hESCs) are one of the most interesting cell types for tissue engineering and cell therapy. The large-scale banking of hESCs for research and future clinical application requires economic, defined, and xeno-free cryopreservation protocols. In this study, the possibility to substitute knockout serum replacement (KO-SR) in the cryopreservation process with vegetal and synthetic hydrolysates was investigated. To our knowledge, the use of hydrolysates in hESC cryopreservation has not been yet explored. Initially, 3 different hydrolysates (Ultrapep Soy, Hypep 4601 and EX-CELL® CD Hydrolysate Fusion) were tested in the cryopreservation solution. A concentration of 8 mg/mL EX-CELL CD Hydrolysate Fusion in the cryopreservation solution leads to the highest recovery ratio; thus, this solution was selected for additional cryopreservation experiments. To ensure reproducibility of the results, 3 hESC lines (HS181, H9, and BG01) were used. The hESCs were collected prefreezing by application of collagenase IV and cell dissociation solution. Experiments showed that it was feasible to substitute the KO-SR in both the cryopreservation solution as the thawing solution. The obtained recovery ratios were comparable to those obtained with KO-SR (no statistical significant difference; Student's t-test, P<0.05). Further optimization protocols showed a doubling of the obtained recovery ratio after addition of Rock-inhibitor Y-27632 post-thawing. The expansion profile and pluripotency analysis revealed no changes in normal hESC behavior. We conclude that the application of vegetal or synthetic hydrolysates is suitable for xeno-free hESC cryopreservation.
Introduction
H
Two techniques are available for hESC cryopreservation: vitrification and slow freezing–rapid thawing. Vitrification is a technically challenging technique, mostly performed with open pulled straws, in which a low amount of hESCs are frozen with a high amount of cryoprotectors in order to quickly obtain the solidified glass-like state [6,7] in which the hESCs are preserved. During the slow freezing–rapid thawing process, large amounts of hESCs can be frozen, with a lower amount of cryoprotectors, in closed cryovials. Regardless of the cryopreservation technique applied, the cryopreservation solutions often contain fetal bovine or calf serum (FBS/FCS) [8 –11], knockout serum replacement (KO-SR) [12 –15], or serum albumin [16]. Currently, FBS or FCS in standard hESC medium is generally replaced by KO-SR in order to obtain more defined culture conditions. However, KO-SR is still prone to batch-to-batch variations and contains Albumax, a lipid-rich bovine albumin fraction, and bovine transferrin. As it has been shown that animal proteins can transfer from KO-SR to the hESCs [17,18], their use in hESC culture and cryopreservation is far from ideal. Another strategy for therapeutic hESC cryopreservation is the use of commercially available cryoprotector solutions [5], such as CE-marked dimethyl sulfoxide (DMSO), and media such as STEM-CELLBANKER™ [19] and CRYOSTOR™ (Stemcell Technologies). However, the fact that not all substances in proprietary commercial solutions are disclosed poses risks regarding the application in a clinical setting.
As an alternative path for therapeutic hESC cryopreservation, we explored the possibility to use vegetal hydrolysates as additive in the cryopreservation process. The use of hydrolysates or peptones is not new in cell culture. Hydrolysates are commonly used in microbial culture media to support microbial, fungal, and bacterial growth, in industrial fermentations, and in human and veterinary vaccine production. Additionally, most of the current serum-free media contain some form of animal-derived peptones or hydrolysates [20]. In general, the hydrolysates act as a source of nutrients, being composed of amino acids, oligopeptides, lipids, and trace elements [21]. Animal, vegetal, and recently also synthetic hydrolysates are commercially available and can be easily produced in high quantities under quality-controlled conditions. Hydrolysates have been experimentally tested in in vitro porcine embryo developmental studies [22] for freezing of bovine embryos [23] and in embryo, keratinocyte, and stem cell culture media [20,24 –26]. To our knowledge, it has not been used in hESC cryopreservation. This suggests that it should be possible to obtain a cryopreservation method with hydrolysates.
In this study, we investigated the use of hydrolysates, from vegetal and synthetic origin, as a substitute for KO-SR in hESC cryopreservation. Our goal was to establish a xeno-free cryopreservation procedure, ideal for hESC banking.
