
Abstract
The reduction of adult stem cell self-renewal can be an important mechanism of aging. MicroRNAs have been reported to be involved in aging processes. Through a microarray approach, we have identified miR-486-5p, the expression of which is progressively expressed in human adipose tissue-derived mesenchymal stem cells (hAT-MSCs) with aging. Overexpression of miR-486-5p induces a premature senescence-like phenotype and inhibits proliferation of hAT-MSCs and inhibits adipogenic and osteogenic differentiation, whereas inhibition of miR-486-5p has the opposite effects. miR-486-5p regulates the expression of silent information regulator 1 (SIRT1), a major regulator of longevity and metabolic disorders. Decrease of SIRT1 deacetylase activity in hAT-MSCs is correlated with their passage number. miR-486-5p inhibits SIRT1 expression through a miR-486-5p binding site within the 3′-untranslated region of SIRT1. Overexpression of miR-486-5p inhibits SIRT1 deacetylase activity in hAT-MSCs, and transfection of miR-486-5p inhibitor shows the opposite effect. Downregulation of SIRT1 in hAT-MSCs induces senescence and inhibits cell proliferation. Exposure to high glucose increases miR-486-5p expression and inhibits SIRT1 expression in hAT-MSCs. Our data pinpoint miR-486-5p as an endogenous inhibitor of SIRT1 that promotes hAT-MSCs senescence and is potentially applicable to therapeutic manipulation of hAT-MSCs dysfunction in metabolic disorders.
Introduction
R
Mesenchymal stem cells (MSCs) represent a multipotent adult stem cell population that, given the appropriate culture conditions, is able to differentiate into different mesodermal cell lineages including osteocytes, chondrocytes, and adipocytes (MSCs: building blocks for molecular medicine in the 21st century [5]. MSCs can also differentiate into myocytes and cardiomyocytes and even into cells of non-mesodermal origin, including hepatocytes [6 –8]. Therefore, human MSCs are promising for various therapeutic applications, and their use is concurrently being tested in various clinical settings [9]. The tissues of any organism are continuously renewed by adult stem cells, and impairment of their function would result in aging [10]. Because murine and human MSCs exhibit reduced differentiation potential upon long-term in vitro culture [11,12], senescence of MSCs might limit their therapeutic applications. Thus, studies of in vitro senescence of MSCs are important for basic research and for quality control in cellular therapy.
Molecular mechanisms of cellular senescence are still largely unknown. The global analysis of mRNA and protein abundance during replicative senescence provides useful clues about molecular mechanisms of senescence. Although changes in several transcription factors during replicative senescence modulate protein abundance, the importance of posttranscriptional processes, including altered mRNA stability and translation, is becoming apparent [13].
microRNAs (miRNAs) are approximately 22-nucleotide (nt) long noncoding RNAs that associate with a target mRNA in a sequence-specific manner and typically repress gene expression by lowering the stability of the mRNA, decreasing its translation rate, or performing both functions [14]. miRNAs are involved in the posttranscriptional regulation of genes implicated in complex cellular processes such as aging, cancer, proliferation, differentiation, and apoptosis [15]. Changes in miRNAs expression during replicative senescence and their roles have been reported. Increased p16 expression during replicative senescence in WI-38 human diploid was associated with decreased levels of miR-24, a miRNA that was predicted to associate with the p16 mRNA coding and 3′-untranslated regions (UTRs) [16]. miR-15b, miR-24, miR-25, and miR-141 overexpression decreases replicative senescence of WI-38 cells through the inhibition of MKK4 protein expression, and inhibitors of these miRNAs increases replicative senescence [17]. miR-217 [18] or miR-34 [19] overexpression induces senescence of endothelial cells through inhibition of silent information regulator 1 (SIRT1). Wagner et al. reported changes in the miRNA expression profile of replicatively senescent human MSCs [20], and Bork et al. showed that overexpression of miR-369-5p, miR-371, and miR-499 reduced proliferation of hMSCs [21]. The impairment of the functions of endothelial progenitor cells (EPCs) in diabetic patients [22] and high glucose-induced inhibition of proliferation of EPCs [22,23] have been reported. We previously showed that exposure of high glucose inhibits the proliferation of adipose tissue-derived MSCs (AT-MSCs) and impairs proangiogenic action of these cells [24]. However, the role of miRNA in these changes is still not clear.
In this study, we determined the role of miR-486-5p in replicative senescence of human AT-MSCs (hAT-MSCs) and its change in high glucose condition.
Materials and Methods
Materials and reagents
BLOCK-iT Pol II miR RNAi Expression vector, Ncode miRNA first-strand cDNA synthesis kits and Lipofectamine Plus were from Invitrogen. mirVana miRNA isolation kit, miRVana miRNA probe construction kit, and pMIR-Report vector were from Ambion. Polyclonal antibodies (anti-SIRT1 and anti-β-actin) were from Cell Signaling Technology. Protein assay kit was from Bio-Rad Laboratories. Peroxidase-conjugated secondary antibody (anti-rabbit or anti-mouse) were from Amersham Biosciences. ECL detection kit was from Amersham Biosciences. DharmaFECT Transfection Reagent and Small interfering RNA (siRNA) duplex oligos (on-TARGET plus SMART pool) were from Dharmacon. Luciferase Assay System was from Promega.
