
Abstract
Mesenchymal stems cells (MSCs) are a population of multipotent cells residing in several readily available adult tissue compartments, thus allowing for their ex vivo expansion. To date, therapeutic applications of MSCs have focused on their ability to hone to and support the repair of damaged tissues. More recent evidence suggests that MSCs possess low immunogenicity and a diverse array of immunosuppressive properties. In thisarticle, we will review the basic biology of bone marrow-derived MSCs including their immunoregulatory effects and the putative mechanisms underlying them. We will then present some of the recent clinical applications of MSCs that have leveraged these effects for the treatment of immune-mediated dermatoses such as graft-versus-host disease and systemic lupus erythematosus. Although MSCs offer great therapeutic promise, we will also highlight a number of pertinent challenges that should be overcome before their successful clinical translation.
Introduction
I
More recently, a growing body of data suggests that in addition to their regenerative potential, MSCs also possess immunosuppressive properties. Human MSCs do not express T-cell costimulatory molecules and when used in mixed lymphocyte cultures (MLCs), they suppress lymphocyte proliferation through mechanisms independent of the human leukocyte antigen (HLA) [7,8]. Further, these interactions appear to remain unaltered even after MSC differentiation [9].
Coupled with their regenerative potential, the immunomodulatory properties of human MSCs have garnered support for their clinical translation as a potential therapy in various immunological disorders. This article will explore our current understanding of bone marrow-derived MSCs including their immunomodulatory properties, the mechanisms underlying these effects, and the clinical experience with these cells as therapies for autoimmune dermatoses.
MSC Phenotype
Traditionally, the “stem cell” concept refers to a discrete cell with the biological potential to self-renew, proliferate, and regenerate tissue through creation of a large number of progeny. This term is erroneous when applied to MSCs given that at the single cell level, no isolated clonal MSC has been shown capable of regenerating its native physiological tissue compartment. Although the widespread adoption of the term “MSC” has diluted this argument into one of mostly semantics, understanding the scientific rational beneath it can highlight critical aspects in our current knowledge base regarding MSC biology. Within the bone marrow stroma, MSCs represent a rare heterogeneous population of cells that comprise approximately 0.001%–0.01% of all nucleated cells [10]. Due to their rarity, the study and utilization of MSCs has relied on their successful ex vivo selection and expansion. The most commonly used paradigm involves low-density plating of aspirated mononuclear bone-marrow cells in basal medium supplemented with low quantities of fetal bovine serum [11]. On subcultivation, MSC cultures demonstrate high, but variable expansive potential. Antigenic analysis of ex vivo expanded MSCs shows a common profile, outlined by Bianco and Robey as negative staining for the common hematopoietic markers CD14, CD34, and CD45, but positive staining for CD13, CD29, CD44, CD63, CD90, CD105, CD106, CD146, and CD166 [12]. This profile, unfortunately, is neither unique nor specific to MSCs given that a similar expression pattern is observed for different fibroblast cell lines in other tissue environments [12].
When examined by light microscopy, MSC cultures exhibit a uniform fibroblast-like morphology, but numerous lines of evidence suggest that this population of cells is a composite of several clonal cell types. Work by Muraglia et al. showed that clonal derivatives of aspirated bone marrow stromal cells vary with regard to their differentiation potential along the osteocyte/chondrocyte/adipocyte lineages, and that this potential narrows after sequential passaging [13]. Similarly, although MSCs isolated from bone marrow stroma readily proliferate in vitro, cell cycle studies revealed that the majority of cells (90%) are in the G0/G1 phase of the cell cycle [14]. Colter et al. were the first to show that within the G0/G1 population there exists a subset of small agranular cells that appear to be quiescent [15]. Selection of this recycling stem (RS-1) cell population using the antimetabolite 5-fluorouracil demonstrated that the expansive capacity of different MSC cultures was directly associated with the proportion of RS-1 cells. Further, these cells were antigenically similar to the typical MSC profile, except they lacked CD90 expression, and appeared to be more uncommitted with regard to cell fate, lacking Cbfa-1 and PPAR-γ2 expression, markers associated with osteogenesis and adipogenesis, respectively. Therefore, it appears that RS-1 cells may represent a distinct earlier progenitor cell type within the MSC population. Subsequent studies have identified additional uncommitted MSC subtypes, and equally important, their maintenance ex vivo has also been verified.
