
Abstract
Both ageing and diabetes are associated with reduced numbers and functional viability of mesenchymal stem cells (MSCs) in vivo which in turn lead to degenerative pathologies of the musculoskeletal system. The overall aim of this study was to elucidate the effects of age and raised glucose levels on the proliferation and self-renewal of rat nonadherent bone marrow MSCs (Na-BM-MSCs) in suspension cultures. MSC cultures isolated from 3- and 12-month-old rats were maintained using the “pour-off” method for up to 14 days in media containing different glucose levels and the phenotype, growth characteristics, colony forming unit-fibroblastic (CFU-f) numbers, and pluripotency characteristics of these cells were determined. This study indicates that rat adult bone marrow harbors pluripotent Na-BM-MSCs that seem to be unaffected by ageing during in vitro expansion. The Na-BM-MSCs express the pluripotency markers Oct4, Sox2, and Nanog. It was found that culture in high-glucose-containing medium had a negative effect on colony formation and differentiation. In contrast to classical MSC cultures, the generation of colonies by Na-BM-MSCs in suspension culture was not reduced in the older animals. The Na-BM-MSCs were found to express the pluripotency markers Oct4, Sox2, and Nanog, suggesting a more primitive stage of differentiation as compared with adherent MSCs. These data indicate that rat adult bone marrow harbors a population of pluripotent Na-BM-MSCs that appear to be relatively unaffected by ageing during in vitro expansion in suspension.
Introduction
M
A number of preclinical and clinical studies have investigated the possible therapeutic applications of MSCs. Some of these therapies address mostly elderly people with, for example, degenerative cartilage defects or stroke, all of which are associated with an increased incidence of type II diabetes. This disease develops from insulin resistance, which leads to hyperglycemia [12]. Thus, we were interested in the quality of MSCs expanded in suspension cultures from older donors using different glucose concentrations during cultivation. Previous work on classical plastic-adherent MSCs showed that glucose can induce senescence and impairs the osteogenic differentiation potential [13]. Alternative forms of MSCs have been described and were termed nonadherent bone marrow MSCs (Na-BM-MSCs) [14,15]. These cells show similar characteristics to MSCs, having a very broad differentiation potential in vitro and in vivo. In addition, these cells have been shown to repopulate the hematopoietic system even in an allogeneic setting without signs of cell rejection [14].
We have shown in in vivo and in vitro studies that raised glucose levels impair MSC quality [13]. Rat derived MSCs were shown to proliferate more rapidly in low-glucose compared with high-glucose media [16]. Studies on human MSCs also showed increased senescence, as well as decreased proliferation and reduced colony forming unit-fibroblastic (CFU-f) numbers in high-glucose media [17].
Materials and Methods
Pour-off culture of bone marrow adult stem cells
Whole bone marrow containing a mixed population of adult stem cells was derived from Wistar rats and expanded in Dulbecco's modified Eagle's minimal essential medium (low- or high-glucose DMEM; Perbio) containing 10% fetal calf serum (Invitrogen) and 10−8 M dexamethasone (Sigma). The cells (2×106/55 cm2) were cultured in Petri dishes in 5 mL DMEM. The nonadherent cells in the supernatants were counted using Trypan blue staining and transferred daily into a new Petri dish. The left-over adherent cells were analyzed using the CFU-f assay every day during the 14 days of cell transfer (pour-offs).
CFU-f assay
Fresh or cultivated bone marrow cells were analyzed using the CFU-f analysis. For pour-off cultures, the nonadherent bone marrow fraction was removed as described previously and the remaining adherent cells were further cultured as in a traditional CFU-f assay. Cells were incubated with DMEM (high or low glucose) for a further 14 days at which point they were fixed in ethanol, sequentially stained for alkaline phosphatase and methylene blue, and photographed. They were then analyzed for colony number as previously described using Bioimage software [18].
Colony analysis
Colony numbers were assessed using the method of Dobson et al. [19]. Briefly, the acquired digital images were imported into “Photoshop,” colony irregularities smoothed out, and converted into a 256-level grayscale format. The grayscale images were then imported into Bioimage “Intelligent Quantifier” and colony number and colony size were calculated.