Cell culture
All protocols involving human subjects were approved by the Institutional Review Board of Pusan National University. Superfluous materials were collected from 4 individuals undergoing elective abdominoplasty after informed consent was given by each individual. The hAT-MSCs were isolated according to the methods described in previous studies [25,26]. Isolated cells were cultured in with α-modified Eagle's medium (α-MEM), containing 10% fetal bovine serum (FBS). Every batchs of culture expanded cells (P2-P3) were characterized by the determination of their surface markers expression and differentiation potential before using experiments. The batches that more than 95% of cultured cells express CD105, CD73, and CD90 and less than 2% express CD45 and CD14, and show high differentiation potential to osteogenic and adipogenic lineages were used for further experiments.
miRNA microarray
Total RNA (200–1,000 ng) isolated from P2 or P15 cells was polyadenylated, reverse-transcribed to cDNA using an oligo(dT) primer. After cDNA synthesis, miRNAs were individually immobilized on specific oligonucleotides (miRNA-specific Oligo_MSO). Hybridized oligonucleotides were then extended and ligated to generate amplifiable templates. A PCR reaction was performed with fluorescently labeled universal PCR primers. The PCR products were hybridized to each miRNA bead array (human-miRNA-V1 bead chip containing 735 human miRNA sequences that were based on miR Database version 9.1) for 30 min at 60°C, 16–18 h at 45°C according to the manufacturer's instructions (Illumina, Inc.). Arrays were scanned with an Illumina bead array Reader confocal scanner according to the manufacturer's instructions. Array data export processing and analysis was performed using Illumina BeadStudio.
Viral vector construction and transduction
The engineered pre-miRNA sequence was cloned into the cloning site of a BLOCK-iT Pol II miR RNAi Expression vector that is flanked on either side with sequences from hsa-miR-486-5p (mature sequence: 5′-UCCUGUACUGAGCUGCCCCGAG) to allow proper processing of the engineered pre-miRNA sequence. Other sub-cloning and viral transduction procedures were performed as previously described [27]. We used virus titers ranging from 5×105 to 1×107 transducing units/mL.
Senescence-associated β-galactosidase
Cell samples on a 6-well plate were fixed with 2% formaldehyde and 0.2% glutaraldehyde in phosphate-buffered saline (PBS) for 10 min at 4°C. The plates were rinsed with PBS and incubated with a senescence-associated beta-galactosidase (SA β-gal)staining solution containing 40 mM sodium acetate (pH 6.0), 5 mM K3Fe(CN)6, 2 mM MgCl2, 150 mM NaCl, and 1 mg/mL 5-bromo-4-chloro-3-indoyl β-D galactoside (X-Gal) solution for 16 h. The number of blue cells in 20 fields was counted at 200× magnification.
Evaluation of cell proliferation
The cells were detached using Hank's balanced salt solution containing 0.05% trypsin and 0.02% ethylenediaminetetraacetic acid (EDTA) to determine the rate of proliferation. The cells were plated on a 6-well plate at a density of 5×103 cells/well. After 3 days, the cells were trypsinized and stained with 0.4% trypan blue (Sigma). Sixty cells were transferred to a 100 mm plate to perform the colony forming unit (CFU) assay. The number of cells in each colony was then counted.
Reverse transcription–PCR analysis
Total cellular RNA was isolated from hAT-MSCs and reverse transcribed using conventional protocols. The primer sequences used in the experiment were as follows: GAPDH: 5′-TCC ATG ACA ACT TTG GTA TCG-3′, 5′-TGT AGC CAA ATT CGT TGT CA-3′; SIRT1: 5′-GCA GAT TAG TAG GCG GCT TG-3′, 5′-TCT GGC ATG TCC CAC TAT CA-3′. All primer sequences were generated from established GenBank sequences.
Determination of osteogenic and adipogenic differentiation
Adipogenic differentiation was induced by culturing MSCs for 7 days in an adipogenic medium (10% FBS, 1 μM dexamethasone, 0.5 mM/mL 3-isobutyl-1-methylxanthine, and 200 μM indomethacin in α-MEM) and assessed by the use of an Oil red O stain as an indicator of intracellular lipid accumulation. To obtain quantitative data, 1 mL isopropyl alcohol was added to the stained culture dish. Osteogenic differentiation was induced through the culturing of the cells for 10 days in osteogenic medium (10% FBS, 0.1 μM dexamethasone, 10 mM β-glycerophosphate, and 50 μM ascorbic acid in α-MEM), and extracellular matrix calcification was estimated using Alizarin red S stain. Osteogenic differentiation was quantified through the measurement of the Alizarin red-stained area and density in 12-well dishes, using an image analysis program (Image Gauge ver 3.1; Fuji).
