
Abstract
Adipose-derived stem cells (ADSCs) are attracting increased attention as a novel source in regenerative medicine. Transplantation of ADSCs promotes functional recovery in animal models of peripheral nerve injury, but the mechanism of enhanced nerve regeneration remains to be elucidated. In addition, it is important to examine whether the supportive functions of ADSCs are dependent on donor age or anatomic site of origin. In this study, we examined the effects of factors produced by mouse ADSCs on Schwann cells (SCs) and dorsal root ganglion (DRG) neurons in vitro and compared these effects among ADSCs from donors of different age and from different anatomic regions. ADSC-derived soluble factors supported survival and proliferation of SCs and promoted neurite outgrowth in DRG neurons. These beneficial effects were far superior to that of factors from 3T3-L1 cells and comparable to those of SC- and astrocyte (AC)-derived factors. ADSCs from different sources similarly retained their neurotrophic activity. Real-time reverse transcription-polymerase chain reaction and enzyme-linked immunosorbent assay analyses demonstrated that ADSCs produced various growth factors, some of which were more abundant than in SCs and ACs. These results suggest that ADSCs promote peripheral nerve regeneration partly through paracrine secretion of trophic factors and regardless of donor age or anatomic site of origin.
Introduction
T
Peripheral nerve injury is a common manifestation of various traumatic insults. Despite the relatively higher regenerative capacity of peripheral nerves compared to the central nervous system, recovery is frequently partial and functional deficits persist. Incomplete regeneration of peripheral nerves is attributable to several factors, including loss of Schwann cells (SCs), fibrotic scarring, and misguided axonal projections [6,7]. Therefore, development of therapies to overcome these obstacles is needed. ADSCs are promising candidates for autologous cell therapy because of their high yield and relatively simple isolation. There is abundant experimental evidence showing that implantation of ADSCs promotes axonal regeneration of peripheral nerves after injury [8 –10], but the precise mechanism through which ADSCs induce this regeneration remains unclear. Most previous studies have focused on the transdifferentiation potential of ADSCs [11,12], including differentiation into Schwann-like cells in vitro and subsequent grafting for cell replacement therapy [13,14]. However, it is unclear whether ADSCs act as truly functional SCs in vivo and if this is critical for their ability to induce regeneration.
In addition to their multipotency, recent studies have revealed a novel function of ADSCs as small molecular factories. ADSCs produce many different kinds of cytokines and growth factors that promote host cell survival and modulate immune functions [15 –17]. These supportive and modulatory effects of ADSCs are superior to those of BM-MSCs and suggest a novel application of ADSCs in regenerative medicine [18]. For clinical application for autologous cell therapy, one important issue to be resolved is how the properties of ADSCs vary depending on donor age and anatomic site of origin. For example, it has been reported that the yield of ADSCs and their differentiation potential are both reduced with age in humans and mice [19 –22]. Also, the subset of adipocytes in the subcutaneous tissue of the head originates from neural crest cells, whereas a mesoderm origin is assumed for adipocytes in subcutaneous fat deposits of the trunk [23]. The influence of age and anatomic site on the function of ADSCs remains largely unknown.
To clarify whether secretion of growth factors by ADSCs contributes to regeneration of peripheral nerves, we studied the effects of mouse ADSC-conditioned medium (CM) on SCs and dorsal root ganglion (DRG) neurons in vitro, and compared the results for ADSCs from mice of different ages and from different anatomical sites.
Materials and Methods
Isolation and culture of mouse ADSCs
ADSCs were isolated as previously described [1,2], from 5 different subcutaneous adipose depots of male C57BL/10J mice: from trunk (abdominal and inguinal) fat pads in 13–16-month-old mice (Old-T), 8-week-old young adult mice (Ad-T), and postnatal 5-day-old neonatal mice (Neo-T), and from cephalic (head and neck) fat pads in 1-year-old mice (Old-C) and 8-week-old young adult mice (Ad-C). Cephalic fat pads from 1-week-old mice were not used because the amount of cells was too small to be isolated and cultured (Fig. 1A).