Immunohistology
Cells were fixed with 4% paraformaldehyde, and washed and permeabilized with 0.5% Triton X-100 in phosphate-buffered saline (PBS). Nuclei were stained with 4′,6-diamidino-2-phenyindole diacetate. Cells were blocked with 10% donkey serum and incubated for 1 h at 37°C with primary antibodies in PBS (1% donkey serum). After washing, cells were incubated with fluorescence-labeled secondary antibodies (Cy2 & Cy3 1:750; Jackson ImmunoResearch Laboratory) at 37°C for 1 h. Microscopical analysis was performed with a fluorescence microscope (Axio Observer; Zeiss). The following antibodies were used: Nanog (1:1,000; Abcam), Oct4 (1:250; Abcam), and Sox2 (1:50; Santa Cruz).
Alamar blue assay
MSCs were seeded at concentrations between 2,500 and 80,000 per well in 24-well multiwall plates. Fifty microliters of alamar blue was added to each well and the plates were incubated at 37°C for different time intervals. Cell number was then determined by analyzing the supernatants spectrophotometrically at 570 and 600 nm at different time points.
Statistical analysis
Values are expressed as mean±standard deviation. The statistical significance of differences among experimental groups was evaluated by one-way ANOVA and multiple comparisons were made using Tukey's test with *P<0.05, **P<0.01, and ***P<0.001 being considered significantly different. The number of animals used is stated in the figure legends. A minimum of 3 rats and 3 technical replicates were used.
Results
CFU-f derived from pour-off cultures
CFU-f numbers were analyzed in pour-off cultures from bone marrow over a time period of 14 days. Initially, the cells were cultured in high-glucose medium. The assay shows the number of CFU-f derived every day from 1 initial bone marrow culture (see Fig. 1). In addition we stained for the levels