Materials and Methods
Cell culture
Feeder preparation: mouse embryonic fibroblasts
Mouse embryonic fibroblasts (MEFs) originating from MF1 mouse embryos between passage 0 and 4 were used as a feeder layer to sustain the hESC culture. The MEFs were treated with 10 μg/mL mitomycin C (Sigma Aldrich) for 3 h to arrest mitosis and seeded at a density of 8,500 cells/cm2 on gelatin-coated tissue culture plates.
hESC standard culture protocol
The hESC lines used in these experiments were the H9 (Wicell), the BG01 (Wicell), and HS181 (Karolinska Institutet, Stockholm, Sweden). The hESCs were cultured on mitomycin-C–inactivated MEFs on tissue culture plates at 37°C and 5% CO2. The hESC medium consisted of 80% Dulbecco's modified Eagle's medium (DMEM)-F12 (Invitrogen)+20% serum replacement (Invitrogen)+1 mM
Cryopreservation and thawing protocols
hESC slow-freezing protocols
The cryopreservation–thawing procedures were performed on the H9, BG01, and HS181 hESC lines. For these experiments, passages 37–50 were used from the H9 cell line, passages 41–54 from the BG01 cell line, and passages 42–55 from the HS181 cell line. The hESC colonies were collected for slow-freezing procedures by detachment with collagenase IV and subsequent treatment with cell dissociation solution [27]. The cryopreservation solution consisted of 5% DMSO+5% hydroxyethylstarch (HES)+the experimental solution. Initially, hydrolysates of different origins were tested: EX-CELL® CD Hydrolysate Fusion (Sigma Aldrich), Ultrapep Soy (Kerry Bioscience), and Hypep 4601 from wheat gluten (Sigma Aldrich). Three different concentrations were examined: 8 mg/mL (medium A, D, and G), 6 mg/mL (medium B, E, and H), and 10 mg/mL (medium C, F, and I). Next, the best experimental condition was chosen for further tests. Three combined cryopreservation–thawing procedures were evaluated: 1. Cryosolution with HM and thawing with 0.1 M sucrose-HM 2. Cryosolution with Excell medium and thawing with 0.1 M sucrose-HM 3. Cryosolution with Excell medium and thawing with 0.1 M sucrose-Excell medium
HM medium consisted of 80% DMEM-F12 (Invitrogen)+20% serum replacement (Invitrogen)+400 μL HEPES. The Excell medium consisted of KO-DMEM supplemented with 8 mg/mL EX-CELL powder. After preparation, 1 mL of the cryosolution with cells was transferred to a cryovial (Cryo.S™; Greiner Bio-One), cooled slowly (cooling rate: −1.5°C/min) to −80°C, and then stored in a liquid LN2 storage device. All protocols were thawed in a water bath at 37°C, until an icy mass remained. Next, the thawing solution was added and left to incubate during 5 min. Then the cell suspension was centrifuged and plated out onto fresh MF1 feeders.
hESC slow-freezing protocols: further optimization
The best condition (Cryosolution with Excell medium and thawing with 0.1 M sucrose–Excell medium) was selected and explored for further optimization. A simplification of the detachment protocol and the addition of Rho kinase inhibitor (ROCK-inhibitor, 5 μm) Y-27632 during the first 24 h post-thawing was investigated. This led to the following cryopreservation protocol: 4. Condition 3, with direct Cell Dissociation treatment (20 min) 5. Condition 3, with 24-h Y-27632 (ROCK inhibitor) treatment 6. Condition 3, with direct Cell Dissociation treatment (20 min) and 24-h Y-27632 (ROCK inhibitor) treatment
Evaluation, data collection, and statistical analysis
All experiments were repeated 3 times in triplicate, and all vials were kept for minimally 1 week in LN2. The amount of hESC colonies, obtained from full-grown plates, was counted before freezing and at day 7 post-thawing. The hESC colonies were morphologically evaluated and counted using a gradual scoring system [14,28] on an inverted phase-contrast light microscope with objective 4× (Olympus inverted Research System Microscope IX81). Only grade A (>80% undifferentiated) and grade B (between 50% and 80% undifferentiated) colonies were selected for further use in the evaluation as these are the only colonies capable of generating new pluripotent colonies in the next passage. The recovery rate was calculated as follows: the amount of grade A+B colonies at day 7 post-thawing versus the amount of frozen grade A+B colonies. For each counted colony number, an uncertainty degree of 5% was taken into account. This degree of uncertainty was established after repeated counting experiments of the same plate and used to determine the error bars in the statistical graphical representations [29]. The Student's t-test was used for determining significant differences between the obtained amounts of colonies after freezing. P<0.05 was considered significant.
