
Abstract
Multipotent mesenchymal stem cells (MSCs) offer great promise for future regenerative and anti-inflammatory therapies. However, there is a lack of methods to quickly and efficiently isolate, characterize, and ex vivo expand desired cell populations for therapeutic purposes. Single markers to identify cell populations have not been characterized; instead, all characterizations rely on panels of functional and phenotypical properties. Glycan epitopes can be used for identifying and isolating specific cell types from heterogeneous populations, on the basis of their cell-type specific expression and prominent cell surface localization. We have now studied in detail the cell surface expression of the blood group i epitope (linear poly-N-acetyllactosamine chain) in umbilical cord blood (UCB)-derived MSCs. We used flow cytometry and mass spectrometric glycan analysis and discovered that linear poly-N-acetyllactosamine structures are expressed in UCB-derived MSCs, but not in cells differentiated from them. We further verified the findings by mass spectrometric glycan analysis. Gene expression analysis indicated that the stem-cell specific expression of the i antigen is determined by β3-N-acetylglucosaminyltransferase 5. The i antigen is a ligand for the galectin family of soluble lectins. We found concomitant cell surface expression of galectin-3, which has been reported to mediate the immunosuppressive effects exerted by MSCs. The i antigen may serve as an endogenous ligand for this immunosuppressive agent in the MSC microenvironment. Based on these findings, we suggest that linear poly-N-acetyllactosamine could be used as a novel UCB-MSC marker either alone or within an array of MSC markers.
Introduction
S
MSCs have been isolated from many adult tissues, such as bone marrow (BM), adipose tissue and skeletal muscle, and also from amniotic fluid and umbilical cord blood (UCB). MSCs from UCB are more primitive and have higher proliferation capacity than MSCs from other sources [7,8]. The immaturity of newborn cells is associated with lower immunogenicity [5,7], and UCB MSCs can, thus, be transplanted across major histocompatibility complex barriers without the need for immunosuppression [4,9].
UCB-derived MSCs are increasingly utilized in clinical trials for degenerative and inflammatory diseases [10]. To fulfil the expectations raised by MSCs for stem cell therapy, several biological questions need to be addressed. There is a growing need for quick and efficient methods to thoroughly characterize MSCs, to distinguish them from other cells, and to control variation within and between different cell preparations. A specific molecular marker for MSCs to be used in the preparation of pure stem cell populations for therapy has not been defined at the moment.
The surface of stem cells, such as all other human cell surfaces, is covered by the glycocalyx formed by glycoproteins and glycolipids. Thus glycans are the first cellular components encountered by approaching cells, pathogens, signaling molecules, and other binders. The cell surface glycans make up cell-type specific signatures. Therefore, glycan epitopes are ideally suited for identifying and isolating specific cell types from heterogeneous populations [11]. Among the most commonly used cell surface antigens to identify human ESCs are the glycosphingolipid antigens SSEA-3 and SSEA-4 [12,13] and keratan sulphate proteoglycans Tra 1-60 and Tra 1-81 [14,15]. We have recently screened the expression of the SSEA-3 and -4 epitopes on UCB-derived stem cells [16] as well as defined the glycan epitope for the Tra antibodies [17].