of spontaneous osteogenic differentiation in the colonies (Fig. 1A). Around 50% of the cells showed ALP staining (Fig. 1A, B) and the CFU-f frequency was not different between male or female rat-derived bone marrow. The number of colonies derived from the initial bone marrow mixtures was stable for 14 days at around 200 colonies (Fig. 1A, B).
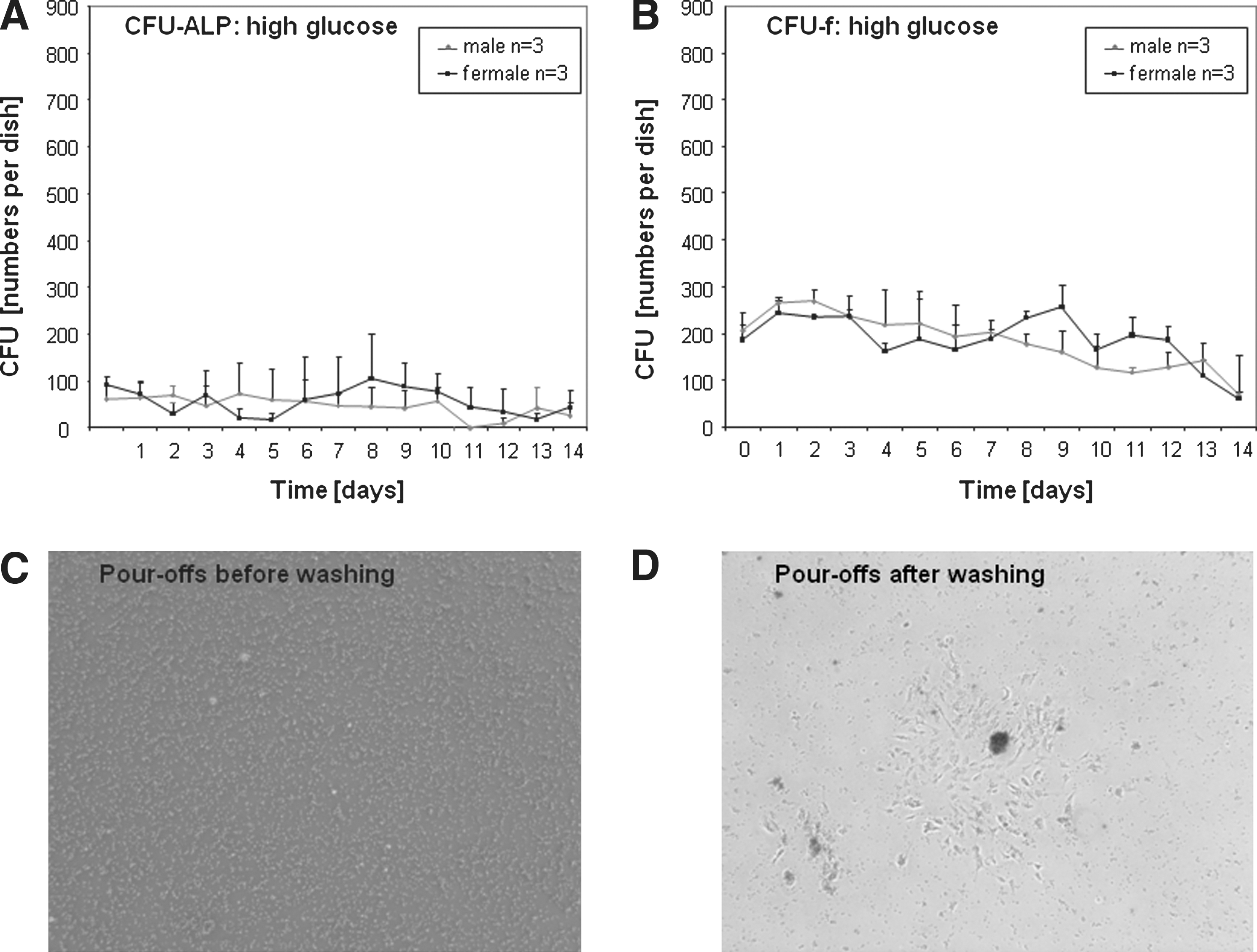
CFU numbers in high-glucose media. Fresh rat bone marrow was cultured in a CFU assay and the supernatant was transferred every day into a new dish. This was done for 14 days and then stopped. All cells were cultured for 14 days and then stained for alkaline phosphatase
After washing off the nonadherent cells, colonies of adherent fibroblastic cells were found with dense clusters of nonadherent cells loosely associated with the surface of the colonies. Observations suggest that these clusters are important for the sustainability of the cultures and that if the loosely attached clusters are not transferred to the next dish, for example, because of insufficient washing, the cultures lose their ability to generate new CFU-f.
As glucose levels are known to affect proliferation of classical adherent MSC cultures, we investigated the effects of low glucose DMEM (1 g/L) on the expansion of pour-off cells. In cultures containing low-glucose media we found that 2–3 times more colonies were generated throughout the entire culture period (Fig. 2A, B). Again no differences were found between male and female rat-derived cells. A representative series of pour-off culture-derived CFU-fs derived from young bone marrow cells cultured in low glucose is shown in Fig. 2C.
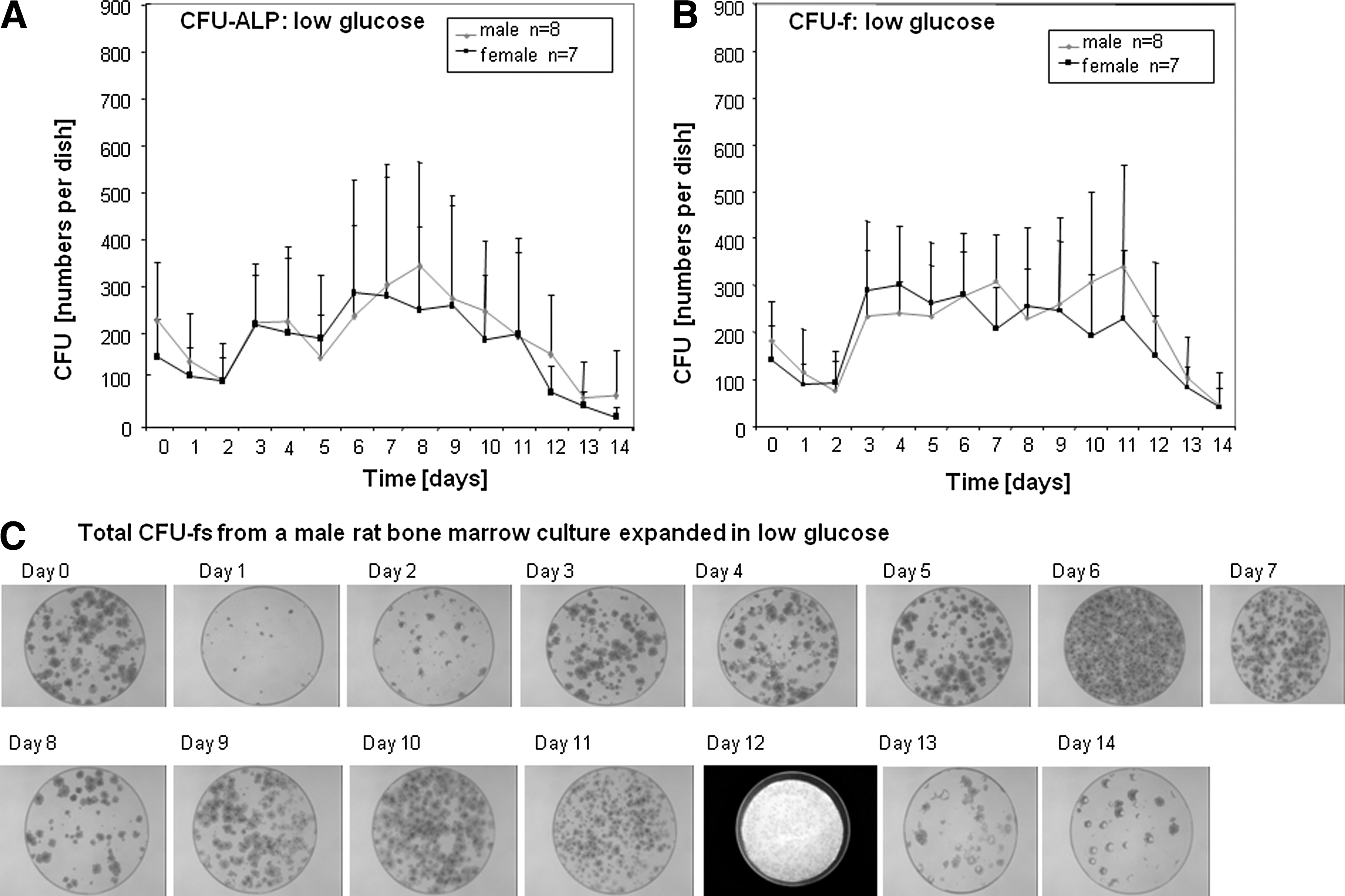