hESC postcryopreservation assessment
hESC: expansion profile
The thawed hESCs were followed for a minimum of 5 passages, to evaluate changes in their expansive behavior. By use of the same counting and gradual scoring system, as mentioned earlier, combined with the known split ratio, a total amount of colonies can be calculated for each passage. The mean of the total amount of colonies (experiments performed in triplicate) was plotted in a 3D bar chart. Next, these mean amounts of colonies were normalized to a reference sample in order to evaluate the changes in expansive behavior due to the cryopreservation process.
hESCs: size analysis
For size analysis, digital images were recorded of each of the cryopreservation experiments, at day 7 post-thawing. A minimum of a 100 randomly selected hESC colonies were measured using the Excellence® software of the Olympus microscope system (Olympus inverted Research System Microscope IX81, Brightfield, Objective 4×). A one-way ANOVA (Kruskal–Wallis) test was used to evaluate the differences. P<0.001 was considered significant.
Immunocytochemical analysis of hESC colonies
hESC pluripotency was determined at passage 5 by immunostaining for SSEA-3, SSEA-4, TRA 1-60, TRA 1-81, and Oct4 (isoform 4A) (antibodies from Santa Cruz Biotechnology, Inc., Heidelberg, Germany) and with alkaline phosphatase staining (BCIP/NBT Liquid Substrate System by Sigma). For the immunostaining, colonies were fixed in 1% paraformaldehyde for 10 min, washed with phosphate-buffered saline (PBS), and blocked with blocking serum (PBS/5% normal rabbit or goat serum/0.2% Tween) for 30 min. The primary antibodies that were used are rat monoclonal anti-SSEA-3 (IgM 1/200), mouse monoclonal anti-SSEA-4 (IgG 1/500), mouse monoclonal anti-TRA 1-60 (IgM 1/100), mouse monoclonal anti-TRA 1-81 (IgM 1/50), and mouse monoclonal anti-Oct4 (IgG 1/100). After 2 h of incubation, the cells were washed with PBS and incubated with the secondary antibody (FITC-labeled goat anti-mouse IgM 1/200 or biotin-labeled rabbit anti-mouse IgG) for 30 min. In case a biotin-labeled secondary antibody was used, visualization was achieved with streptavidin–FITC coupling (30 min). After washing, the cells were covered with PBS and evaluated by fluorescence microscopy (Olympus inverted Research System Microscope IX81, GFP filter, CellM software).
hESC embryoid body formation
Plates with full-grown hESC colonies were selected. The hESCs were detached from the feeder layer by incubation with collagenase IV solution (1 mg/mL) for 30–40 min. The colonies were pelleted by centrifuging for 3 min at 90 g, resuspended in embryoid body (EB) medium, and transferred to shaker flasks. The EB medium consisted of 80% DMEM-F12 (Invitrogen)+20% FBS (Invitrogen)+1 mM
Real-time RT-PCR analysis of hESC colonies and embryoid bodies
The colonies were detached from the feeder layer using collagenase IV solution (1 mg/mL; Invitrogen) for 30–40 min, collected, and centrifuged at 90 g for 3 min. After removal of the supernatant, TRI reagent was added to obtain lysis of the cells. The RNA was isolated using chloroform and further purified with isopropanol and ethanol. Afterward, DNA treatment was performed with a DNase kit (Invitrogen) and the RNA content and purity was examined by spectrophotometry. The RNA was transcribed to cDNA using the Reverse Transcriptase Core Kit (Eurogentec) on a Thermocycler (Applied Biosystems 2720 Thermocycler). Real-time PCR was performed on the ABI 7500 Fast Real-Time PCR device with ABI Taqman probes (Applied Biosystems) for the following genes: OCT4 (Hs00732896_s1), NANOG (Hs02387400_g1), HAND1 (Hs00231848_m1), COL2A1 (Hs01064869_m1), CD34 (Hs00990732_m1), RUNX2 (Hs00231692_m1), NESTIN (Hs00707120_s1), SOX9 (Hs00165814_m1), PAX6 (Hs01088112_m1), CERBERUS (Hs00193796_m1), AFP (Hs00173490_m1), and GATA4 (Hs00171403_m1). GAPDH was selected as endogenous control. Analysis of the gene expression was done using the ABI software for gene expression analysis on the ABI 7500 Fast Real-Time device.