Flow cytometric analysis
hAT-MSCs were cultured in control medium for 72 h before analysis. Flow cytometry was performed on a FACScan argon laser cytometer (Becton, Dickinson and Company, Franklin Lakes, NJ,
Real-time PCR
Small RNA species-enriched RNA isolation was performed per the manufacturer's instructions (mirVana miRNA isolation kit; Ambion). miRNA was reverse-transcribed using the Ncode miRNA first-strand cDNA synthesis kit (Invitrogen), according to the manufacturer's specified guidelines. The forward primer sequences were designed to correspond with the mature miRNA sequences. The forward primer sequences used in the experiment were as follows: miR-486-5p: 5′-TCC TGR ACT GAG CTG CCC CGA G-3′; miR-34a: TGG CAG TGT CTT AGC TGG TTG TT-3′; miR-217-5S primer: 5′-GAT CGG GCG CGT TCA GGG TGG TAT-3′. Real-time quantitation was performed with the LightCycler assay, using a fluorogenic SYBR Green I reaction mixture for PCR with the LightCycler Instrument. Threshold cycle (Ct) for each miRNA was extracted using Thermal Cycler Dice Real Time System ver2.10B. The raw Ct was normalized using the values of 5S rRNA [27].
Northern blot
Oligonucleotides complementary to the mature miRNAs were end-labeled with a T7 promoter sequence (miRVana miRNA probe construction kit; Ambion), and were used as probes. The probe sequences were as follows: miR-486-5p: 5′-TCC TGT ACT GAG CTG CCC CGA G cctgtctc-3′; miR-34a: 5′-TGG CAG TGT CTT AGC TGG TTG TT cctgtctc-3′; and U6: 5′-GCA GGG GCC ATG CTA ATC TTC TCT GTA TCG cctgtctc-3′. Other northern blot procedures were performed as described [27].
Western blot analysis
Confluent hAT-MSCs and adipose tissue were treated under the appropriate conditions and lysed, after which the protein content of the lysate was determined using a protein assay kit. The proteins were loaded on 10% sodium dodecyl sulfate polyacrylamide gels, transferred to nitrocellulose membranes (Hybond-ECL; Amersham Biosciences), and probed with polyclonal antibodies. Immunoreactive bands were detected with anti-rabbit or anti-mouse peroxidase-conjugated secondary antibodies and visualized via enhanced chemiluminescence.
RNA transfection
Anti- miRNA inhibitors (anti-miRs) and a scrambled RNA oligomer were purchased from Ambion. These reagents were transfected into hAT-MSCs at a final concentration of 50 nM using DharmaFECT Transfection Reagent, per the manufacturer's instructions. siRNA duplex oligos (on-TARGET plus SMART pool) targeting SIRT1 mRNA or a non-targeting duplex oligo (negative control) were transfected into cells using the DharmaFECT Transfection Reagent.
Reporter vectors and DNA constructs
A putative miRNA 486-5p-recognition element (single copy) from the SIRT1 gene was cloned into the 3′-UTR of a firefly luciferase reporter vector according to the manufacturer-specified guidelines. The oligonucleotide sequences were designed to carry the HindIII and SpeI sites at their extremities to allow ligation into the HindIII and SpeI sites of pMIR-Report. The oligonucleotides for pMIR-SIRT1 used in these studies were 5′-CTA GTG TAA TAA TTG TGC AGG TAC AGG AAT TGA-3′ and 5′-AGC TTC AAT TCC TGT ACC TGC ACA ATT ATT ACA-3′.
Reporter gene assay
All transient transfections were conducted using Lipofectamine Plus. The pMIR-Report, pMIR-SIRT1, and pMIR-β-gal plasmids were used as reporter constructs. Cells were harvested 48 h after transfection in cell culture lysis (CCLR) buffer and were subsequently assayed for luciferase activity. Transfections were performed in duplicate, and all experiments were repeated several times. Luciferase expression was normalized to β-galactosidase activity in all cases.
Measurement of SIRT 1 deacetylase activity in hAT-MSCs
NAD+-dependent SIRT1 activity was determined with a SIRT1 Fluorometric Kit (Biomol International) according to the manufacturer's instructions and the method of Escande et al. [28]. Values were determined by reading fluorescence on a fluorometric plate reader (VICTOR™ ×3 Multilabel Plate Reader) with an excitation wavelength of 360 nm and an emission wavelength of 460 nm.
Statistical analysis
All results are presented as the mean±standard error of the mean. Comparisons between 2 groups were analyzed via Student's- tests. Multiple group comparisons were conducted by 1-way analysis of variance with Tukey adjustments. P values <0.05 were considered statistically significant.
Results
Changes in miR-486-5p levels during repeated rounds of hAT-MSCs replication
To induce replicative senescence, we repeatedly subcultured hAT-MSCs. Based on the cumulative population doubling level, it was assumed that hAT-MSCs began to lose their proliferation potential at around the 10th passage. To confirm the senescence of late stage hAT-MSCs, SA β-gal staining was performed. At the 15th passage, we confirmed that hAT-MSCs completely lost their self-renewal capability, as evidenced by a substantial decrease of proliferation potential and the development of senescent phenotypes, which were revealed using SA β-gal staining (Fig. 1A).

Replicative senescence of human adipose tissue-derived mesenchymal stem cells (hAT-MSCs).