Based on these data, it is clear that the concept of a single, ubiquitous MSC isolated from bone marrow stroma lacks substantial support. Rather, MSC cultures isolated for research and clinical use represent a mixture of different progenitors cells spread across a spectrum of differentiation potential and proliferative ability. In this sense, the term “stem cell” may be erroneously applied to most MSC cultures, given the diversity of their cellular constituents. At present, there is no clear or reliable marker that can discriminate the least differentiated cells within the mix, precluding identification of the “true” MSC. However, a number of highly detailed MSC hierarchies have been proposed that outline the putative pathways between the various clonal cell lineages [13,16].
MSC Multipotency
Since the early work by Friedenstein et al., it has become clear that subsets of bone marrow-derived MSCs retain their multipotency when transplanted in vivo. Traditionally, the analysis of MSC differentiation was limited to the mesenchymal trilineage of bone [3,17,18], cartilage [19 –21], and adipose tissues [3,22,23]. However, more recent studies examining the injection of MSCs in utero in sheep or into newborn mice have found engraftment and site-specific differentiation of MSCs into broader tissue types including various muscle subtypes and even neuronal tissue [4,24 –26]. Validation of differentiated phenotypes in these studies consists of a mixture of molecular markers such as lipoprotein lipase for adipocytes [3], cellular markers such as mineralized matrix formation for osteoblasts [17], and functional experiments such as measurement of neurotransmitter release or patch-clamp analysis of neuronal action potentials [27,28]. A thorough summary of the various differentiation stimuli along with the corresponding phenotypic markers they induce can be found in the review by Minguell et al. [16]
MSC Antigenic Profile
Adult human MSCs express moderate levels of the major histocompatibility complex (MHC) class I antigen and intracellular deposits of the MHC class II antigen [29]. Treatment with interferon-gamma (IFN-γ) upregulates expression of both HLA class I and class II antigens in undifferentiated MSCs; however, this effect is altered as MSCs differentiate. After MSCs are grown in media to stimulate adipogenesis, there is a reduction in both HLA antigens before or after IFN-γ stimulation, in particular, class II expression is undetectable. When MSCs undergo chondrogenic and osteogenic differentiation, only HLA class I expression is increased after IFN-γ stimulation, whereas HLA class II expression is reduced, as compared with undifferentiated MSCs [9]. The moderate yet inducible levels of MHC class I antigens suggests that MSCs may evade innate immune responses mediated by natural killer (NK) cells; indeed, this has been recently verified in vitro [30].
Transplantation of donor tissue can elicit a powerful immune response in various manners. Specifically, the indirect pathway of allorecognition, which involves T-cell recognition of donor peptide fragments presented by self-MHC molecules, has been implicated in the pathogenesis of chronic rejection [31]. Paradoxically, the indirect pathway has also been found to suppress immune response through the induction of regulatory T-cells [31]. This latter finding is of immense relevance to the future of clinical transplantations. Further investigations should aim at uncovering the factors that gear the indirect pathway toward tolerance and away from rejection.
Despite also possessing thymic antigens such as CD90, and lymphocyte adhesion molecules such as CD106 (VCAM-1), MSCs do not express T-cell costimulatory molecules B7-1, B7-2, or CD40, even after exogenous stimulation, and, therefore, are not able to stimulate T-cells directly [32 –34]. This last property suggests that MSCs may be transplanted across MHC barriers, including non-HLA-matched individuals.