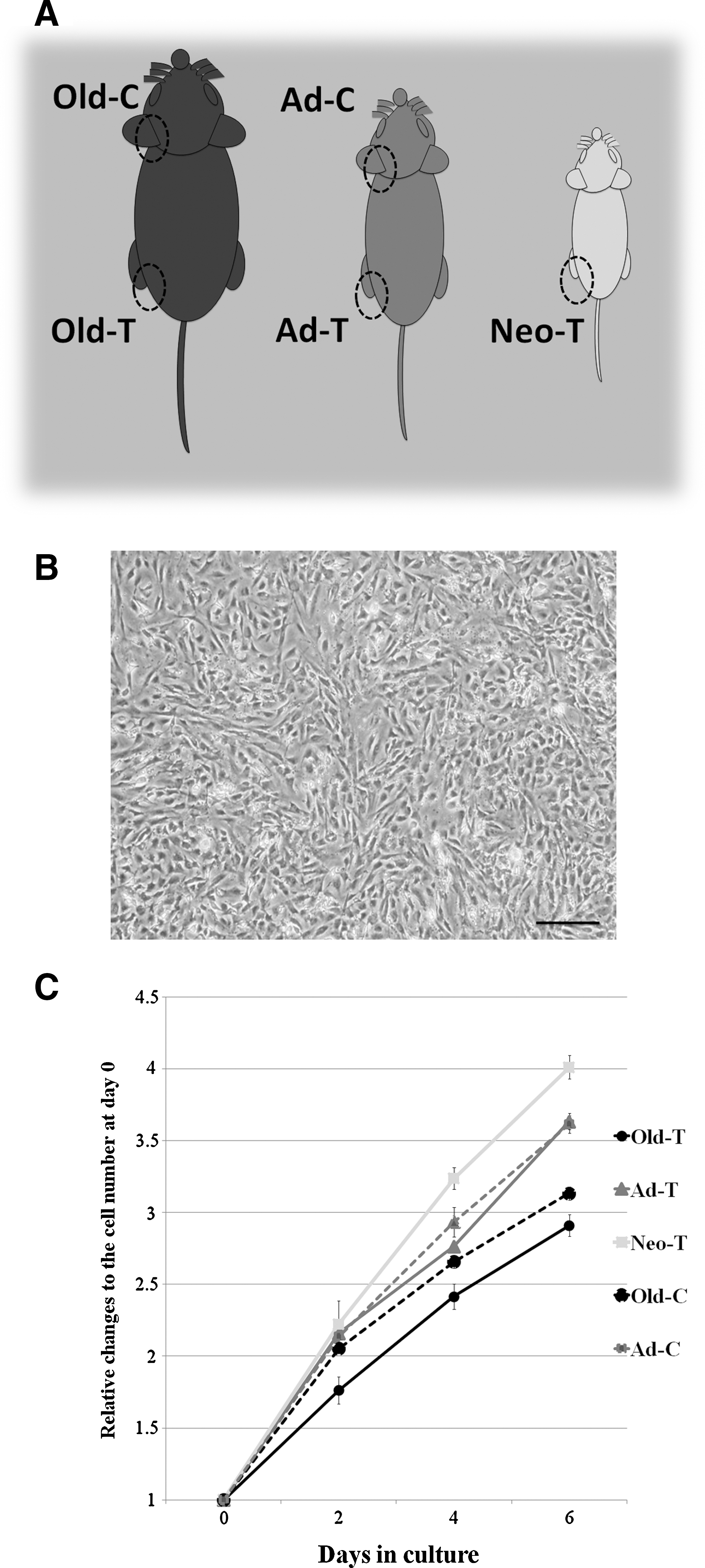
Isolation and culture of adipose-derived stem cells (ADSCs) from different adipose depots.
Mice were housed under a 12-h light/dark cycle in an SPF facility with controlled temperature and humidity, and were allowed access to food and water ad libitum. Experiments were conducted according to protocols approved by the Committee for Animal Research, Kyoto Prefectural University of Medicine, Japan, and the animals were handled in accordance with the guidelines for the Care and Use of Laboratory Animals of Kyoto Prefectural University of Medicine, which comply with the Guidelines for Proper Conduct of Animal Experiments issued by the Science Council of Japan.
For isolation of ADSCs, adipose depots from each group of mice were minced and digested using 0.075% collagenase type I (Sigma-Aldrich) for 45 min at 37°C. Enzyme activity was neutralized by treatment with Dulbecco's modified Eagle's medium (DMEM; Nacalai Tesque) containing 10% fetal bovine serum (FBS). The cell suspension was centrifuged at 1,200 g for 5 min to separate floating adipocytes from the stromal vascular fraction (SVF). After filtration of the suspension through a 40-μm nylon strainer to remove cellular debris, the SVF cells were resuspended in DMEM with 10% FBS and 100 U/mL penicillin–streptomycin (Nacalai Tesque) and plated onto plastic culture dishes at a density of 1×104/cm2, followed by incubation at 37°C in 5% humidified CO2. After 24 h, nonadherent cells were removed by a medium change. Fresh culture medium was added and replaced every 3 days. After 7 days (80%–90% confluency) the cells were treated with 0.25% trypsin (Gibco) and diluted 1:3 per passage for further expansion. The ADSCs at passage 2 were seeded at a density of 1×104/cm2 in DMEM containing 10% FBS.
Characterization of ADSCs from different subcutaneous adipose depots
Flow cytometry was used to characterize ADSCs obtained from each adipose tissue depot after 2 passages. The cells were treated with 0.25% trypsin and 0.01% EDTA for 5 min, washed 3 times with phosphate-buffered saline, and allowed to react with the following primary antibodies (applied in optimal amounts): Alexa Fluor 488-conjugated hamster anti-CD29 antibody (Biolegend), Alexa Fluor 488-conjugated mouse anti-CD44 antibody (BD PharMingen), FITC-conjugated rat anti-Sca-1 antibody (BD PharMingen) as MSC markers, and PE-Cy5-conjugated mouse anti-CD45 antibody (eBioscience) as a hematopoietic cell marker using a FACSCalibur flow cytometer and CellQuest software (Becton Dickinson). Isotype antibody control samples for each cell population were used to set the dot-plot intercepts used for the analysis.