Results
To eliminate the KO-SR from the cryopreservation medium, hydrolysates of different origin were tested to determine which had the highest potential as a protein substitute in the hESC cryopreservation process. For this screening test, the HS181 cell line (p42-p44) was used. Figure 1 shows the resulting mean recovery ratios after freezing with the different hydrolysates. Recovery ratios were obtained with all hydrolysates tested, indicating that all the hydrolysates are potentially useful for hESC cryopreservation. Clearly, the concentration of 8 mg/mL is overall the best concentration, independent of the hydrolysate origin. As the use of EX-CELL® CD hydrolysate fusion at a concentration of 8 mg/mL resulted in the highest recovery ratio, this solution was selected for further evaluation.
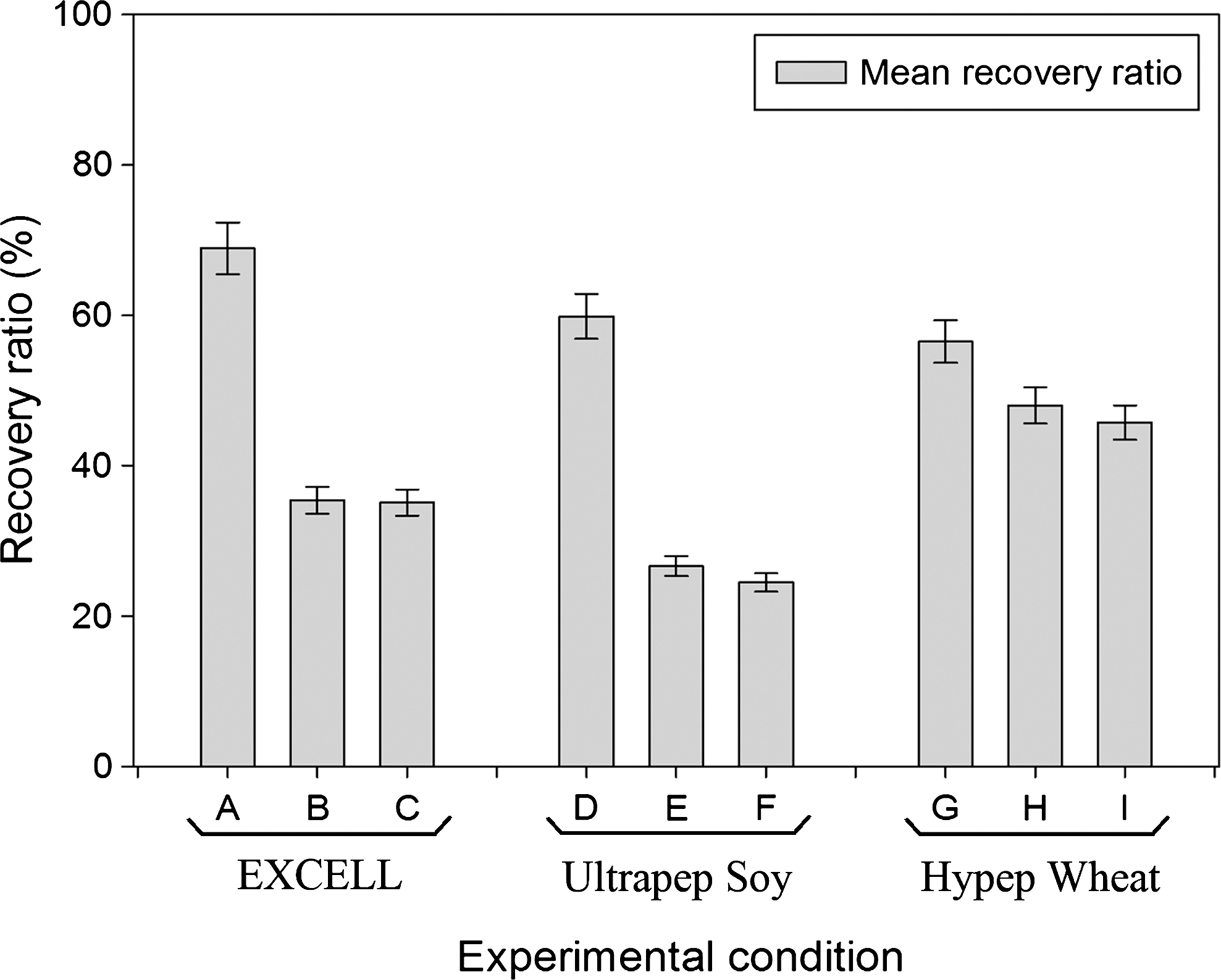
Overview of the mean recovery ratios, obtained after freezing hESCs without knockout serum replacement but with hydrolysates in the cryopreservation medium. Three different concentrations were tested of the selected hydrolysate substances: 8 mg/mL (A, D, G), 6 mg/mL (B, E, H), and 10 mg/mL (C, F, I). The results indicate that the use of 8 mg/mL is the overall best concentration, regardless of the hydrolysate origin. The best result was obtained with medium A, containing synthetic EX-CELL® CD Hydrolysate Fusion. hESCs, human embryonic stem cells.