We have previously characterized the glycome of human MSCs derived from BM [18]. Linear poly-N-acetyllactosamine (poly-LacNAc) structures are enriched in these cells. Poly-N-acetyllactosamines consist of repeating N-acetyllactosamine disaccharide units (LacNAc; Galβ1-4GlcNAc, where Gal: galactose; GlcNAc: N-acetylglucosamine) (Fig. 1A). The LacNAc units can form linear (blood group i type) or branched (blood group I type) chains [19]. Poly-LacNAc structures are ligands for galectins, a large family of mammalian lectins, which have been shown to play an important role in cell adhesion, microbe-host interactions, and modulation of immune responses [20].
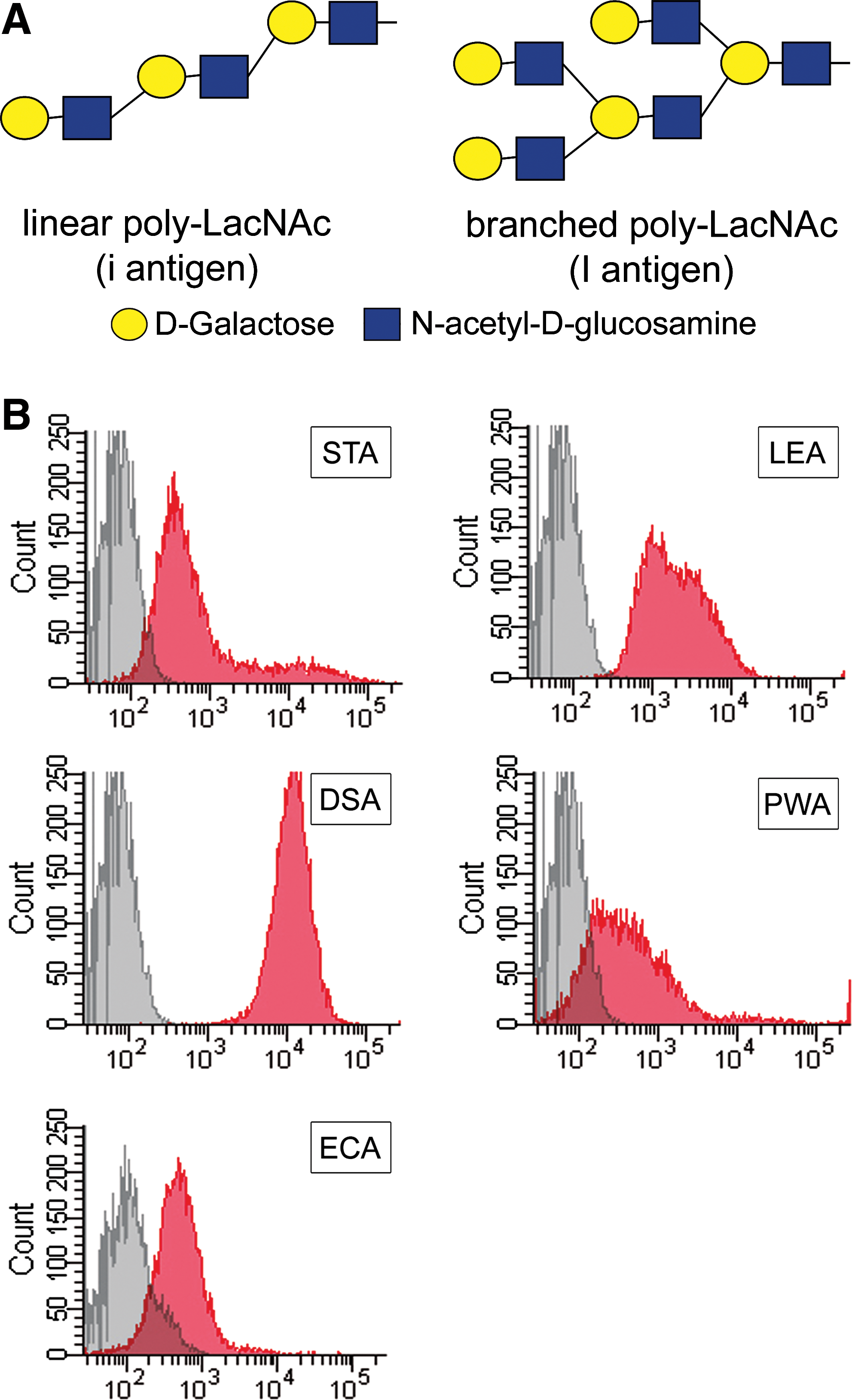
Flow cytometric analysis of lectin binding properties of UCB-MSCs.
We have now further studied the expression of linear poly-LacNAc (blood group i type) structures in UCB-derived MSCs. We demonstrated that linear poly-LacNAc structures are expressed in UCB-MSCs, but not in the cells differentiated from them. Based on these findings, we suggest that linear poly-LacNAc epitopes could be used as a novel marker for MSCs isolated from UCB.
Materials and Methods
UCB-derived MSCs
UCB units were obtained via the Finnish Cord Blood Bank, Finnish Red Cross Blood Service, Helsinki, Finland. Voluntary donors gave informed consent, and the study protocol was accepted by the ethical review boards of the Helsinki University Central Hospital and the Finnish Red Cross Blood Service. Cord blood was collected after normal vaginal delivery as described [21]. MSC lines were prepared as described [22, 23]. UCB-MSCs were characterized by flow cytometry to be negative for CD14, CD34, CD45, and HLA-DR; and positive for CD13, CD29, CD44, CD90, CD105, and HLA-ABC [24]. The cells were shown to be able to differentiate along osteogenic, adipogenic, and chondrogenic lineages as described [16].
All antibody and lectin staining experiments were carried out with cells from 3 separate cord blood units between passages 4 and 6, and the experiments were repeated at least thrice.
Anti-i serum
The human serum was obtained from a patient, and it was serologically shown to contain immunoglobulin class M (IgM) anti-i antibodies. The serum has been routinely used for 15 years in blood group determination of red blood cells for clinical use, as well as in family studies in several families expressing the genetical I negative phenotype (of noncongenital cataract type found in Finns) [25]. The human serum used as a control serum was shown not to contain antibodies against the i blood group antigen.
Lectin and antibody stainings and flow cytometry
For lectin stainings, fixed cells were stained with fluorescein isothiocyanate (FITC)-conjugated Solanum tuberosum agglutinin (STA), Lycopersicon esculentum agglutinin (LEA), Datura stramonium agglutinin (DSA), and Phytolacca Americana agglutinin (PWA) lectins (all from EY Laboratories, Inc., San Mateo, CA), in 0.3% bovine serum albumin in phosphate-buffered saline using the following concentrations: STA 4 μg/100 μL, LEA 4 μg/100 μL, DSA 0.5 μg/100 μL, and PWA 4 μg/100 μL. Biotin conjugated Erythrina cristagalli agglutinin (ECA) lectin (EY Laboratories, Inc.) was used in the concentration of 2 μg/100 μL followed by FITC-conjugated streptavidin (eBioscience, Inc., San Diego, CA). In fact, 5 mM N,N′,N′′ triacetylchitotriose (Sigma Aldrich, St. Louis, MO) was used to inhibit lectin staining.
For antibody stainings, the cells were incubated with galectin-3 monoclonal antibody (R&D Systems Inc., Minneapolis, MN) followed by goat anti-mouse Alexa 488 (Molecular Probes, Invitrogen by Life Technologies, Carlsbad, CA), or 10% anti-i serum or control serum followed by PE-conjugated anti-human IgM (Rockland Immunochemicals, Inc., Gilbertsville, PA). The stained cells were analyzed with BD FACSAria flow cytometer (BD Biosciences, San Jose, CA). The results were analysed with BD FACSDiva™ Flow Cytometry Software software version 5.0.2 (BD Biosciences).