ADSCs were also characterized immunocytochemically with antibodies against fibronectin and the endothelial cell marker CD31. The multilineage differentiation potential of each group of ADSCs was also assessed using a Mouse Mesenchymal Stem Cell Functional Identification Kit (R&D Systems), which was used according to the manufacturer's protocol at about the same time as immunocytochemical characterization. The total cell number was also measured with the FACSCalibur flow cytometer to obtain a growth curve for each group of ADSCs.
Preparation of CM
When the second-passage cells reached 80% confluency, the medium was replaced with DMEM containing 2% FBS, followed by culture for an additional 48 h. The supernatant of the culture medium was then collected with filtration through a 40-μm filter (BD Falcon), thus yielding ADSC-CM. To eliminate the influence of variations in biological characteristics (eg, secretion patterns) between different ADSC donors, CM was collected from 6 independent groups (3 mice per group). CM from 3T3-L1 cells (3T3-L1-CM), astrocytes (AC-CM), and SCs (SC-CM) was obtained by the same procedure for comparison with ADSC-CM. 3T3-L1 cells were purchased from ATCC (no. CL173). The procedures of SC and astrocyte (AC) cultures are described below.
Culture of mouse SCs
SCs were isolated from the sciatic nerves of 5-day-old C57/BL6 mice, as described previously [24]. The isolated SCs were digested with 1% collagenase type I (Sigma-Aldrich) for 30 min at 37°C. After incubation, SCs were rinsed with DMEM containing 5% FBS, mechanically dissociated in standard culture medium, and seeded on laminin-coated 8-chamber dishes at a cell density of 5×103/cm2. A purity of >90% SCs was obtained using 2 rounds of purification to eliminate contaminants, in combination with use of cytosine-B-arabinoside hydrochloride (Ara-C; Sigma-Aldrich) and differential cell detachment. The purity was confirmed by cell morphology and immunostaining with anti-S-100 antibody (Dako). SCs were treated with a 1:1 mixture of ADSC-CM and control medium (Neurobasal medium, supplemented with 0.5 mM
Culture of mouse DRG neurons
DRG neurons were obtained from 7-day-old C57/BL6 mice, using the protocol established by Malin et al. [25]. Briefly, DRGs from the cervical to the lumbar level were dissected from each animal and dissociated into single cells with 0.7% collagenase type I and 0.25% trypsin.
The cells were further purified by density gradient centrifugation (5 min, 200 g) with 30% Percoll (Sigma) to eliminate the myelin sheath, followed by culture with Ara-C to reduce the number of non-neuronal cells such as SCs, satellite cells, and fibroblasts.
Culture of mouse cortical ACs
Primary AC cultures were prepared as described previously [26]. Briefly, 1-day-old C57/BL6 mouse cortex was dissected, dissociated, and seeded in DMEM with 10% FBS. AC-enriched culture was established by shaking methods and the experiments were performed at passage 2.
Immunocytochemistry
Immunocytochemistry was performed as previously described [27]. Briefly, cells were fixed with 4% paraformaldehyde for 30 min, followed by overnight incubation at 4°C with the following primary antibodies: rabbit anti-fibronectin (Calbiochem), mouse anti-CD31 (BD PharMingen), rabbit anti-S-100 (Dako), rabbit anti-β-III tubulin (Covance), and mouse anti-BrdU (clone G3G4; Developmental Studies Hybridoma Bank). The cells were then incubated with Alexa Fluor-tagged secondary antibodies Alexa 488 or Alexa 568 (Invitrogen). For the BrdU incorporation assay, DNA was denatured with 2 N HCl for 20 min at room temperature. Nuclei were counterstained with 4′,6′-diamidino-2-phenylindole dihydrochloride (DAPI). The cells were observed and photographed under a fluorescent microscope (Nikon E1000 Eclipse).
Assays of proliferation and viability of SCs
After 24 h of incubation with ADSC-CM, SC proliferation was quantified using a BrdU incorporation assay. SCs were cultured in the presence of 10 μM BrdU for 12 h followed by immunocytochemical analysis, as described above. The proportion of cells with BrdU incorporation among total cells was quantified under a fluorescent microscope (Nikon E1000 Eclipse) in 3 independent cultures (>2,000 cells of 5 randomly selected fields in each culture; n=6 independent experiments for each condition). SC viability was assessed using a WST-1 assay according to the manufacturer's protocol (Takara) after 3 days of incubation in each CM group (n=6 independent experiments for each condition). Cell viability is expressed as the absorbance OD value.
Measurements of neurite outgrowth in a DRG neuron bioassay
Digital images of DRG neurons stained with anti-β-III tubulin antibody were transferred into image analysis software (NIH ImageJ v.1.37v) for neurite morphometric analyses. Primary neurites were defined as processes directly emerging from the cell body. These processes usually have a thicker diameter than branching neurites. All primary and branching neurites were manually traced on the digital images. The following parameters were measured, as previously described [28]: (1) mean percentage of neurite-bearing cells, (2) mean number of primary neurites per single DRG neuron, (3) mean total primary neurite length per single DRG neuron, and (4) mean number of branching neurites per single DRG neuron. The total DRG neuron density (calculated as the total number of DRG neurons/cm2) was also measured. The parameters were measured at 24 h after culture, since neurite outgrowth was too extensive to be measured after this time point. Four independent cultures were analyzed for each condition, with data collected from at least 25 cells per coverslip and a minimum of 12 coverslips per condition.