To test how the pattern of expressed miRNAs is affected by replicative senescence, we performed miRNA microarray analysis of total RNAs isolated from P2 and P15 AT-MSCs and identified 20 miRNAs of which expression was changed more than 2-fold among 735 miRNAs (Table 1). From the list we selected 5 miRNAs that may be related to control of cell proliferation from Pubmed database. In this experiment we focused on the function of miR-486-5p because real-time PCR analysis showed the highest increase of its expression in P15 cells among indentified miRNAs. To examine the role of miR-486-5p on the replicative senescence of hAT-MSCs, we measured the level of miR-486-5p expression during replicative senescence by northern blot and real-time PCR analysis. The results showed that miR-486-5p expression increased according to the passage numbers (Fig. 1B, C), like the expression of miR-34a which induces EPC senescence and impedes angiogenesis via suppressing SIRT1 [19]. We also measured the changes in the expression of miR-217 during the passage of hAT-MSCs. Real-time PCR analysis showed that expression level of miR-217 in hAT-MSCs is undetectable, although it is expressed in human umbilical vein endothelial cells (data not shown).
n.d., not determined; PCR, polymerase chain reaction.
Effect of miR-486-5p lentivirus transduction on cellular senescence of hAT-MSCs
To determine the role of miR-486-5p in hAT-MSCs function, we used a lentivirus system to overexpress miR-486-5p in hAT-MSCs. Because the lentivirus vector contains the green fluorescent protein (GFP) coding sequence driven by a cytomegalovirus (CMV) promoter, the efficiency of lentivirus transduction can be determined with a fluorescent microscope. Single transductions with the lentivirus showed that most hAT-MSCs (>90%) expressed GFP and that miR486-5p-overexpressing cells were morphologically similar to naive hAT-MSCs or LV-miLacZ-infected hAT-MSCs (Fig. 2A). Real-time PCR analysis and northern blot analysis with a miR-486-5p probe confirmed that hAT-MSCs transduced with the miR-486-5p lentivirus exhibited increased miR-486-5p expression (Fig. 2B, C).
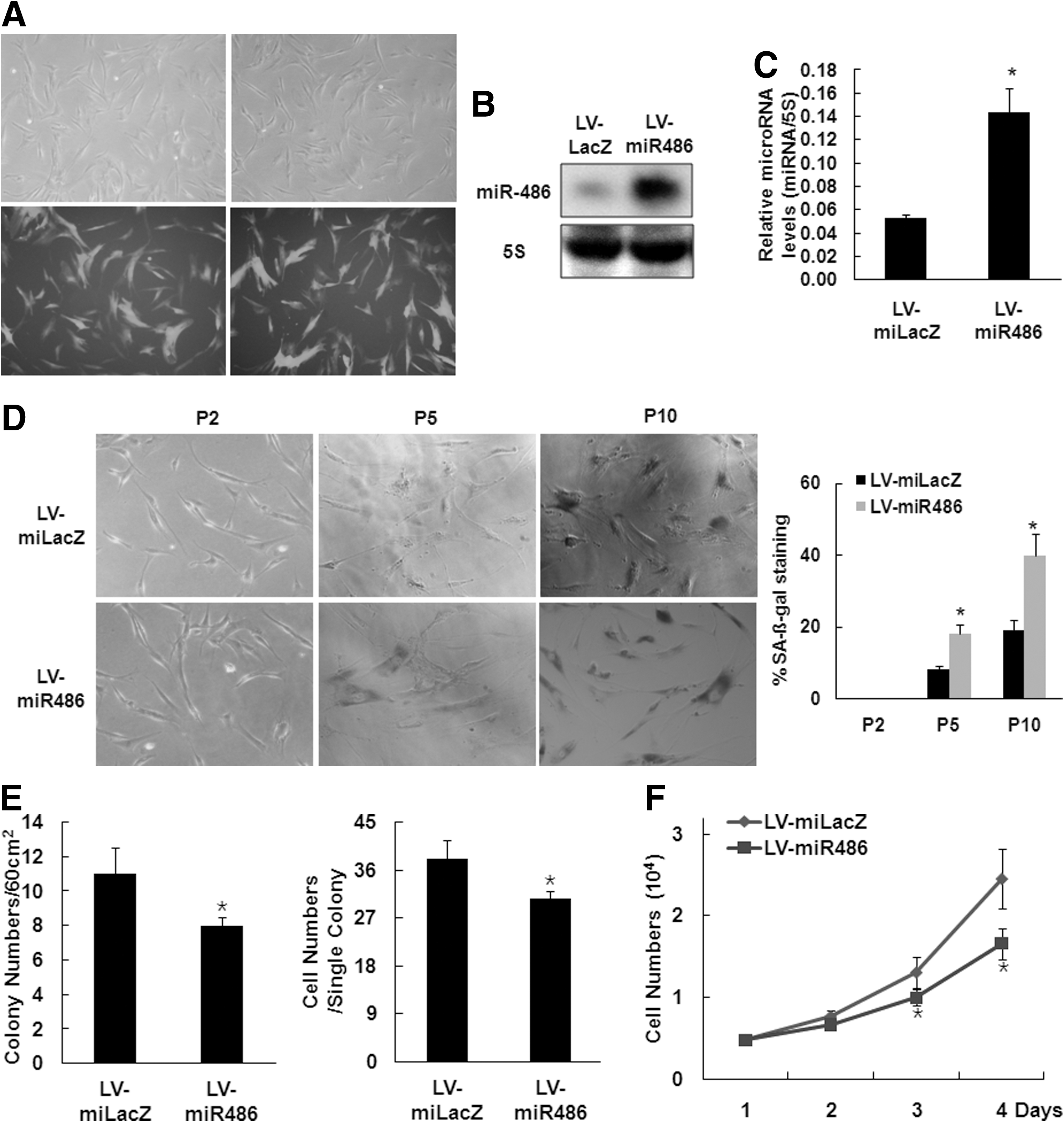
Overexpression of miR-486-5p induces cellular senescence of hAT-MSCs.