MSCs and Innate Immunity
A majority of the literature on MSC immunomodulation focuses on the adaptive immune system. However, a few studies have examined the effects of MSCs on players within the innate immune system, namely NK cells and neutrophils. With regard to NK cells, MSCs have been found to be immunosuppressive when grown in MLCs at a ratio greater than 1:10 MSC to NK cells. Under this setting, MSCs are able to inhibit the cytotoxic activity of both resting and interleukin (IL)-2 or IL-15 activated NK cells through either contact-mediated inhibition of activating proteins NKp30 and NKG2D or by the release of PGE2 in transwell studies [30,35,36]. The translation of this and other MSC growth ratio requirements in relation to other human immune cells in vivo is illustrated in Table 1 by using reference values in adults older than 18 years of age assuming an average blood volume of 5 liters [37]. With regard to neutrophils, MSCs have recently been found to play a significant role in dampening their respiratory burst cycle through IL-6 secretion, which, in turn, preserves neutrophil lifespan in critical reserve sites such as the lungs and bone marrow [38].
Assumes immunosuppression achieved at a ratio of 1:10.
MSCs and Adaptive Immunity
MSC effects on antigen presenting cells
When cultured with monocytes, MSCs inhibit the differentiation of dendritic cells and further reduce their potential to activate CD4+ T-cells through the down-regulation of MHC class II molecules [39,40]. Further, when cocultured with mature dendritic cells, MSCs have been found to reduce secretion of the inflammatory cytokines TNF-α, IL-12, and IFN-γ, while promoting the release of immunosuppressive IL-10 [39,41 –43]. Although these effects were observed in vitro, they may translate in vivo as an indirect T-cell suppression pathway, as inhibition of dendritic cell function would abrogate efficient antigen presentation, precluding the clonal expansion of T-cells.
MSC effects on T-cells
When present in MLCs at a ratio of 1:10 or higher, MSCs exhibit an immunosuppressive effect on T-cells, thus resulting in decreased cellular proliferation and IFN-γ production, even in the presence of mitogens [8,9,29,32,41,43,44]. However, when a smaller dilution of MSCs is used in the range of 1:100–1:10,000, enhanced proliferation of lymphocytes is elicited [8,34,45]. This dose-dependent immunosuppressive effect is retained even after MSC differentiation [9]. Concurrent with the general immunosuppression response seen in MLC, specific responses are evident in subtypes of T-cells. CD8+ T-cell mediated cytotoxicity is suppressed if MSCs are introduced early, but not late in the MLC, thus suggesting that their inhibitory ability can only target the activation of CD8+ cytotoxic T-cells [44]. Further, the population of regulatory CD4+CD25+ T-cells known to possess suppressor activity has been shown to increase in MLCs after MSCs are introduced, but only in the absence of mitogens [41,43]. Lastly, when MSCs are cocultured with naïve T-cells exposed to the TH1 promoting signals IL-2, IL-4, and IL-12, a greater proportion of cells develop into TH2 cells with a concomitant reduction in IFN-γ secretion [41]. Overall, MSCs appear to suppress T-cell activation and proliferation nonselectively through multiple mechanisms.
MSC effects on B-cells
Many studies that examined the effects of MSCs on B-cells have shown conflicting results despite their common usage of either Staphylococcal Protein A or pokeweed mitogen to activate B-cells through T-cell-dependent pathways [8,46]. Corcione et al. bypassed this issue by using purified B-cells activated with anti-CD40, anti-IL-2, and anti-IL-4 immunoglobulins before their exposure to MSCs in a transwell setup [47]. Their results indicate that B-cell proliferation and immunoglobulin production are suppressed only if the population of MSCs is at a 1:1 or greater ratio [47]. Although in vitro data tracing the direct interactions between MSCs and B-cells is lacking, this may have little pragmatic impact in vivo, as a majority of B-cell activity is T-cell dependent, thus suggesting that T-cell responses to MSCs may subsequently diminish resultant B-cell responses.