Real-time quantitative polymerase chain reaction
Real-time quantitative polymerase chain reaction (RT-qPCR) was performed using the ABI PRISM® 7000 Sequence Detection System (Applied Biosystems). The following oligonucleotide primers were used for PCR analysis of gene expression in ADSCs and 3T3-L1 cells: brain-derived neurotrophic factor (BDNF) (NM_007540) forward CAG GTT CGA GAG GTC TGA CGA, BDNF reverse CGC GTC CTT ATG GTT TTC TTC G; nerve growth factor (NGF) (NM_001112698) forward GGA GCG CAT CGA GTT TTG GCC, NGF reverse ACG CTA TGC ACC TCA CTG CGG; glial growth factor (GGF) (NM_178591) forward GAA TTT ATG GAA GCG GAG GA, GGF1 reverse CAG TAG GCC ACC ACA CAC AT; hepatocyte growth factor (HGF) (NM_010427) forward ATT TCC CAG CTG GTC TAT GG, HGF reverse TAA TAA ACC ATC CGC GTT GA; insulin-like growth factor-1 (IGF-1) (NM_008084) forward TGA GCT GGT GGA TGC TCT TCA GTT, IGF reverse TCA TCC ACA ATG CCT GTC TGA GGT; vascular endothelial growth factor (VEGF) (NM_001025250) forward CCA CGT CAG AGA GCA ACA TC, VEGF reverse TCT CCT ATG TGC TGG CTT TG; and GAPDH (NM_013633) forward TGT GTC CGT CGT GGA TCT GA, GAPDH reverse TTG CTG TTG AAG TCG CAG GAG. All measurements were performed in triplicate. Expression levels were normalized using the expression level of the GAPDH housekeeping gene. Each cDNA underwent the reaction in a 25.0 μL volume containing 0.5 μL cDNA template and 24.5 μL PCR solution, using an RNeasy Mini Kit (Qiagen). The PCR conditions were 2-min incubation at 95°C, followed by 40 cycles at 95°C for 5 s and 60°C for 34 s. A relative amount of transcript was estimated by the standard curve method.
Enzyme-linked immunosorbent assay
The supernatants from cultured ADSCs (after 2 passages), ACs, SCs, and 3T3-L1 cells were analyzed by enzyme-linked immunosorbent assay (ELISA) to determine the concentration of BDNF, NGF, and VEGF. Sandwich ELISA kits for mouse BDNF and NGF (Promega) and for mouse VEGF (RayBiotech) were used according to the manufacturer's specifications and analyzed in duplicate in 3 independent experiments. A standard curve was plotted to determine their concentrations.
Statistical analysis
All data are expressed as means±SEM. One-way ANOVA and Tukey multiple comparison tests were used to compare differences between groups. All analyses were conducted with GraphPad Prism 5.0 software (GraphPad). Differences were considered significant at *P<0.05, **P<0.01, and ***P<0.001 versus control; # P<0.05, ## P<0.01, and ### P<0.001 versus 3T3-L1; $ P<0.05, $$ P<0.01, and $$$ P<0.001 versus ACs; and + P<0.05, ++ P<0.01, and +++ P<0.001 versus SCs.
Results
Characterization of mouse ADSCs from different adipose tissue depots
We first characterized the growth kinetics and immunophenotypes of ADSCs cultured from mice of different ages and from different anatomic sites. In our isolation and culture methods, ADSCs from all adipose tissue depots expanded easily in vitro and exhibited similar fibroblast-like morphology to that of BM-MSCs (Fig. 1B). There was no significant difference in growth rate between ADSCs from the 2 anatomic sites, but the growth of ADSCs significantly declined with the age of the animal (P<0.05). However, all ADSCs were able to be expanded over at least 5 passages (Fig. 1C).
Immunofluorescent staining showed that all ADSCs homogeneously expressed a fibroblast marker, fibronectin, but were negative for an endothelial cell marker, CD31 (Fig. 2A). Flow cytometry analysis revealed that the vast majority of cells in the ADSC cultures were immunopositive for mouse MSC markers, CD29, CD44, and Sca-1, but few were positive for the hematopoietic cell marker, CD45. There were no significant differences in the expression of these antigens among ADSC cultures from mice of different ages and from different anatomic sites (Fig. 2B). Thus, all ADSC groups were a relatively homogenous cell population after the second passage, with little or no contamination with hematopoietic/endothelial cells. We also confirmed that all ADSCs had the potential to differentiate into adipocytes, osteocytes, and chondrocytes (data not shown). Collectively, these results show that ADSCs exhibit similar characteristics in vitro regardless of age and anatomic site, except for a modest decline in growth rate with age.