Endo-β-galactosidase assay
About 3×105 UCB-MSCs were fixed with paraformaldehyde, washed, and incubated with 100 mU of Escherichia freundii endo-β-galactosidase (Seikagaku Biobusiness Corporation, Tokyo, Japan) in sodium phosphate buffer pH 5.8 for 2.5 h at 37°C.
mRNA expression analysis
Total RNA from MSCs, adipogenically differentiated MSCs, and osteogenically differentiated MSCs for microarray was isolated using RNase Mini Kit (Qiagen, Hilden, Germany). Sample labeling and hybridization were carried out at the Finnish DNA Microarray Center at Turku Centre for Biotechnology, Turku, Finland. Samples were prepared using One-Cycle Target Labeling protocol and hybridized to GeneChip® Human Genome U133 Plus 2.0 Array (Affymetrix, Santa Clara, CA). AffyReader was used to extract gene expression measures. The Affymetrix probeset expression data of transcripts were newly clustered to represent genes by recombining the probes that represented the same gene using GeneChip library files (Custom CDF, version 10) with esembl gene ID [26,27]. All samples were normalized using robust-multiarray (RMA) Average background adjustment for intensities, quantile normalization, and median-polish summarization [28]. The RMA normalization was implemented using Bioconductor R, package affy. Values were transformed into log2.
Mass spectrometric analysis of cell surface glycans and proteins
Mass spectrometric analyses of proteins were performed with liquid chromatography–mass spectrometry (LC-MS) of tryptic peptides digested from intact proteins essentially as previously described [29]. Protein linked N-glycans were first released by PNGase F (Sigma Aldrich), purified by C18 ZipTip (Millipore, Billerica, MA), and reduced by using NaBH4 in 10 mM NaOH. The N-glycan alditols were desalted by using graphitized carbon pipette tips (TopTip, Glygen, Columbia) and permethylated by a solid-phase spin-column technique essentially as described [30].
Both peptides and permethylated glycan alditols were loaded to a reversed-phase precolumn (NanoEase Atlantis dC18, 180 μm ×23.5 mm; Waters, Milford, MA) and separated in a reversed-phase analytical column (PepMap 100, 75 μm ×150 mm; Dionex Corporation, Sunnyvale, CA) with a linear gradient of acetonitrile. Ultimate 3000 LC instrument (Dionex Corporation) was operated in a nano scale. Eluted peptides or glycans were introduced to linear trap quadrupole Orbitrap XL mass spectrometer (Thermo Fisher Scientific, Inc., Waltham, MA) via an electrospray chip interface (NanoMate Triversa, Advion Biosciences Inc., Ithaca, NY) in the positive-ion mode.
Data from MS measurements were processed with Mascot Distiller (Matrix Science Ltd., London, UK, version 2.2.1.0) via Mascot Daemon Client (Matrix Science Ltd., version 2.2.2). Protein identifications were on Mascot Server (Matrix Science Ltd., version 2.2.04) against the human proteome in UniProtKB database (Release 2010_09) [31]. Glycan compositions were matched against MS and MS/MS data using GlycomeDB database (
Results
The i-epitope is expressed by UCB-MSCs
We have previously reported that MSCs derived from BM express linear poly-LacNAc structures on their surfaces [18]. To determine whether poly-LacNAc structures are also expressed by MSCs derived from UCB, we stained UCB-MSCs with various plant lectins. STA, LEA, and DSA recognize mainly linear poly-LacNAc structures (ie, blood group i antigens). Mitogen PWA recognizes mainly branched poly-LacNAc structures (ie, blood group I antigens), and ECA recognizes terminal LacNAc structures. Flow cytometric analysis of the lectin stained cells revealed that UCB-MSCs express linear poly-LacNAc structures in great amounts (Fig. 1B). LEA and DSA stained 100% of the cells and STA 98%, compared with unstained control cells. UCB-MSCs were also shown to contain branched poly-LacNAc structures to some extent, as PWA stained 77% of the cells. In fact, 80% of the cells stained positive for terminal LacNAc structures with ECA. To verify the specificity of the lectin stainings, the binding of STA, DSA, LEA, and PWA to UCB-MSC was shown to be inhibited by chitotriose (GlcNAc3) (data not shown).
To obtain more specific evidence on the poly-LacNAc expression on UCB-MSCs, we stained the cells with human serum containing anti-i IgM. Flow cytometric analysis revealed that the anti-i serum stained approximately 60% of the undifferentiated UCB-MSCs (Fig. 2A). Nonspecific control serum did not stain the cells. The experiments were repeated 5 times with 3 individual UCB-MSC lines, and 42%–98% of the UCB-MSCs stained positive with the anti-i serum (Fig. 2B). To confirm that the anti-i serum recognizes linear poly-LacNAc structures, we treated UCB-MSCs with endo-β-galactosidase, an enzyme that hydrolyses internal β-galactosidic linkages in oligosaccharides belonging to the linear poly-N-acetyllactosamine series with the common structure GlcNAcβ1-3Galβ1-4GlcNAc/Glc. Cells that had been originally 98% positive for the i-antigen in FACS analysis stained only 48% positive after incubation with endo-β-galactosidase (Fig. 2C), thus indicating the disappearance of linear poly-LacNAc structures.