CFU numbers in low-glucose media. Fresh rat bone marrow was cultured in a CFU assay and the supernatant was transferred every day into a new dish. This was done for 14 days and then stopped. All plates were cultured for 14 days and then stained for alkaline phosphatase
CFU-f numbers decline with age of rats [20]. We therefore repeated the experiments with bone marrow from adult rats (12 months). Surprisingly there were no significant differences between young and adult bone marrow pour-off cultures (Fig. 3A, B).

CFU numbers derived from bone marrow of aged rats. Fresh rat bone marrow from 12-month-old rats was cultured in a CFU assay and the supernatant was transferred every day into a new dish. This was done for 14 days and then stopped. All plates were cultured for 14 days and then stained for alkaline phosphatase
We followed the cellularity and the frequency of CFU-f in the suspension cultures. Over a period of 4 days, total cell numbers decreased from 4×106 to 0.2×106 cells/mL (Fig. 4A). Simultaneously, the numbers of CFU-f in the nonadherent population of these cultures increased by almost 2 orders of magnitude (Fig. 4B). We tried to further characterize the cultures so we stained the cell clusters that are being transferred during the pour-offs and that give rise to the new MSCs (Fig. 4C–F). These clusters were stained for pluripotency markers and we found them to be positive for Nanog, Oct4, and Sox2 (Fig. 4G–I). Analysis of the proliferation of classical adherent MSCs compared with pour-off cultures showed that they grew with the same speed (Fig. 5).

Cell characterization. Cells were counted in the cultures of the first 4 days to follow the changes in cellularity

Proliferation times. MSCs and pour-off cells were seeded at different densities between 2,500 and 80,000 cells per well. The cells were cultured for up to 144 h (between 24 and 144 h) and the proliferation was measured using alamar blue.