MSC Immunomodulatory Mechanisms
Despite the extensively documented immunosuppressive effects of MSCs, knowledge of their mechanistic underpinnings is partial at best. A recently elucidated contact-mediated mechanism in T-cell suppression appears to involve activation of the programmed death 1 protein and its inhibitory pathway in T-cells via a ligand-dependent pathway on MSCs [46]. A number of soluble factors that are either situation specific or constitutively expressed have also been reported as playing a role in mediating MSC immunosuppression. Three inducible factors that have recently been discovered are indoleamine 2,3-dioxygenase (IDO), inducible nitric oxide synthase (iNOS), and HLA-G5 [48 –50]. In response to IFN-γ, MSCs increase secretion of IDO, which enzymatically degrades tryptophan, an essential amino acid for lymphocyte and NK cell proliferation [35,51]. In a mouse model, iNOS secretion by MSCs has been found to be inducible by IFN-γ alone or in combination with TNF and IL-1, thereby increasing the synthesis of nitric oxide, an innate inhibitor of T-cell activation [48,52]. Lastly, MSC contact has been found to stimulate T-cell secretion of IL-10, which subsequently induces MSC secretion of HLA-G5, a soluble factor known to inhibit T-cell and NK cell proliferation, while increasing expansion of CD25+ regulatory T-cells [49].
In addition to inducible factors, a number of constitutively expressed cytokines have been linked to the immunosuppressive effects of MSCs. Compounds such as hepatic growth factor, transforming growth factor beta 1 (TGF-β1), IL-10, PGE2, and IL-6 have been associated with partial restoration of lymphocyte proliferation when they are selectively inactivated in MLCs by using specific antibodies [8,29,34,39,41,53]. These results, however, are inconsistent when repeated in other experimental series, a discrepancy most likely due to differences in the methodology used to procure and activate relevant immune cells [32,54]. Further, none of these factors were able to completely eliminate MSC induced immunosuppression when targeted alone. This suggests that these factors may act in concert to generate synergistic or additive effects in parallel with contact-dependent pathways. A summary of these pathways is shown in Fig. 1.
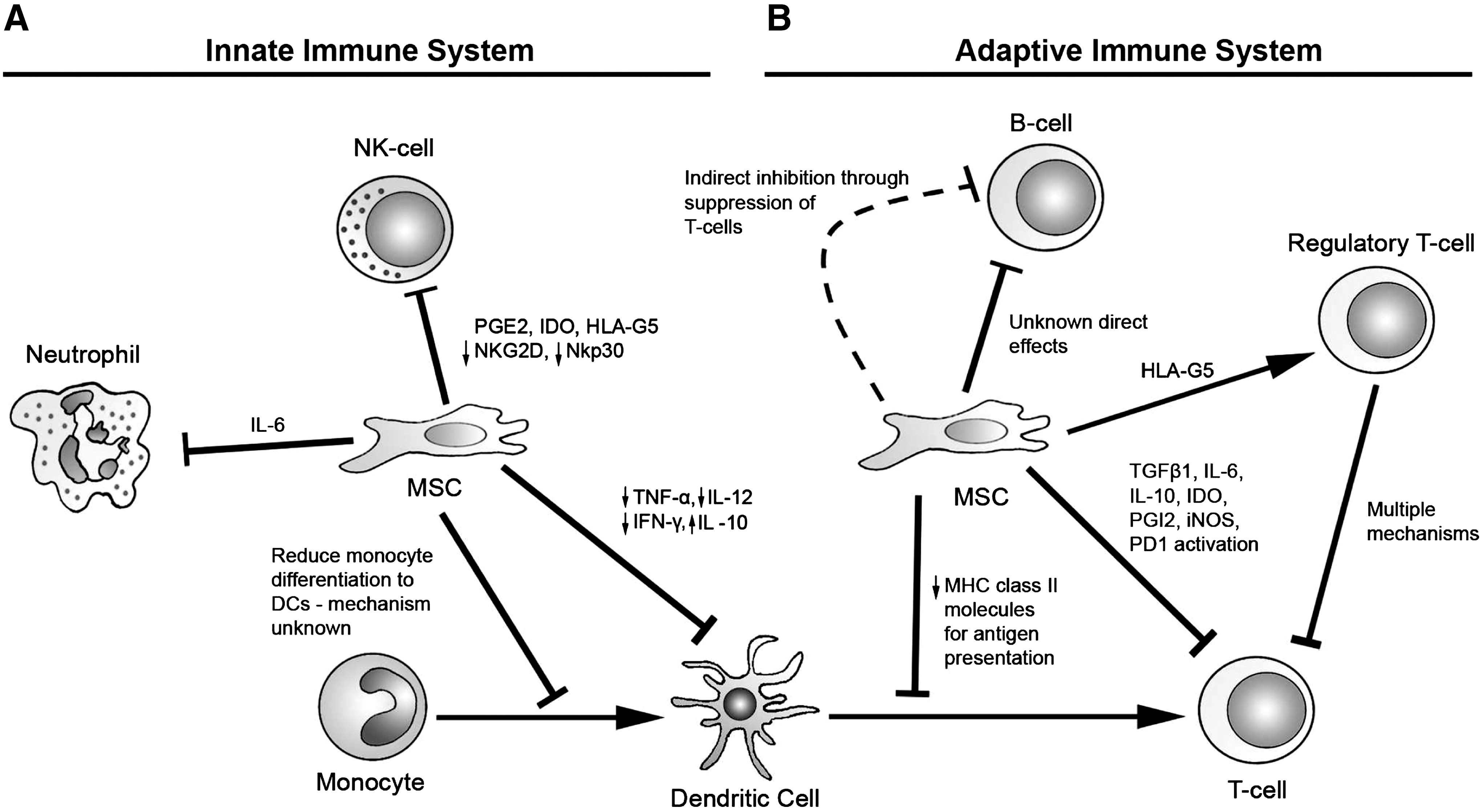
MSC Immunomodulatory mechanisms. MSCs are immunosuppressive against multiple players within the innate and adaptive immune systems.
Limitations of In Vitro Studies