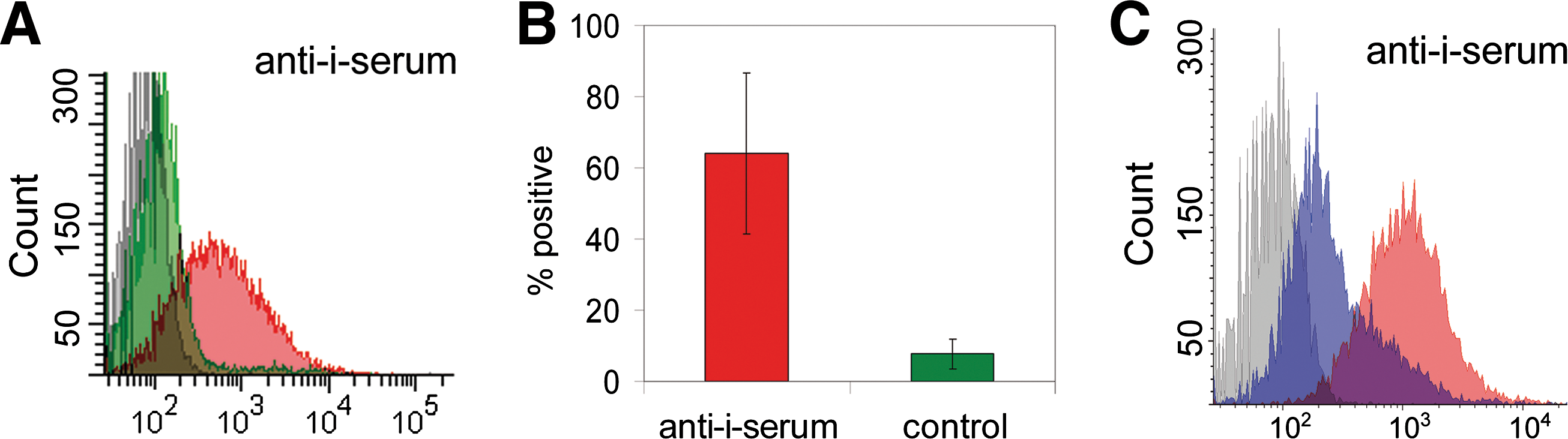
Analysis of UCB-MSCs with human serum containing anti-i immunoglobulin class M.
The mononuclear cell fraction from UCB was stained with anti-i serum to see whether the expression of the i-antigen is specific for MSCs in the context of UCB. Anti-i serum stained most of the UCB T-cells (CD3+) and B-cells (CD20+), and a subpopulation of NK-cells (CD56+), but not monocytes (CD14+) or CD34 positive cells (hematopoietic progenitors) (data not shown).
Cell differentiation abolishes poly-LacNAc from UCB-MSCs
MSCs from cord blood and cells were differentiated from them either along adipogenic or osteogenic lineages that were stained with human anti-i serum, to study whether the expression of the linear poly-LacNAc (i blood group antigen) can serve as a UCB-MSC marker. The anti-i serum strongly stained the UCB-MSCs but not the adipogenic or osteogenic cells (Fig. 3). The osteogenic cells were only 6%–16% positive for anti-i serum staining. Similarly, 10%–15% of the cells stained positive with the control serum. The adipogenic cells were 3%–18% positive, whereas 13%–14% of the cells stained positive with the control serum. These experiments were repeated twice with 3 individual UCB-MSC-lines.
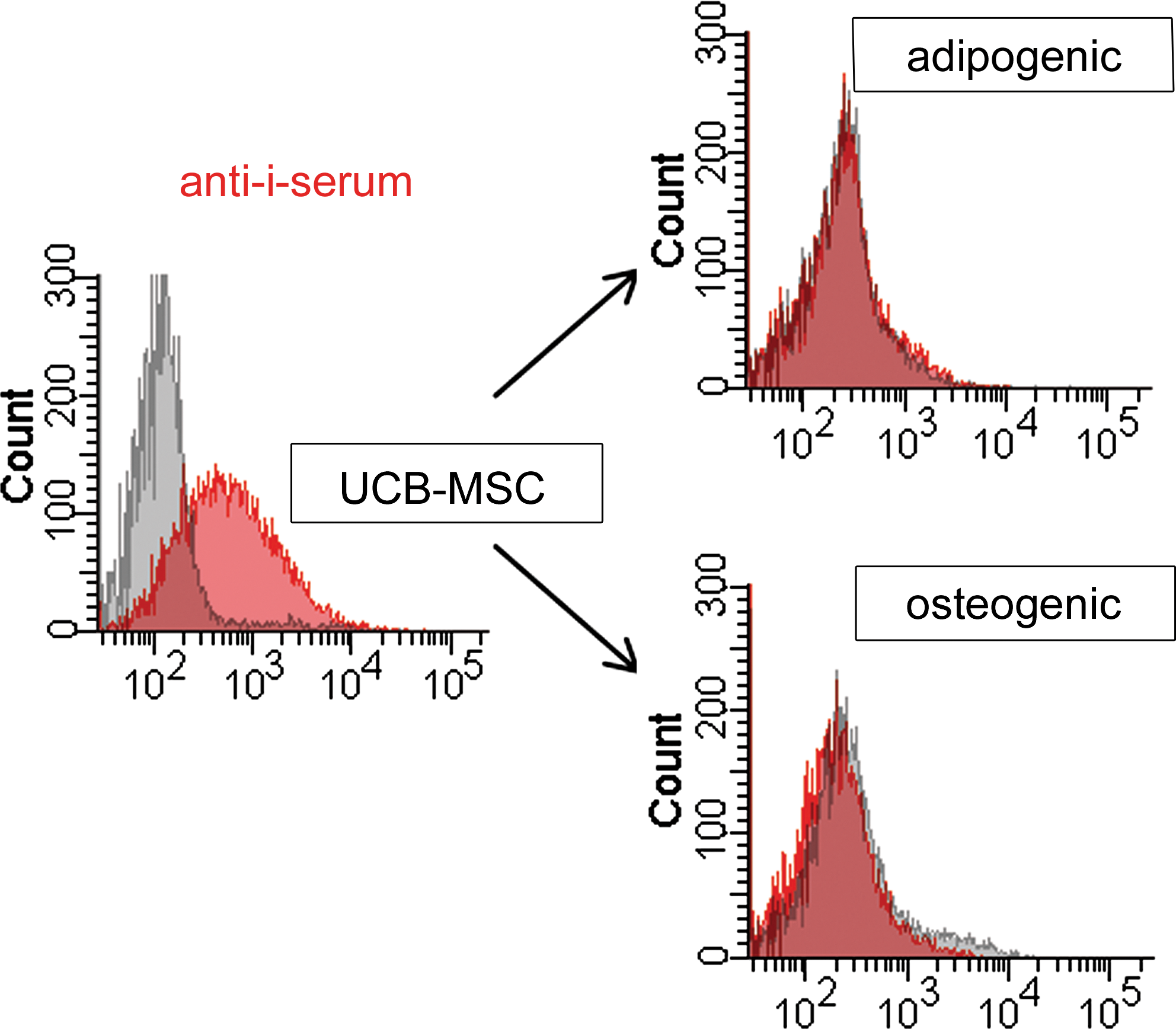
Characterization of the i blood group antigen in cells differentiated from UCB-MSCs. Flow cytometric analysis showing cells stained with anti-i serum (red) or control serum (gray). UCB-MSCs and cells differentiated from them along adipogenic or osteogenic lineages are shown. Color images available online at
The i blood group antigen is displayed by N-glycans on UCB-MSC surface
Cell surface proteins were biotinylated on intact adherent UCB-MSCs, captured on streptavidin, and N-glycans were released from them to verify the linear poly-LacNac structure by mass spectrometric analysis. Permethylated and reduced cell surface N-glycans were first separated by nanoscale LC and subsequently analyzed by high-resolution MS using an electrospray source. The 2-dimensional LC-MS plot showed a wide range of different glycan ions derived from the cell surface proteins of UCB-MSC (Fig. 4A). In the glycan profile derived from the LC-MS/MS data, we could observe 67 different N-glycan compositions from the surface of these cells (data not shown). The glycan compositions can represent multiple different isomeric glycan structures. For example, the most commonly occurring isomer of the glycan composition S1H6N5F1 (S=N-acetylneuraminic acid, ie, sialic