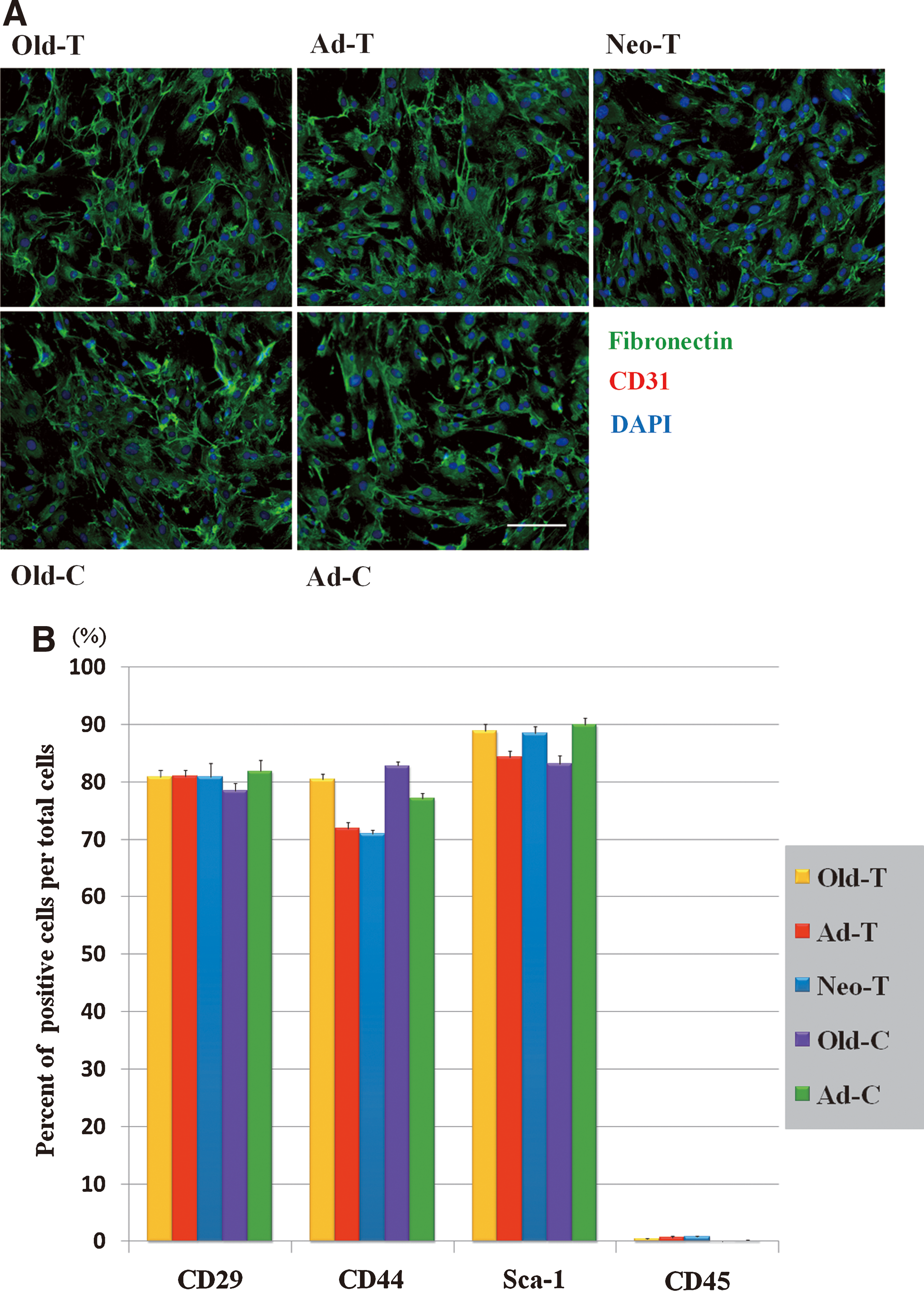
Characterization of ADSCs from different adipose depots.
Effects of ADSC-CM on mouse SCs
Survival and proliferation of SCs is an important aspect of peripheral nerve regeneration in vivo. We first analyzed the effects of factors derived from ADSCs on the survival and proliferation of SCs in vitro. The cell density of SCs treated with ADSC-CM for 3 days was visibly higher than that of control SCs. At a single-cell level, both cultures exhibited a typical bipolar SC morphology, but those treated with ADSC-CM had longer processes than control SCs (Fig. 3A). A WST-1 assay demonstrated that the cell densities of SCs treated with ADSC-CM for 36 h were more than twice that with control culture and more than 1.3 times as high as that after treatment with 3T3-L1-CM. There were no significant differences among SCs treated with ADSC-CM from different sources and AC-CM (Fig. 3B).