An overview of the mean recovery ratios of test conditions 1–3 can be found in Fig. 2A. Our previously optimized protocol [27] (5% DMSO+5% HES+HM medium, thawed with 0.1 M sucrose-HM) for hESC slow freezing–rapid thawing led to a mean recovery rate of 168% for the H9 cell line, 101% for the BG01 cell line, and 86% for the HS181 cell line. Replacement of the HM medium with Excell medium in the cryopreservation solution (test condition 2) led to an important drop in recovery rate, leading to 59%, 21%, and 32% recovery ratio for the H9, BG01, and HS181 cell lines, respectively. The use of EX-CELL CD hydrolysate fusion in the cryopreservation and thawing solution (test condition 3) led to much better results: 137%, 111%, and 127% for the H9, BG01, and HS181 cell lines, respectively. Comparison of condition 1 with condition 3 revealed no statistically significant differences in the results (Student's t-test, P<0.05). The colony quality (Fig. 2B) was also investigated, as this is an important parameter for hESC culture. The colony quality was, in general, not influenced by the cryopreservation–thawing procedures. Overall, more than 95% of the colonies remained grade A.
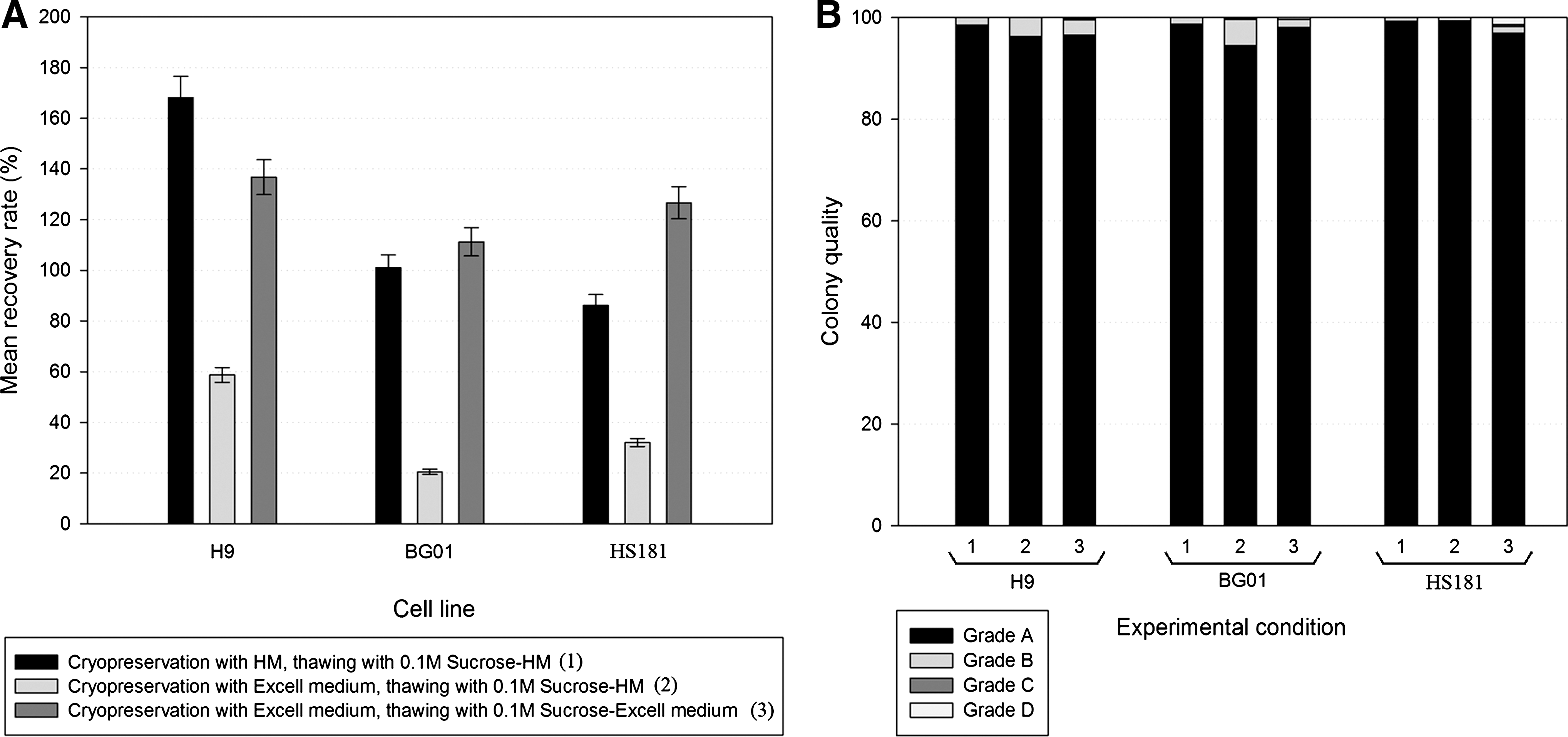
Overview of the mean recovery ratios and colony quality, obtained with the different experimental protocols for the H9, BG01, and HS181 cell lines.