Almost all the studies that have examined the immunosuppressive effects of MSCs have been carried out in vitro. Unfortunately, this raises several issues regarding variability in experimental design as well as in vivo reproducibility. It is critical to note upfront that the documented effects of MSCs in MLCs are heavily dose dependent, thus representing a potential source of bias. As noted by Le Blanc et al., the most common finding associated with MSC introduction into MLCs is a reduction in T-cell proliferation and IFN-γ secretion. However, most of these studies utilize an MSC-to-lymphocyte ratio of 1:10 [39,47,48,53,55], with some even using a 1:1 dilution [9,29,32,34,44]. These ratios are significantly skewed away from physiological and therapeutically practical levels, as a conservative estimate of the proportion of MSCs in bone marrow is less than 1:10,000 [10].
Moreover, as shown in Table 1, the human circulatory system of an average 70 kg adult contains 9.1×109 T-cell lymphocytes. This implies that at least 1.3×107 MSCs per kg would be required for in vivo immunosuppression based on a 1:10 ratio of MSCs. However, most clinical applications dose MSCs at 1–5×106 cells/kg of total body weight, or roughly the equivalent of a 1:100 to 1:20 ratio of MSC to T-cells. At this ratio, in vitro studies have shown MSCs to be immunostimulatory and not immunosuppressive [8,34,45]. Thus, MSC clinical use may instead be limited by their side-effect profile or physical limitations such as pulmonary embolism due to cell aggregation on intravenous injection. Further, many of these studies do not use masterbanked, clonally-derived MSCs, precluding association of any immunological response to a specific cell line. Implications of the former may ultimately be minor in vivo if MSCs are utilized in a setting of immunologically mediated tissue damage, whereby preferential migration of MSCs to sites of tissue injury may allow them to attain a high enough local concentration sufficient to exert their observed in vitro effects [56,57]. Clearly, further studies are required to clarify this critical issue.
Clinical Use of MSCs
Clinical translation of MSC therapy has sought to leverage 3 main processes: regeneration of damaged or defective tissues, providing paracrine support for hematopoietic engraftment, and modulation of host immune responses. This article will focus on the last indication with emphasis on the role of MSC therapy in immunological disorders affecting the skin. Although several studies have utilized animal models to study basic MSC biology, significant differences exist between species with regard to the immunomodulatory mechanisms and the effects that they achieve. For example, when using mouse MSCs, cell-to-cell contact is a prerequisite for immunosuppression that can further induce long-term T-cell anergy, features that are not shared by human MSCs [7,58]. Consequently, pre-clinical animal work may be a poor predictor of human therapeutic responses. Not surprisingly, a majority of human MSC clinical trials have not progressed beyond Phase I and have involved severe disease states. A summary of all relevant clinical studies exploring MSC therapy for immune-mediated dermatologic disorders can be found in Table 1.