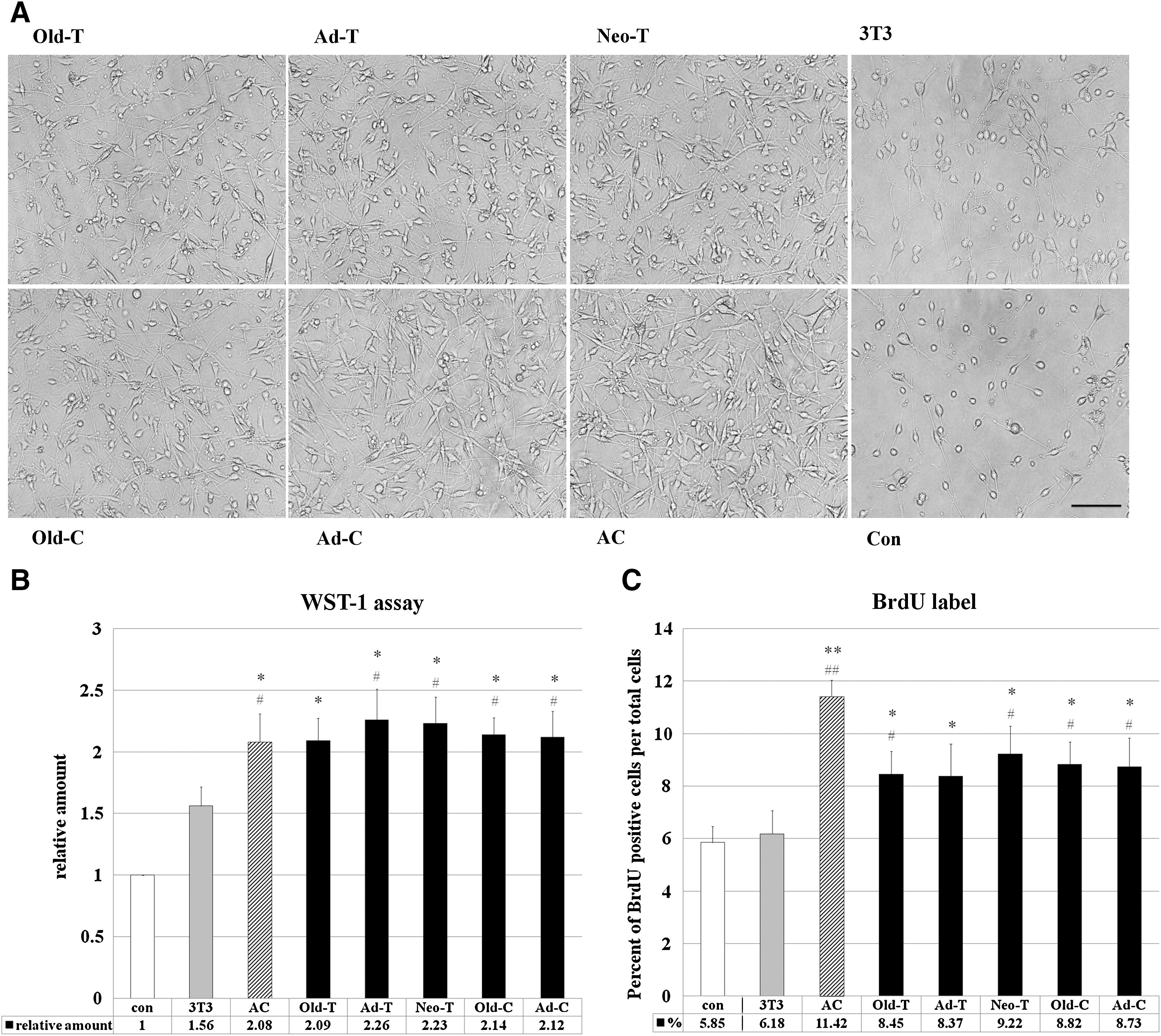
Effects of ADSC-conditioned media (CM) on Schwann cells (SCs) in vitro.
The higher cell density in cultures treated with ADSC-CM can be attributed to an increase in cell survival, induction of cell proliferation, or both. We therefore quantified SC proliferation with a BrdU incorporation assay. The proportions of BrdU-positive cells in ADSC-CM-treated SC cultures were lower than those of AC-CM-treated cultures (∼75%) but higher than those of control cultures and 3T3-L1-CM-treated cultures (∼1.5-fold and 1.4-fold higher, respectively). There were no significant differences among ADSC-CM groups (Fig. 3C). Since the proportions of BrdU-positive cells in all groups examined were relatively low, it is likely that both an increase in cell survival and induction of cell proliferation contributed to the higher cell density.
Effects of ADSC-CM on DRG neurons
We next examined whether ADSC-derived factors have a direct influence on neurite outgrowth of DRG neurons in vitro. Photomicrographs of DRG neurons stained with anti-β-III tubulin antibody showed that treatment with ADSC-CM from all groups produced many longer primary neurites with increased branching, compared to control and 3T3-L1-CM-treated neurons (Fig. 4A). Quantitative analysis showed that >80% of DRG neurons in ADSC-CM-treated cultures were neurite-bearing cells, whereas only approximately half of the cells had extended neurites in 3T3-L1-CM and control medium (58.5%±10.9% and 49.4%±11.2%, respectively) (Fig. 4Bi). ADSC-CM-treated cultures also showed significantly higher values for the mean number of primary neurites, the total primary neurite length, and the mean number of branching neurites per single DRG neuron, compared to these values in 3T3-L1-CM and control cultures (Fig. 4Bii–iv). For example, the total length of neurites was 2.5 and 4 times greater in ADSC-CM cultures compared to 3T3-L1-CM and control cultures, respectively. These positive effects of ADSC-CM in all parameters were closely similar to those of SC-CM and AC-CM, both of which are known to induce neurite outgrowth [29]. There were no significant differences among all ADSC groups, but the youngest ADSCs (Neo-T) tended to have greater effects. 3T3-L1-CM also increased all parameters compared to control medium, suggesting that 3T3-L1-CM has modest effects on neurite outgrowth [30,31].

Effects of ADSC-CM on dorsal root ganglion (DRG) neurons.