acid, H=hexose, N=N-acetylhexosamine, F=deoxyhexose, ie, fucose) is a typical triantennary complex-type N-glycan. Additional diversity is introduced by fucose, which can be either linked to the core GlcNAc or to 1 of the antennae. We observed differential LC retention times for this glycan composition, and showed that among the glycans eluting at 19.5 min there was a S1H6N5F1 isomer with linear di-LacNAc epitope (Fig. 4B). The fragmentation spectrum of this specific glycan precursor included a fragment representing fucose linked to the chitobiose core, as well as the di-LacNAc (poly-N-acetyllactosamine) portion either with or without the sialic acid (Fig. 4C).

Mass spectrometric analysis of linear poly-LacNAc on the UCB-MSC surface.
Specific glycosyltransferases involved in the synthesis of linear poly-LacNAc structures are expressed in UCB-MSCs
An important factor in determining the nature of the glycan structures expressed by a cell is the expression of specific glycosyltransferases that synthesize the glycans. Therefore, we studied the mRNA expression of the glycosyltransferases capable of making linear poly-LacNAc structures in UCB-MSCs (Fig. 5). Poly-N-acetyllactosamines occur on specific types of glycan backbones: in glycolipids on neolactoseries glycosphingolipids [33], in glycoproteins on core 2 type O-glycans [34,35], and on N-glycans, especially on the β1,6-branch [34,36,37]. The biosynthesis of neolactoseries glycosphingolipids is initiated by β1,3-GlcNAc-transferase 5 (B3GNT5) [38 –40]. Core 2 O-glycan synthesis is initiated by the core 2 β1,6-GlcNAc-transferase (C2GNT) [41]. The synthesis of the β1,6-branch of tri- and tetra-antennary N-glycans is under the control of β1,6-GlcNAc-transferase V (MGAT5) [42]. Of these glycosyltransferases, especially B3GNT5 was highly expressed in UCB-MSCs and also upregulated in stem cells when compared with cells differentiated from them (Fig. 5A). The elongation of poly-N-acetyllactosamines occurs further by the sequential addition of β1,4-linked galactose and β1,3-linked N-acetylglucosamine. Of the β1,4-galactosyltransferase family (B4GALTs) [43,44], especially B4GALT2, -3, -4, and -5 were expressed in UCB-MSCs (Fig. 5B). Of the β1,3-GlcNAc-transferases (B3GNTs) capable of poly-N-acetyllactosamine elongation, B3GNT1 and -5 were markedly expressed in UCB-MSCs (Fig. 5C) [39,45 –49]. B3GNT1 is also called iGnT and poly-N-acetyllactosamine extension enzyme, and it is highly likely to be involved in the synthesis of linear poly-LacNAc synthesis.