To confirm the role of miR-486-5p in cellular senescence of hAT-MSCs, SA β-gal staining was performed. According to the cumulative population doubling level, hAT-MSCs overexpressing miR-486-5p showed a strong increase in SA β-gal activity compared with the control infection group (Fig. 2D).
We also determined the effect of miR-486-5p on hAT-MSCs proliferation. After infecting the hAT-MSCs with LV-miR486-5p, we plated 60 infected cells on 100-mm culture plates and counted the number of colonies, and the number of cells in each colony, after 1 week. The numbers of CFU and numbers of cells per colony in LV-miR486-5p-infected hAT-MSCs were significantly lower than those in LV-miLacZ-infected hAT-MSCs (Fig. 2E). Cell counting on the indicated days after plating 5×103 cells/well showed that LV-miR486-5p-infected cells grew slower than LV-miLacZ-infected hAT-MSCs (Fig. 2F).
Effect of miR-486-5p inhibitor on cellular senescence of hAT-MSCs
Recent studies have used reverse complement 2′-O-methyl sugar-modified RNA to block miRNA function in cell-based systems [29]. Thus, we determined the role of miR-486-5p by investigating the effect of a specific miRNA inhibitor using oligo transfection. According to the cumulative population doubling level, anti-miR-486-5p-transfected hAT-MSCs showed a significant delay in SA β-gal activity compared with the control oligonucleotide-transfected hAT-MSCs (Fig. 3A).
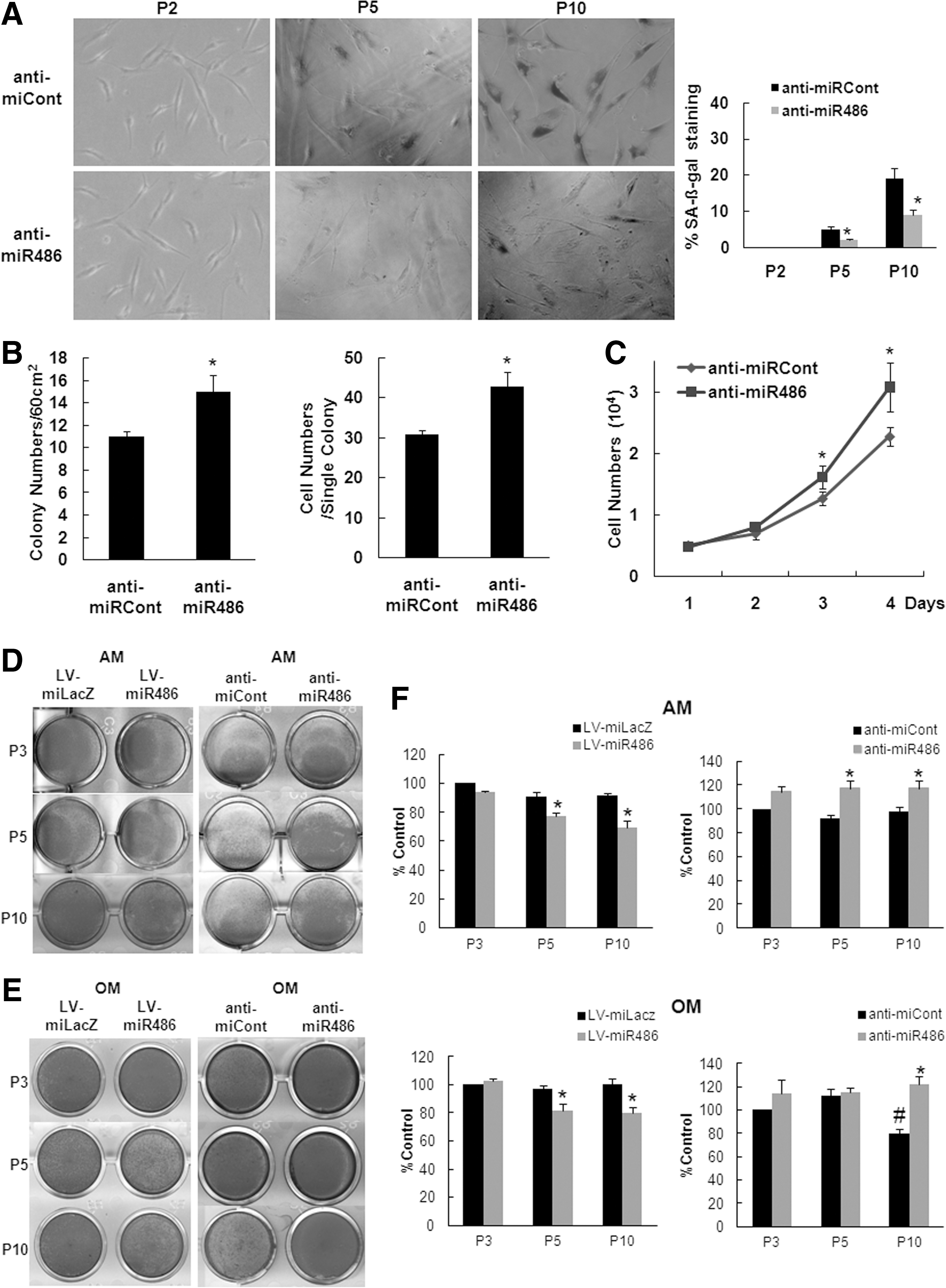
Inhibition of miR-486-5p delays cellular senescence of hAT-MSCs. hAT-MSCs were transfected with anti-miR Cont or anti-miR 486-5p to assess senescence.