We also analyzed the influence of ADSC-CM on survival of DRG neurons to test the possibility that induction of neurite outgrowth is due to an increase in cell survival. The densities of β-III tubulin-positive DRG neurons did not differ significantly among all groups at the time point analyzed (Fig. 4C). Thus, it is likely that ADSC-CM directly promotes neurite outgrowth.
Expression and secretion of growth factors in ADSCs
The results described above demonstrate that ADSC-derived factors possess much greater ability to induce SC survival and growth and DRG neurite outgrowth compared to 3T3-L1-derived factors, whereas there were no significant differences among the ADSC groups. To explore the potential factors involved in these effects, we first compared the expression levels of mRNA for BDNF, NGF, GGF, HGF, IGF-1, and VEGF in ADSCs from trunk fat pads in 8-week-old mice: Ad-T (a typical age and site for donor cells) with those in SCs, ACs, and 3T3-L1 cells, using RT-qPCR. The reason we chose these factors were that they have been reported to enhance the survival and proliferation of SCs and stimulate axonal outgrowth [32 –37]. For all growth factors analyzed, the levels of mRNA in ADSCs were much higher than those in 3T3-L1 cells. Furthermore, this analysis showed significantly higher levels of mRNAs for GGF (18.5-fold) and HGF (18.9-fold) in ADSCs compared with SCs, and for IGF (30.5-fold and 2.5-fold) and VEGF (2.8-fold and 2.3-fold) in ADSCs compared with SCs and ACs, respectively (Fig. 5A). We also quantified 3 growth factors at protein level in culture supernatants using ELISA. This assay confirmed the secretion of all 3 factors by ADSCs; BDNF (58.1±7.1 pg/mL), VEGF (569.7±14.3 pg/mL), and NGF (47.01±0.1 pg/mL). The concentrations of all factors were higher in ADSC-CM than in others. For example, a larger amount of VEGF was significant compared with SC-CM (143.1±5.4pg/mL) and 3T3-L1-CM (204.5±5.9pg/mL) (Fig. 5B). These results show that the production of a variety of trophic factors by ADSCs is much higher than 3T3-L1 cells and is at least comparable to SCs and ACs.
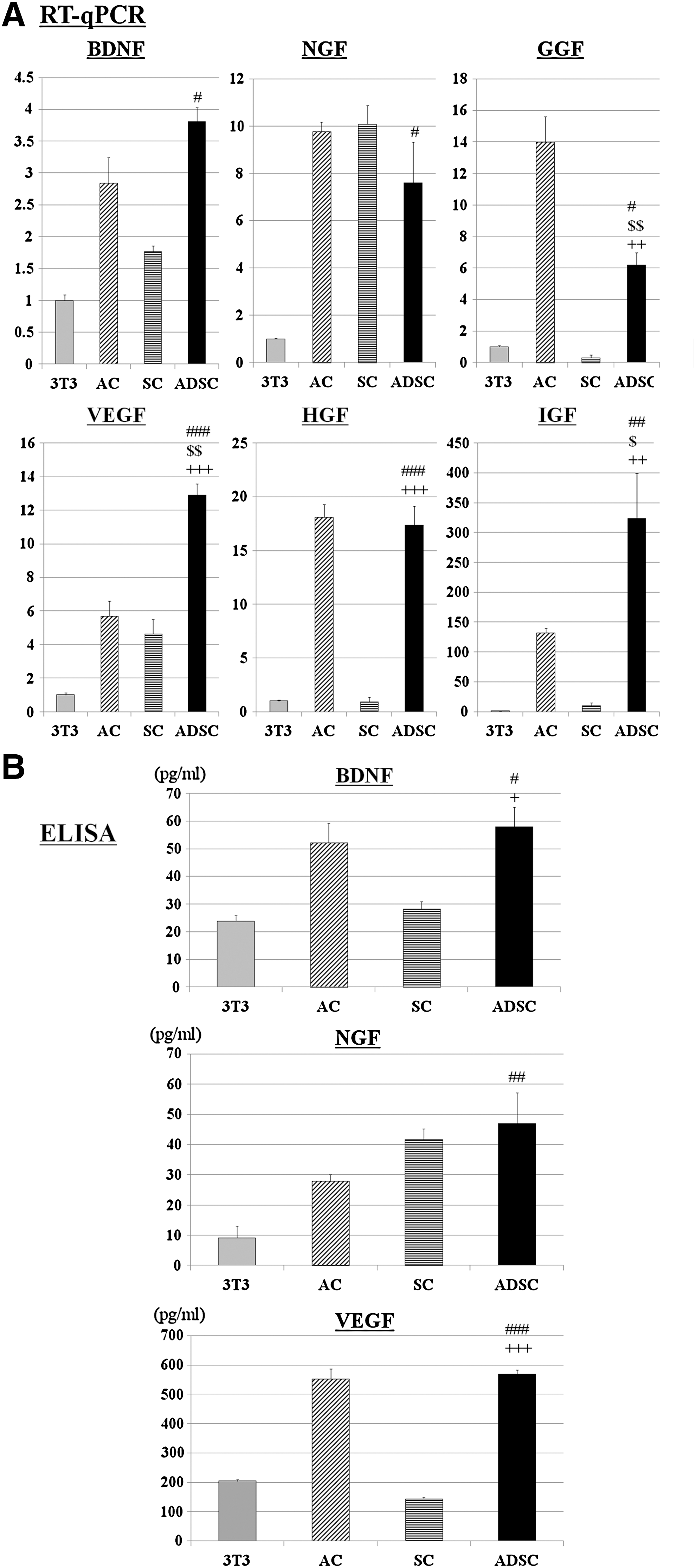
Comparative analysis of mRNA and protein levels of growth factors in ADSCs, SCs, ACs, and 3T3-L1 cells.