Discussion
NA-BM-MSCs are a mixture of cells derived from the bone marrow. We have shown in mice that these cells can repopulate the hematopoietic system as well as other organs like the liver [14]. Allogeneic Na-BM-MSCs were transplanted and no signs of rejection were observed [14,15].
Glucose
As we found that CFU-f numbers decreased in type I diabetic rats [13], we wanted to test the effect of glucose levels on the expansion of Na-BM-MSCs in culture. Hyperglycemia is frequently encountered as part of many pathologic conditions, including diabetes mellitus; as a postinjury stress response, during sepsis; and as part of an inflammatory response. Our former studies confirmed that high-glucose media [16] or a high-plasma glucose level [13] induces senescence, decreases proliferation rates, and increases apoptosis in rat MSCs. Surprisingly media containing 5.5 mM glucose are currently most often used and recommended for maintaining human MSCs in culture [21]. Glucose had a profound effect on CFU-f numbers obtained in the pour-off cultures. High-glucose level led to a reduction in the CFU-f frequency over the whole culturing time. The beneficial effect of low glucose, however, was gradually declining to the level observed in high-glucose media after around 14 days. We did not observe any differences due to the gender of the Na-BM-MSC donors.
A study on human umbilical cord blood–derived passaged [5 –8] MSCs showed an increased proliferation at 25 mM glucose compared with 5 mM glucose (n.b. serum glucose concentration in normal individuals is 5.55 mM) [22]. Low-glucose compared with high-glucose concentration did not affect osteogenic or adipogenic differentiation; however, no quantification was used and the MSCs were kept in serum-free medium prior to the experiment. Oct4 or Foxo3a expression levels were not different, only Sox2 showed increased expression under 25 mM glucose. Glucose induces TGF-β expression via the PKC phosphorylation, which leads to increased proliferation of MSCs [23].
Colony forming units of MSCs were increased according to addition of glucose and the maximum increase was observed at a concentration of 25 mM glucose (5 mM glucose, 53±2.5 CFU/wells; 25 mM glucose, 69±4.3 CFU/wells).
Short-term exposure (24–48 h) to glucose did not affect MSC proliferation or growth factor secretion [24]. Recent evidence suggests that high glucose may suppress growth factor production in rat multipotent adult progenitor cells, which are closely related to MSCs [22].
Ageing
As the Na-BM-MSCs have demonstrated the ability to form CFU-f and have been shown to increase in frequency over time in bone marrow cultures derived from young animals [15], we wanted to test whether this capacity was preserved for bone-marrow-derived Na-BM-MSCs from aged animals. The Na-BM-MSC cultures produce an astonishing number of CFU-f over a period of time when transferred on a daily basis [15]. Interestingly we found the same level of CFU-f in Na-BM-MSCs derived from young or aged rats. Studies on MSC numbers of young compared with aged humans, rats, or mice frequently show a reduction in colony numbers using the CFU-f assay [20,25,26]. This might suggest that aging is affecting the frequency of more mature, more differentiated stem cells, the classical plastic adherent MSCs, but not the potential of more unmature cells, the nonadherent MSCs.
Our findings of extensive MSC expansion using the nonadherent MSC fraction are in agreement with the results by Zhang et al. on rat Na-BM-MSCs [15]. Fricke however found a fast decline after 4 days in Na-BM-MSCs derived from mice [27]. This might be either due to species differences or different cultivation properties. Na-BM-MSCs have some features similar to the lately described very small embryonic-like stem cells found in the bone marrow and cord blood [28].
To analyze the cultures and cells initiating the new poured-off colonies, we stained the MSCs in the dishes. We found larger clusters of cells that were positive for pluripotency markers. Such larger colonies are not uncommon in unpassaged MSC cultures but do get lost during culturing. We could observe such clusters in most of the NA-BM-MSC cultures. It might be that these clusters contain cells that are the CFU-initiating cells. No tumor or teratoma formation was observed in mice after transplantation using mice, rat, or human cells [14,29]. These cells are therefore an interesting alternative source of stem cells for regenerative therapies.
Footnotes
Author Disclosure Statement
No competing financial interests exist.