In a CFU assay, anti-miR-486-5p-transfected hAT-MSCs displayed higher colony numbers and cell numbers per colony than control oligonucleotide-transfected hAT-MSCs (Fig. 3B). Cell counting on the indicated days after plating 5×103 cells/well showed that the anti-miR-486-5p-transfected hAT-MSCs proliferated more than the control-transfected cells (Fig. 3C).
To determine whether miR-486-5p modulation affects the properties of AT-MSCs, we examined the percentage of surface markers expression and differentiation potentials. Inhibition of miR-486-5p did not affect the pattern of surface marker expressions (Inhibitor transfected cells vs. controls, CD90=98.6 vs. 97.2, CD73=96.4 vs. 97.5, CD105=95.7 vs. 95.3, CD45=1.2 vs. 0.9, CD14=1.4 vs. 1.1 and HLA-DR=0.9 vs. 0.7). Overexpression of miR-486 inhibited adipogenic and osteogenic differentiation and transfection of miR-486-5p inhibitor showed the opposite effect on them (Fig. 3D–F).
miR-486-5p targets the 3′UTR of SIRT1 mRNA
Among the candidate miR-486-5p targets in the Targetscan 5.1 database, SIRT1 was the best candidate for explaining the miR-486-5p-induced replicative senescence of hAT-MSCs because SIRT1 is important in the aging of mammalian cells [30]. Therefore, we investigated SIRT1 expression levels during senescence. The mRNA and protein levels of SIRT1 gradually decreased according to cellular senescence (Fig. 4A, B). We also determined whether replicative senescence is accompanied by the decrease of the endogenous SIRT1 activity of hAT-MSCs. SIRT1 deacetylase activity decreased according to the passage number of hAT-MSCs (Fig. 4C).
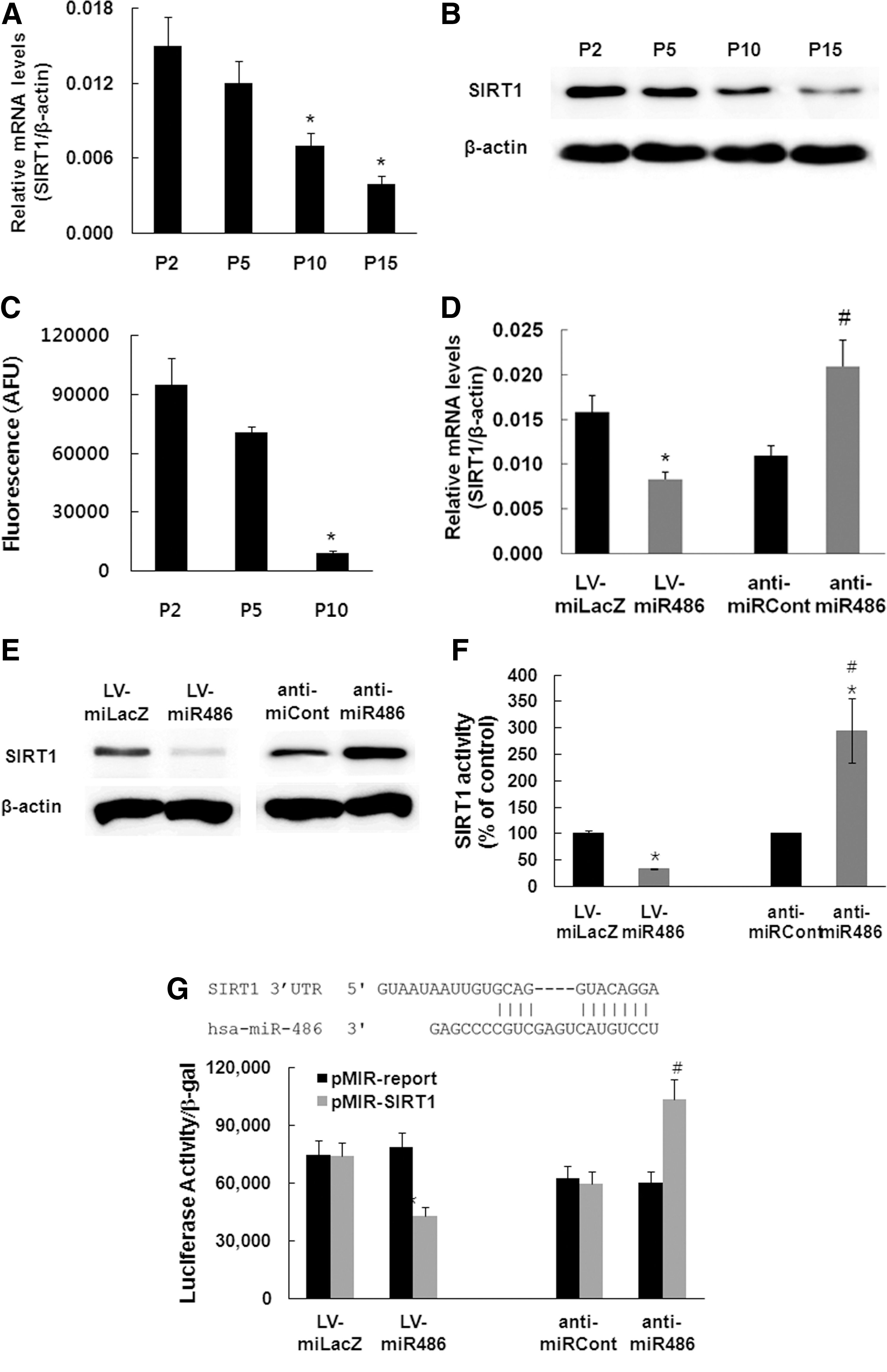
miR-486-5p targets the silent information regulator 1 (SIRT1) mRNA 3′untranslated region (UTR).
To further confirm the relationship between miR-486-5p and SIRT1, we determined the mRNA and protein levels of SIRT1 in hAT-MSCs overexpressing miR-486-5p or cells transfected with a miR-486-5p-specific inhibitor. Real-time PCR and western blot analyses showed that miR-486-5p-overexpressing hAT-MSCs exhibit less SIRT1 expression and that anti-miR-486-5p-transfected hAT-MSCs exhibit increased SIRT1 expression (Fig. 4D, E). We examined the effect of miR-486-5p