Graft Versus Host Disease
One of the first clinical applications of MSC immunomodulatory therapy is for the prophylaxis and treatment of graft versus host disease (GVHD). Occurring in 44%–66% of all allogenic stem cell transplants, acute GVHD is thought to be mediated by donor lymphocytes that are alloreactive to recipient antigens [59 –61]. The skin is the most frequently affected organ, and its involvement correlates with increasing disease grade from a rash that covers less than 50% of total body skin in Grade I disease to generalized erythroderma with bullous formation in Grades III-IV [61]. In contrast to acute GVHD, the pathophysiology of chronic GVHD remains poorly understood. However, similar to the acute form, skin involvement remains the most commonly affected organ with extensive involvement occurring in 30–46% of patients who have undergone transplants [59 –61].
In 2002, Frassoni et al. published a prospective study examining whether MSCs present in bone marrow could be responsible for GVHD incidence differences seen in stem cell transplants utilizing grafts derived from bone marrow versus peripheral blood. Their results showed that patients who had received a hematological stem cell transplant with an expanded MSC infusion after myeloablation had a significant reduction in the incidence of both acute and chronic GVHD at 6 months post-transplantation [62]. Data reported by 3 subsequent phase I/II clinical trials looking at stem cell transplantation after myeloablation for hematologic malignancies echoed similar findings (Table 1) [63 –65]. The mechanism underlying this effect is thought to be MSC-induced tolerance of alloreactive donor T-cells through the previously discussed in vitro pathways. However, this effect may be a double-edged sword based on a recent study by Ning et al. that found higher rates of malignancy relapse in their MSC receiving cohort [64]. A potential explanation may be that since the patients in Ning et al.'s study received, on average, higher MSC doses, the immunosuppressive effect may have additionally blunted the therapeutic graft-versus-leukemia effect.
Beyond GVHD prophylaxis, preliminary studies have also examined the safety and efficacy of using MSCs to treat acute GVHD. In 2004, Le Blanc et al. published the first reported case of successful steroid-resistant GVHD treatment by using MSC infusions [66]. In their report, a 9-year-old boy developed grade IV GVHD after receiving a hematopoietic stem cell transplantation from his mother, was treated with an initial dose of 2×106 MSCs per kg that produced remission, but then developed mild GVHD recurrence 77 days later, thus requiring an additional 1×106 MSCs per kg bolus that elicited a sustained response. The only confounder in their initial report is that the patient's recurrence occurred roughly 30 days after cessation of cyclosporine therapy, which was reinitiated at the recurrence and maintained. Just the fact that there was an initial short-lived response is interesting in itself and calls into question whether true immune tolerance had occurred or whether a transient effect was at play. Mechanistic possibilities include differentiation of MSCs, redistribution of cells outside the bone marrow milieu, or a more limited role for MSCs where they aid solely with the initial engraftment of donor hematopoietic stem cells without contributing any immunomodulatory actions.
Since this initial case report, their group has reported data on 55 patients in an ongoing Phase II clinical trial (Table 2) [67]. Within the cohort, over half of the patients (55%) experienced a complete response, thus recovering fully from the disease. Further, a majority of MSC infusions were across HLA-mismatched donors, thereby supporting the hypothesis that MSCs can be safely transplanted across MHC barriers. However, this claim has not yet been fully validated, as the group was unable to demonstrate or track actual engraftment of the injected MSCs. This limitation again casts doubt on the underlying in vivo immunomodulatory activities of MSCs that one would expect to coincide with sustained engraftment versus a transitory role such as paracrine support. An additional dilemma that remains is determining limited improvement in the 45% of patients who did not respond favorably to therapy. Some possibilities such as variations in treatment procedure and dosing can be readily answered with larger studies. Other prospects such as a lack of standardization in defining the cell types used and their expansion characteristics point to current lax standards. For example, present methodologies typically rely on flow cytometry selection of postexpanded cells for a combination of positive and negative markers, which are by themselves nonspecific, and these cell populations are not assayed for multipotency before injection. Overcoming these limitations by developing a meticulous and consistent working protocol for gathering, assessing, and utilizing MSCs still requires bridging fundamental gaps in our understanding and evaluation of their basic biology.