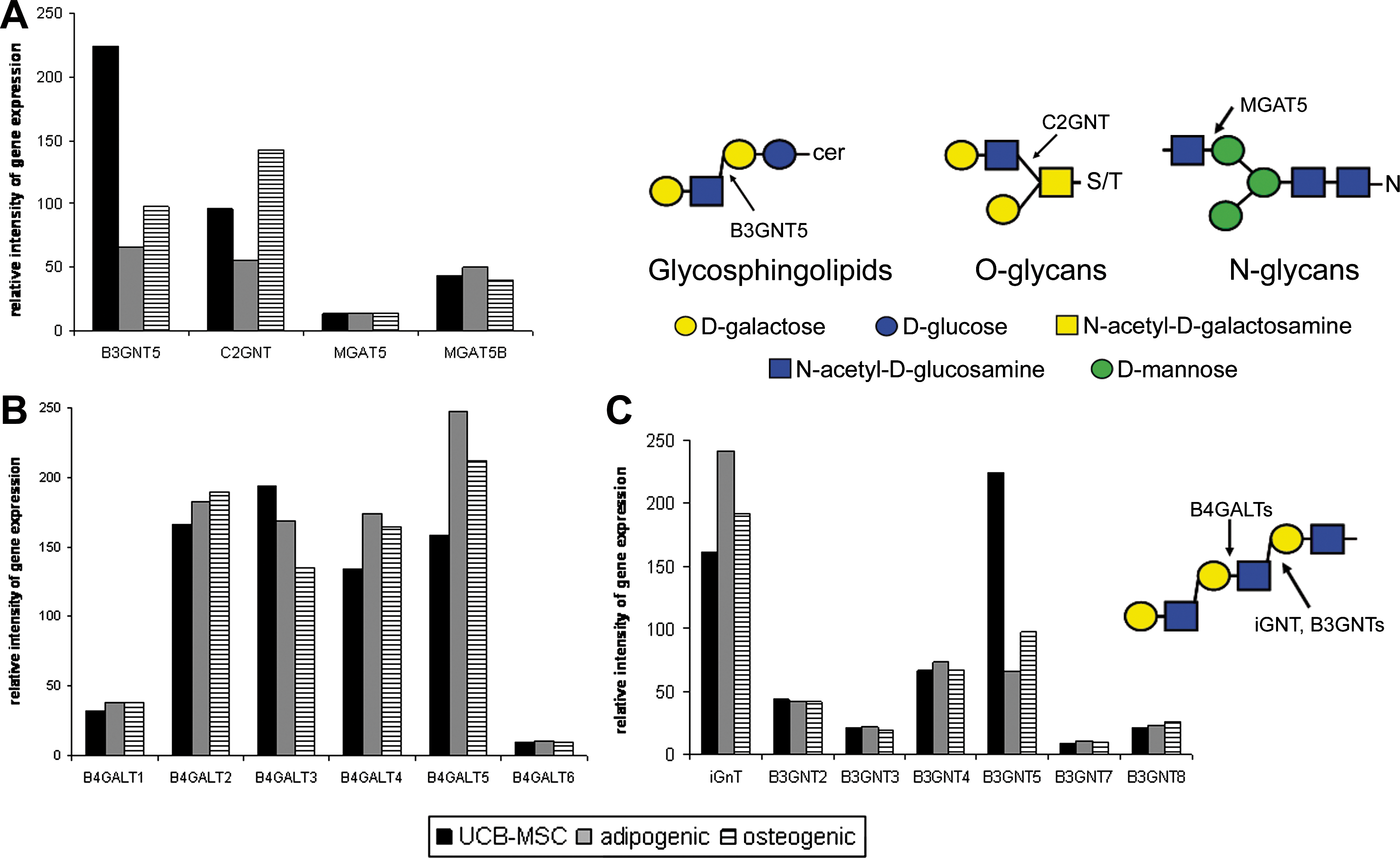
mRNA expression analysis of glycosyltransferases involved in linear poly-LacNAc synthesis. Gene expression analysis of
The poly-N-acetyllactosamine binding lectin galectin-3 is expressed on UCB-MSC surface
A family of mammalian lectins, galectins, has been shown to bind LacNAc-containing glycans. Galectin-3 has been reported to bind to the nonreducing terminal as well as internal LacNAc structures, and its affinity increases with multiple LacNAc units [20,50]. We tested whether UCB-MSCs express galectin-3. The mRNA expression of galectin-3 was analyzed using in-house generated gene expression microarray data from UCB-MSCs [16], where notable expression of the galectin-3 gene was observed (Fig. 6A). The expression of the galectin-3 protein was verified by mass spectrometric proteome analysis of UCB-MSCs, where 3 different tryptic peptides from human galectin-3 were identified (Fig. 6B). The cell surface localization of galectin-3 was confirmed by flow cytometry. In fact, 80% of the UCB-MSCs were positive when stained with an antibody recognizing galectin-3 (Fig. 6C).