modulation on SIRT1 deacetylase activity. Overexpression of miR-486-5p inhibited SIRT1 activity, whereas the transfection of the SIRT1 inhibitor increased it compared with control oligonucleotide-transfected hAT-MSCs (Fig. 4F). A luciferase reporter assay was used to demonstrate that miR-486-5p directly decreased SIRT1 expression. We inserted the portion of the SIRT1 3′UTR that is complementary to miR-486-5p into a luciferase expression vector and used the resulting construct to transfect hAT-MSCs. Transfection of LV-miR486-5p-infected hAT-MSCs with the parental luciferase construct (without the SIRT1 3′UTR) or a scrambled construct did not significantly change the expression of the reporter. However, cells transfected with a luciferase construct in which the miR-486-5p target site from the SIRT1 3′UTR (pMIR-SIRT1) was inserted exhibited significantly lowered luciferase activity in LV-miR486-5p-infected hAT-MSCs compared with control lentivirus-infected cells. Cotransfection with an anti-miR486-5p oligo increased the luciferase activity of pMIR- SIRT1 in hAT-MSCs relative to cotransfection with a control oligo (Fig. 4G).
Effect of SIRT1 oligo transfection on cellular senescence of hAT-MSCs
To determine the role of SIRT1 in cellular senescence of hAT-MSCs, we suppressed SIRT1 expression in hAT-MSCs with an RNA interference technique using oligonucleotide transfection. Reverse transcription–PCR analysis confirmed that RNAi effectively inhibited SIRT1 expression in hAT-MSCs (Fig. 5A). According to the cumulative population doubling level, SIRT1 oligo transfection in hAT-MSCs showed a strong increase in SA β-gal activity compared with the control infection group (Fig. 5B). To determine the effect of SIRT1 oligo transfection on hAT-MSCs proliferation, a CFU assay and total cell counting after cell plating were carried out. In the CFU assay, SIRT1 oligonucleotide-transfected hAT-MSCs displayed fewer colonies and fewer cells per colony than control oligonucleotide-transfected hAT-MSCs (Fig. 5C). Cell counting on the indicated days after plating 5×103 cells/well also showed that the SIRT1 oligonucleotide-transfected hAT-MSCs proliferated less than the control-transfected cells (Fig. 5D).