Discussion
Several previous studies have shown that transplantation of ADSCs promotes axonal regeneration and functional recovery in animal models of peripheral nerve injury [8 –10], but the mechanisms of such effects remain poorly understood. In most studies, ADSCs have been differentiated toward Schwann-like cells in vitro before grafting, but it remains to be elucidated whether these cells replace host cells to act as functional SCs. For example, neural-like cells chemically induced from BM-MSCs are not similar to neural cells and still resemble undifferentiated BM-MSCs judging from the gene expression profiles [38]. Ribeiro-Resende et al. have shown that transplanted BM-MSCs fail to differentiate into SCs in vivo. Instead, BM-MSCs markedly increase the number of invading macrophages, resulting in acceleration of debris removal and increase the local concentration of trophic factors [39]. Our results may favor the assumption that enhanced secretion of neuroprotective/neurogenerative factors is likely to underlie accelerated nerve regeneration after the transplantation of ADSCs. We thus suggest that undifferentiated ADSCs produce factors that enhance the survival and proliferation of SCs and promote axonal growth in peripheral neurons in vivo. The effects of the undifferentiated ADSC-derived factors were far superior to those of 3T3-L1 cells, and were comparable to SCs and ACs, both of which are known to promote neurite outgrowth [29]. In addition, ADSCs produced various growth factors, including BDNF, NGF, GGF, HGF, IGF-1, and VEGF [3,15 –18]. These factors are well known to stimulate axonal outgrowth and enhance the survival and proliferation of SCs [32 –37].
It remains to be clarified whether any of these factors plays a critical role in the effects on SCs and DRG neurons in vitro. However, Zhao et al. have recently shown that the neuroprotective effect of ADSCs is highly dependent on the presence of BDNF and, interestingly, have found that the effects of ADSC-CM are more robust than those of exogenous BDNF alone [40]. These findings suggest that multiple factors produced by ADSCs may act in a synergistic or additive manner to enhance neural regeneration. In this regard, it is noteworthy that the expression of several factors (eg, VEGF and HGF) was much higher in ADSCs than SCs. Addition of ADSCs could create a more favorable extracellular environment for peripheral nerve repair in vivo.
It has been suggested that donor age and the anatomic site of origin influence the behavior of ADSCs. This issue must be addressed to achieve safe and effective application of ADSCs in a clinical setting. Our results showed that ADSCs derived from older mice grew more slowly than those from younger mice under standard culture conditions, which is consistent with previous reports [21,41,42]. The age-related decline in cellular proliferation in vitro could be attributed to intrinsic cell processes such as telomere shortening [43], DNA damage accumulation [44], and oxidative stress [45,46]. However, ADSC cultures of all groups were expandable over 5 passages. In addition, their immunophenotypic features were indistinguishable from each other. More importantly, we found no significant differences among the groups in terms of the beneficial effects on neural cells and all were far superior to 3T3-L1 cells. Thus, the secretion of growth factors by ADSCs seems to be well maintained regardless of donor age and site of origin. A further study is needed to examine whether human ADSCs isolated from diseased or aged donors similarly retain this ability, but our results suggest that ADSCs could be employed as promising candidates for autologous cell therapy in patients with peripheral nerve injury.
We are currently underway in examining whether ADSCs retain the activity promoting peripheral nerve regeneration in vivo after transplantation. In this context, it will be important to identify a way to maximize the neuroprotective/neuroregenerative effects of ADSCs. Pre-differentiation of ADSCs into Schwann-like cells is a promising candidate since it has been shown to increase the production of BDNF [47]. Hypoxic stress and a low serum culture system can be an alternative way to stimulate secretion of these factors [48 –50]. Nevertheless, our current results show that ADSCs have the capacity to produce relatively high levels of various neurotrophic factors even in an undifferentiated state. Indeed, several recent studies have reported the beneficial effects of undifferentiated ADSCs for neuronal regeneration [51 –53]. Future studies will be needed to determine the most effective and practical preparation of ADSCs for peripheral nerve repair in vivo.
Conclusion
To summarize, we demonstrated that ADSCs produced factors that were capable of promoting survival and proliferation of SCs and enhancing neurite outgrowth in DRG neurons in vitro. This supportive function of ADSCs is likely to be maintained regardless of donor age and anatomic site of origin, and may not require complicated manipulation before grafting. Our results would thus provide a basis for autologous cell therapy in cases of peripheral nerve injury.
Footnotes
Acknowledgments
This work was supported by grants-in-aid from the Ministry of Education, Culture, Sports, Science and Technology of Japan (22791727).
Author Disclosure Statement
The authors declare that no competing financial interests exist.