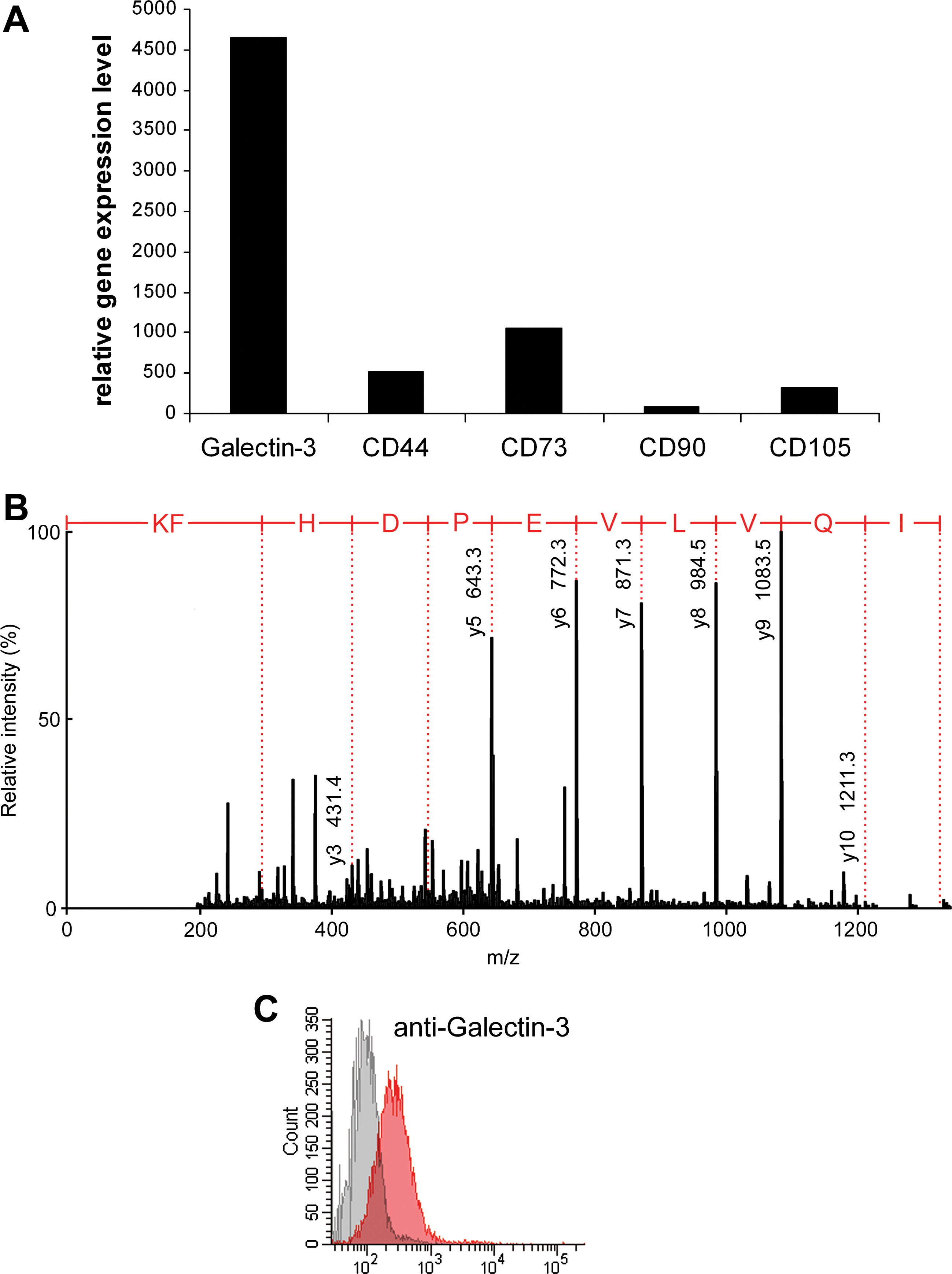
Analysis of galectin-3 expression of UCB-MSCs.
Discussion
In this study, we report a detailed characterization of blood group i antigenic glycan structures in UCB-derived MSCs. We performed complementary analysis using MS, flow cytometry, and gene expression microarray to evaluate the presence of the i antigen on these cells. We found that the i antigen is expressed in UCB-MSCs, but not in the cells differentiated from them. We also discovered that the expression of the i antigen is dependent on a specific set of glycosyltransferases. Further, we were able to show that UCB-MSCs also express galectin-3, a major immunomodulatory lectin binding to the i antigen, on their surface.
MSCs show great potential in future medical therapies for the regeneration of damaged tissues such as bone, cartilage, tendon, and ligament, as well as for inflammatory disorders. They have 4 major advantages: MSCs secrete biologically important molecules, express specific receptors, can be genetically manipulated, and are susceptible to molecules that modify their natural behavior [51]. MSCs have typically been isolated from their original source (eg, BM or UCB) by attachment to tissue culture plastic. In many cases, this is ineffective, and the result is a mixture of MSC populations with different functional characteristics and contaminating adherent cells such as fibroblasts. For therapeutic purposes, there is a clear need to more efficiently isolate and characterize these cells and cell populations.
Expression of cell surface glycans is dependent on the cell type and cellular status. Therefore, glycan epitopes may potentially serve as valuable markers for distinct cell populations. Typical glycosylation features of BM-derived MSCs include poly-N-acetyllactosamines, α2,3-sialylated N-glycans, the sialyl Lewis x epitope, and the globoseries glycosphingolipid antigen SSEA-4 [18]. UCB-MSCs also express SSEA-4 [16], but the glycosylation of MSCs from other sources than BM is scarcely studied.
The blood group i antigen (consisting of linear poly-LacNAc, Fig. 1A) is a cell surface antigen known to be expressed by red blood cells of fetal and UCB origin. After birth, the quantity of I antigens (branched poly-LacNAc, Fig. 1A) on red blood cells starts to gradually increase, whereas the i antigen disappears [52,53]. This increase is due to developmental regulation of the gene encoding the enzyme, which adds branches into the i antigen (N-acetyllactosaminide β-1,6-N-acetylglucosaminyltransferase, ie, GCNT2, ie, I-branching enzyme) [54]. The normal adult Ii status is reached in about 18 months of life. In adults, the expression of the i antigen on red blood cells defines a rare blood group. The expression of the i antigen is developmentally regulated in the course of early embryonic development and in embryonic carcinoma cells [12]. However, the expression of the i and I antigens in other tissues does not correlate with the red blood cell phenotype. The blood group i antigen has been reported to be present in a variety of proliferative (fetal) human tissues and absent from quiescent tissues (adult liver, kidney, etc.) [55].