GVHD denotes graft versus host disease; HLA, human leukocyte antigen; HSC, hematopoietic stem cell; MSCs, mesenchymal stem cell; PBSC, peripheral blood stem cell; SCT, stem cell transplantation; SLE, systemic lupus.
Systemic Sclerosis
Systemic sclerosis (SS) is a debilitating connective-tissue disorder that is characterized by fibrosis of cutaneous and visceral tissue, microvascular pathology, generation of autoantibodies, and prominent lymphocytic infiltration within affected tissue [68]. Although the origin of SS is unknown, the disease shares many clinical features of GVHD, and early observations have led some to conjecture that SS may be a variant of chronic GVHD [69 –71]. This hypothesis has recently gained momentum after allogeneic cells were detected in the peripheral blood and tissue of postgravid SS patients [72,73], but definitive evidence regarding their role in the pathogenesis of SS remains elusive. Currently, effective therapy for SS is lacking, and the disease remains resistant to conventional immunosuppressive regimens with poor prognosis once diffuse involvement occurs.
The demonstrated safety of MSC therapy coupled with its usage in patients with GVHD led Christopeit et al. to explore its utility in the treatment of a single SS patient as accounted within a case report [74]. The patient presented with diffuse cutaneous disease involvement with a modified Rodnan skin score of 25, decreased transcutaneous partial oxygen pressure in both hands, and 6 ulcerations covering 17 cm2 of body surface area. The patient received a single infusion of allogenic MSCs without complication, and at follow-up 6 months later, she showed a reduction in her modified Rodnan skin score to 11, restoration of normal partial oxygen pressure in both hands, and healing of all ulcerations. This preliminary result suggests that MSC therapy may not only deter SS disease progression, but also reverse its course as seen in the restoration of the patient's peripheral circulation. This anecdotal result has spurred a new Phase I/II clinical trial investigating MSC therapy for diffuse SS, with enrollment currently scheduled to be completed by the end of 2011 (NCT00962923).
Systemic Lupus Erythmatosus
Systemic lupus erythmatosus (SLE) is a common yet potentially fatal autoimmune disease characterized by multisystem tissue damage mediated through autoantibody production and disruption of normal B- and T-cell functions. Cutaneous manifestations of SLE are the second most common clinical finding and affect over half of all patients [75]. Traditional medical therapies for SLE have centered on the use of immunosuppressive agents such as cyclophosphamide and mycophenolate mofetil, but within the past decade, therapies such as the anti-CD20 B-cell-depleting drug, rituximab, have emerged to target specific disease processes. Recently, MSCs have come to the forefront as a potential SLE therapy based on the work of Sun et al. In this study, the investigators noticed that within MRL/lpr mice, a common murine model of SLE that displays identical serological and pathological features of the disease [76], bone marrow derived MSCs showed impairment of differentiation potential especially within the osteoblast lineage [77]. The authors hypothesized that MSC dysfunction contributes to generalized dysregulation of precursor lymphocytes within the bone marrow niche, and demonstrated that allo-transplantation of MSCs within MRL/lpr mice significantly reduced levels of autoantibodies, improved renal function, and diminished tissue damage associated with proliferation of CD4+CD25+ regulatory T-cells [77].
Based on their animal data, Sun et al. have transitioned to human clinical trials, specifically focusing on SLE patients who are refractory to prednisone and cyclophosphamide treatment for at least a year (Table 1). All 4 patients met the revised SLE diagnostic criteria established by the American College of Rheumatology, and at the time of treatment had severe tissue involvement such as lupus glomerulonephritis. Each patient was treated with an infusion of MSCs expanded from HLA-disparate family members without complication. Within 6 months of treatment, the SLE disease activity index was halved in all patients [78], and steroid therapy was successfully tapered. After 12 months of follow-up, all patients showed significant restoration of kidney function based on decreased 24-h urine protein and creatinine measurements. Concomitant with these clinical findings was an increase in the population of CD4+CD25+Foxp3+ T-cells found in the patients' peripheral blood [77].