The recovery ratios obtained with condition 3 are satisfactory; however, different strategies were further investigated to optimize the protocol and increase the recovery rate. The results can be found in Fig. 3A. Initially, a direct application of cell dissociation solution (condition 4) onto the colonies was tested, as this could reduce the overall costs and process time of the cryopreservation procedure. However, immediate application of cell dissociation solution onto the hESCs resulted in decreased recovery rates: 70%, 69%, and 39% for the H9, BG01, and HS181 cell lines, respectively. Additionally, application of condition 4 also resulted in a diminishment of the colony quality (Fig. 3B). More grade D colonies (dead colonies) could be found on the plates. Second, the addition of Y-27632 (condition 5) was investigated. This led to increased recovery rates for the 3 tested cell lines: 242%, 212%, and 282%. Additionally, the colony quality remained high. Finally, the combination of direct application of cell dissociation solution and addition of Y-27632 (condition 6) was also investigated. Here, low recovery ratios were obtained and there was a drop in colony quality, with the appearance of many grade D colonies.

Overview of the mean recovery ratios and colony quality after the optimization procedures for the H9, BG01, and HS181 cell lines.
The impact of the different cryopreservation strategies on the hESC colonies was evaluated. As colony size is an important factor when comparing different methods with counting strategies, we analyzed the colony size distribution for the different cryopreservation methods. The data, presented in Fig. 4, are from the HS181 cell line. Similar results were obtained for the H9 and BG01 cell lines. In general, unfrozen colonies (reference, day 7 of growth) appear to be slightly larger than cryopreserved–thawed colonies (day 7 post-thawing). However, this difference is not statistically significant. There is also no statistical difference in colony sizes between cryopreservation conditions 1, 2, 3, and 5. However, there is a clear statistical difference between these conditions and conditions 4 and 6 (one-way ANOVA test, P<0.001). The direct application of cell dissociation solution results in smaller colonies at day 7 after thawing. Size distribution analysis reveals that the largest variance in colony size can be found in the reference condition. Conditions 1, 2, and 3 show a similar size distribution. The colonies obtained with conditions 4, 5, and 6 are more uniform in their size.

Overview of the colony size analysis for the HS181 cell line. There is no statistical difference in size between the reference condition (unfrozen colonies) and conditions 1, 2, 3, and 5. Colonies of conditions 4 and 6 are statistically smaller than those of the other conditions, but also show a more uniform spreading.
The expansion of the colonies cryopreserved under different conditions was followed for 5 passages to affirm that there were no changes in expansive behavior. Figure 5 shows the expansion data of the HS181 cell line, as representative for all 3 cell lines tested. In Fig. 5A, the total amount of colonies versus the passage number and condition is plotted. A clear difference in mean amount of total colonies at passage 1 can be seen in Fig. 5A. These differences remain visible throughout the culture period. Conditions 3 and 5 show the highest expansion. Figure 5B shows the normalized amounts of colonies versus the passage number, using the initial conditions as a normalizing value to allow for fair comparison. No statistically significant differences in expansion could be detected (one-way ANOVA test, P<0.001).

Overview of the expansion of the HS181 cell line cryopreserved under different experimental conditions.