The expression of the i antigen is influenced by the expression levels of glycosyltransferases synthesizing both the underlying backbone structures and the poly-LacNAc chain itself. Poly-N-acetyllactosamines typically occur in specific types of N-linked, O-linked, and glycosphingolipid glycans. GlcNAc-transferase V coded by the MGAT5 gene initiates the β1,6-linked branch of tri- and tetra-antennary N-glycans, and MGAT5 gene expression has been shown to determine poly-N-acetyllactosamine expression in several models [34,36,37,42]. Gene expression analysis revealed that MGAT5 and MGAT5b were not expressed by UCB-MSCs. In the MS analysis, we saw poly-LacNAc extensions on biantennary N-glycans. Based on these observations, it seems that the expression of the i antigen on UCB-MSCs is independent of MGAT5.
In O-glycans, poly-LacNAc occurs as an elongation of the so-called core 2 structure synthesized by the core 2 N-acetylglucosaminyltransferase, C2GNT [41]. C2GNT expression was low in UCB-MSCs, and not enriched in stem cells as compared with differentiated cells. C2GNT is, therefore, unlikely to be responsible for the UCB-MSC specific expression of the i antigen.
Glycosphingolipids carrying poly-LacNAc belong to the neolactoseries group, the biosynthesis of which is initiated by β3-N-acetylglucosaminyltransferase 5 (B3GNT5) [38 –40]. B3GNT5 was expressed at high levels by UCB-MSCs, and its expression was particularly enriched in stem cells. B3GNT5 is also able to elongate poly-LacNAc chains. Poly-LacNAc elongation occurs by the alternating action of β3-N-acetylglucosaminyltransferases and β4-galactosyltransferases. Of the putative poly-LacNAc elongating β3-N-acetylglucosaminyltransferases, B3GNT5 and iGNT were expressed by UCB-MSCs. The β4-galactosyltransferases expressed by UCB-MSCs were B4GALT2, -3, -4, and -5, which were expressed by comparable levels also in differentiating cells and, therefore, are not likely to determine the stem-cell specific expression of the i antigen.
Our gene expression data suggest that the i antigen expression in UCB-MSCs is under the control of B3GNT5, either as the initiator of neolactoseries glycosphingolipid biosynthesis, or as a poly-N-acetyllactosamine elongation enzyme. In the latter case, the i antigen may be carried by either N-glycans, O-glycans, or glycosphingolipids. Our structural data indicate that the i-antigen related structures are present at least on N-glycans in UCB-MSCs. The O-linked glycans and glycosphingolipids of UCB-MSCs warrant further study.
Cell surface glycans exert their biological effect by binding to their cognate receptors, lectins that occur either in soluble or transmembrane form. Galectins are a family of animal lectins that bind N-acetyllactosamine-containing glycans. Galectin-3 is evolutionarily highly conserved, and it is found both intra- and extracellularly. It is a soluble protein that is secreted to the extracellular space. Once secreted, it tethers to its ligands on the same or adjacent cell. Galectin-3 binds the nonreducing terminal as well as internal LacNAc units. The affinity increases with multiple LacNAc units: the i antigen observed in N-glycans currently characterized on the MSC surface is a high-affinity ligand [50]. Galectin-3 is involved in various cellular processes such as cell growth, adhesion, differentiation, angiogenesis, and apoptosis, and its expression has also been correlated with cancer aggressiveness and metastasis. Notably, galectin-1 and galectin-3 have been suggested to be responsible for the immunosuppressive properties of MSCs, although the exact mechanisms by which galectins suppress T-cell function remain unknown [6].
We observed notable mRNA expression of galectin-3 on UCB-MSCs. We were also able to detect galectin-3 on UCB-MSC surface by MS and flow cytometry. Galectins are known to selectively cross-link their ligands on the cell surface to form lectin-carbohydrate lattices. The poly-LacNAc ligands on the UCB-MSC surface can tether galectin-3 on the cell surface and thereby participate in galectin-mediated signaling.
UCB is likely to be an important source of therapeutic cells in the future. Therefore, there is an increasing need for markers that define specific UCB cell populations. Based on our findings, the expression of the i antigen and consequent sequestration of the immunomodulatory lectin galectin-3 on the cell surface can be considered characteristic of proliferating UCB-MSCs. Within the mononuclear cell fraction of UCB, anti-i serum stained most of the T-cells and B-cells, and a subpopulation of NK-cells, but not monocytes or CD34 positive cells. Cells of the monocyte/macrophage lineage are common contaminants in MSC cultures, whereas cells of the lymphoid lineage do not adhere to plastic and are, thus, eliminated during the initial phase of the culture. We suggest that the i antigen could be used as a novel marker for MSCs derived from UCB, either alone or within an array of other MSC markers, thus improving the quality and efficiency of the isolation and characterization of UCB-MSCs for therapeutic purposes.
Footnotes
Acknowledgments
The authors would like to thank the Finnish Red Cross Blood Service Cord Blood Bank, acknowledge Teija Kupari, Lotta Andersson, Birgitta Rantala, and Sirkka Hirschovits-Gerz for excellent technical assistance, and are grateful to Adjunct Professor Jukka Partanen and Dr. Jari Natunen for valuable comments and careful review of this article. This work was supported by the Finnish Funding Agency for Technology and Innovation (TEKES); the EVO Medical Research Fund of Finnish Red Cross Blood Service, and the Finnish Glycoscience Graduate School.
Author Disclosure Statement
The authors declare potentially competing interests, as Tero Satomaa and Juhani Saarinen are shareholders of Glykos Finland Ltd.