More recently, additional work done by Carrion et al. has examined the efficacy of using autologous derived MSCs to treat patients with SLE [79]. In their phase I pilot study, Carrion et al. enrolled 2 female patients with SLE with skin, gastrointestinal, and musculoskeletal system involvement who did not respond to multiple immunosuppressive regimens spanning over the course of a year. The patients were infused with autologous derived MSCs and followed for 14 weeks. Unfortunately, no clinical improvements were seen with either patient at the end of follow-up based on sequential SLE disease activity indices. Interestingly, both patients still developed significantly elevated levels of CD4+CD25+Foxp3+ cells despite the lack of clinical improvement. The authors postulate that since MSCs and regulatory T-cells have been demonstrated in vitro to be activated by pro-inflammatory cytokines, the patients may not have had severe enough disease involvement. Although this may be a reasonable hypothesis, the authors did not explore the equally likely possibility that the autologous MSCs may have possessed inherent functional deficits such as impaired proliferative ability and earlier onset of senescence, all of which have previously been reported [80,81]. Further, although the authors did confirm that the isolated MSCs demonstrated immunosuppressive activity in MLCs using a 1:1 MSC-to-lymphocyte ratio, their infusion dose of 1×106 cells/kg would achieve a dramatically lower dose in vivo, and based off of typical lab values would only achieve at best only a 1:150 ratio of MSC to lymphocytes in an 80 kg adult.
Based on these 2 pilot studies, experience with using MSC-based therapies for SLE is limited. Apart from the low incidence of procedure-related complications and adverse effects, there are clearly key factors that substantially affect treatment efficacy which need further elucidation, such as the source of MSCs used, differential responses based on patient disease severity, and the role of T-regulatory cells within these patients. At present, there are 2 larger Phase I/II clinical trials examining the usage of MSCs as treatment for refractory SLE and lupus nephritis (NCT00698191 and NCT00659217). Both studies are estimated to be complete by 2012 and may provide additional insights into the clinical utility of MSCs for SLE.
Future Directions and Conclusion
After more than 4 decades since the initial discovery of MSCs, significant insights have been gained at the cellular and molecular level, but large gaps still exist in our fundamental knowledge regarding basic MSC biology. At the forefront of those limitations is our inability to identify and isolate different cellular constituents of an MSC population based on their level of differentiation toward a particular lineage, or rather their “stemness.” Currently, MSC attributes are only population specific, thus making it difficult to reproduce experimental data both in vitro and in vivo by using different MSC cultures. Further characterization of MSCs that will allow separation of cells based on their progenitor status will enable rigorous control over the identity and quality of cells when assaying MSC functionality and their immunomodulatory properties. These measures will allow standardization of the methodologies used to handle and utilize MSCs, thus enabling greater data consistency and extrapolation between studies.
Although these limitations have been a stumbling block for advancing therapeutic applications of MSCs, they have not been complete obstacles. From the current body of literature, it is clear that at least a portion of the immunosuppressive and anti-inflammatory activities of MSCs found in vitro appear to be active when they are used in vivo to treat certain autoimmune diseases. These activities are putatively mediated by a diverse assortment of mechanisms from the release of soluble cytokines to contact-mediated signal transduction pathways. A critical issue that needs to be addressed is to determine which pathways are the most relevant depending on the indication and conditions that MSCs are used, and how they may be potentially governed. For example, the success achieved with the usage of MSCs during acute disease states suggests that their therapeutic effect may stem from selective migration to areas of active tissue damage, whereby they become activated by the disease microenvironment. Understanding the governance of this selectivity may have dramatic implications on not only the spectrum of diseases and injuries that MSC therapy may effectively treat, but also the timing of treatment based on disease severity. Further, unraveling these mechanisms may shed light on whether their dysfunction in native MSCs may be contributory to the disease process itself. The answer may have far reaching implications on not only the efficacy of autologous versus allogeneic sources of MSCs, but also our basic understanding of disease pathogeneses.
Footnotes
Author Disclosure Statement
No competing financial interests exist.