The pluripotent nature of the hESCs was checked by immunostaining and real-time PCR analysis after EB formation. Figure 6 shows the immunostaining of the HS181 cell line, p5, after cryopreservation with condition 3. Positive results were obtained for SSEA-3 (Fig. 6A), SSEA-4 (Fig. 6B), TRA 1-60 (Fig. 6C), TRA 1-81 (Fig. 6D), and Oct4 (Fig. 6E). The cells also expressed a high amount of alkaline phosphatase (Fig. 6F). Real-time PCR analysis of EBs, formed from hESCs previously frozen with condition 3, was also performed to verify the differentiation capacity of the cells. The derived EBs showed a decrease in expression of Oct4 and Nanog, indicating that differentiation is occurring. There is an upregulation of ectodermal markers Nestin, Sox9, and Pax6. Upregulation of Hand1, Col2A1, CD34, and Runx2 indicates differentiation along the mesodermal lineage. Upregulation of AFP and GATA4 indicates differentiation along the endodermal lineage. Cer1 is downregulated at this stage.

Pluripotency evaluation of the hESC colonies previously frozen with synthetic hydrolysates (condition 3):
Discussion
The aim of the present study was to investigate whether hydrolysates could substitute KO-SR or FBS/FCS in hESC cryopreservation. From the perspective of therapeutic applications of hESC or hESC-derived cell products, the culture and banking of hESCs should be performed under GMP conditions. GMP is a quality assurance system that ensures that the end product, in this case a cellular therapy product, meets its preset specifications and thus is safe to apply in a clinical setting. By extensive control over the manufacturing and testing process, validation of devices, raw materials, used reagents, and protocols combined with risk analysis regarding quality and safety, GMP-approved products can be made. For cellular therapy products, the elimination of all animal-containing products from the culture and cryopreservation process is one of the issues that needs to be tackled. Many strategies can be applied to obtain animal-free cryopreservation conditions, such as the application of proprietary commercially available solutions [5,19] or the addition of small molecules or growth factors [11,19,30,31]. However, these options can be quite expensive, especially when large amounts of hESCs need to be frozen and stored. We chose to investigate the application of hydrolysates, as these are inexpensive, easily available, less dependent on batch-to-batch variation, and much safer to use than animal-containing substances.
The use of hydrolysates is not uncommon in cell culture and cryopreservation. A range of hydrolysates of different origin (soy, wheat gluten, yeast, meat, rice, etc.) are commercially available, each having their own beneficial effect on different cell types [21,22,25,26,32,33]. Hydrolysates can act as serum or KO-SR substitute, not merely by providing growth-promoting peptides and amino acids, but also by providing key carbohydrates, lipids, minerals, and vitamins that can improve the culture conditions by their osmoprotective and antiapoptotic characteristics [20 –22,25,33]. Hydrolysates are generally produced through enzymatic processes or hydrolysis, followed by ultracentrifugation and size-exclusion chromatography, so that quite pure hydrolysate products arise. Additionally, regarding the safety aspect, it is generally accepted that plant pathogens that might be present in the produced hydrolysates are not immunogenic for mammalian cells. A known exception is the presence of immunogenic plant N-glycans [34]. However, it should be examined in clinical trials whether this would be a concern, and moreover, the extensive processing of the hydrolysates during production nearly eliminates this problem.
To our knowledge, plant hydrolysates have not been used in hESC culture and cryopreservation, with the sole exception of a recent article investigating the use of spelt wheat-derived Vegetacell [26] in hESC culture. Our focus was on hESC cryopreservation. Our study included hydrolysates from soybean (Ultrapep Soy), wheat gluten (Hypep 4601), and synthetic origin (EX-CELL CD Hydrolysate Fusion). Figure 1 of our results clearly indicates that all 3 substances have the potential to be used in hESC cryopreservation. Because the use of EX-CELL CD Hydrolysate Fusion led to the highest recovery rate, further tests were performed with this product. Additionally, as EX-CELL CD Hydrolysate Fusion is a synthetic formulation of hydrolysates, based upon data generated from 4 hydrolysates types (soy, wheat gluten, yeast extract, and meat), there is no problem with batch-to-batch variability or with cross-contaminations of potential plant pathogens or immunogenicity.
A concentration of 8 mg/mL appeared the most optimal for our application. This concentration is quite high compared with the hydrolysate concentrations for bovine embryo cryopreservation [23,24]. Here, concentrations of 1.8–5.6 mg/mL were adequate for comparable embryo development and hatching to the use of bovine serum albumin. Most manufacturers recommend adding between 1 and 6 mg/mL of the hydrolysates to the culture medium. Nevertheless, it is always advised to examine the concentration for each application and optimize it for each cell type. Additionally, as the hydrolysate is applied in a cryopreservation process, other concentrations might be beneficial than those applied in a culture setting. It is clear from the data (Fig. 2) that the application of EX-CELL CD Hydrolysate Fusion in both the cryopreservation medium and the thawing solution (condition 3) is feasible, and that the obtained recovery ratios are better than with a single application in the cryopreservation solution. There is no loss in colony quality related to the cryopreservation process. The obtained recovery ratios are quite comparable to the recovery ratios obtained with KO-SR. Depending on the hESC line used, there is a variation between condition 1 and 3 of 20% reduction to 47% increase in recovery ratio. Statistical analysis revealed that these differences are not significant (Student's t-test, P=0.05), and thus, EX-CELL CD Hydrolysate Fusion can substitute KO-SR in the cryopreservation medium without loss in recovery rate and quality.
Further optimization strategies were examined to simplify the protocol and increase the recovery rates. A direct application of cell dissociation solution onto the hESC plates would not only simplify the procedure but would also eliminate the use of collagenase IV, thus realizing a less time-consuming and more economic approach. Unfortunately, this approach (Fig. 3, condition 4) resulted in low recovery rates and a reduction in colony quality. We suggest that this might be caused by the additional feeder cells, which are present in the cryopreservation mixture and interfere with the reach of an optimal equilibrium during the freezing process. Addition of the Rock-inhibitor Y-27632 led to an increase in recovery ratio. This was expected, as the survival enhancing effect of Y-27632 has been well documented in hESC cryopreservation and culture [31,35]. The recovery ratios almost doubled for all tested hESC lines and the overall colony quality remained high. However, a combination of both strategies (condition 6) did not result in high recovery ratios. The obtained recovery rates in this condition were sometimes even lower compared with those of condition 4. Although this result was consistently observed in all 3 cell lines, we cannot explain it.
This study was performed with hESCs that were routinely cultured on MEF feeders. The animal origin of the feeder also poses a safety problem with regard to future applications. However, instead of MEFs, clinical-grade human foreskin fibroblast (HFF) feeder cells could be used and would be a safe option for therapeutic hESC culture [4]. The switch of traditional hESC culture on MEF to culture on large-scale expanded HFFs can be easily performed [36]. Another option is the combination with feeder-free alternatives. As cryoprotectors in our protocol, we applied 5% DMSO and 5% HES, as previously optimized [27]. A clinical-grade version of DMSO is available and HES, which is commonly used for the clinical cryopreservation of cord blood stem cells and bone marrow stem cells, has also been approved for clinical use. Additionally, hydrolysates are already extensively applied in vaccine production and are thus acceptable in production processes for clinical applications. The proposed protocol can thus be easily modified to be compatible with GMP standards.
The obtained cells, frozen with condition 3, were analyzed by immunocytochemistry and real-time PCR. The hESCs were positive for SSEA-3, SSEA-4, TRA 1-60, TRA 1-81, and Oct4. They also expressed high levels of alkaline phosphatase. Real-time PCR analysis of derived EBs showed that the hESCs could still differentiate into ectodermal, mesodermal, and endodermal lineages. Overall, the hESCs remained pluripotent.
In conclusion, the application of hydrolysates of vegetal or synthetic origin is feasible for hESC cryopreservation. The use of synthetic EX-CELL CD Hydrolysate Fusion in both the cryopreservation and the thawing solution led to high recovery rates, comparable to those obtained with the addition of KO-SR. Optimization of the protocol by addition of Y-27632 doubled the previously obtained recovery rates. This protocol is suitable for xeno-free large-scale hESC cryopreservation, ideal for future therapeutic applications.
Footnotes
Acknowledgments
This study was conducted as part of a Ph.D. project, funded by BOF (BOF08/24J/063). The authors express their gratitude to Professor Outi Hovatta (Karolinska Institutet, Sweden) for donating the HS181 hESC line. The authors thank Johanna Aernoudt, Greet De Smet, and Leen Pieters, lab assistants, for their practical aid in realizing this work. An abstract regarding preliminary data of the cryopreservation procedures has been published as a supplement of the November 2011 issue of Regenerative Medicine.
Author Disclosure Statement
There are no potential conflicts of interest.