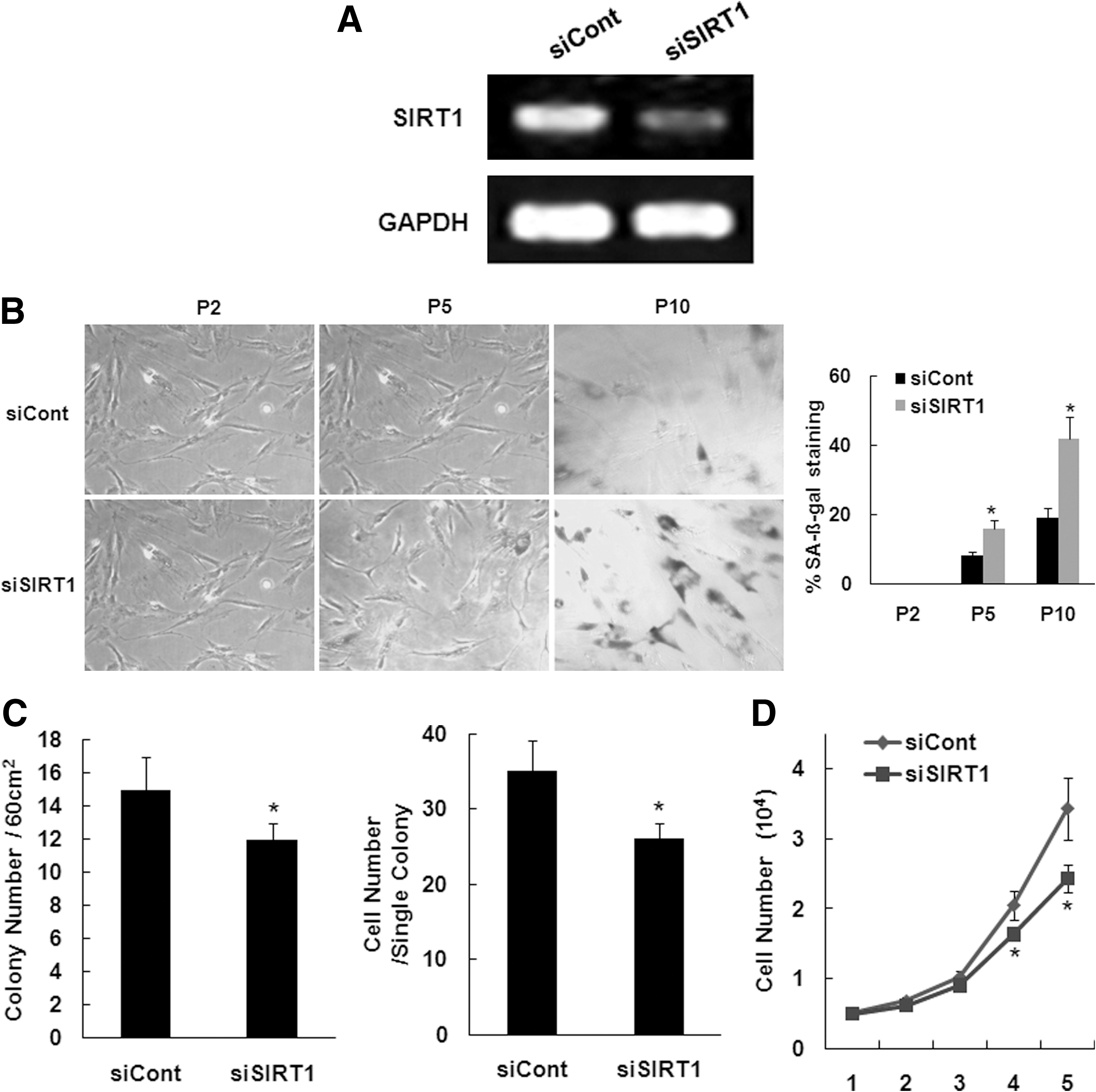
Effect of SIRT1 RNAi on hAT-MSC senescence.
Effect of high glucose on miR-486-5p and SIRT1 expression of hAT-MSCs
SIRT1 plays an important role in regulating pathogenesis of chronic conditions, including diabetes and aging-related disease [31]. We analyzed the effect of high glucose media on miR-486-5p and SIRT1 expression. Real-time PCR analysis showed that high glucose treatment (25 and 50 mM) induced an increase in miR-486-5p (Fig. 6A) but did not affect miR-34 levels. To determine whether the high glucose-induced decrease of SIRT1 expression resulted from the alteration of miR-486-5p expression, we examined the effect of the miR-486-5p inhibitor on high glucose-induced decrease of SIRT1 expression. Transfection of the miR-486-5p inhibitor blocked the decrease of SIRT1 expression induced by the incubation of hAT-MSCs in high glucose media (Fig. 6B). To determine the effect of high glucose on hAT-MSCs proliferation, hAT-MSCs cultured in control or high glucose media were counted on the indicated days. The treatment of high glucose inhibited proliferation of hAT-MSCs in a dose-dependent manner (Fig. 6C).

Effect of high glucose on miR-486-5p and SIRT1 expression of hAT-MSCs. hAT-MSCs proliferation was treated with control (5.5 mM glucose) or high glucose (25 mM or 50 mM glucose) media for 2 weeks. miRNAs were isolated from each hAT-MSCs population.
Discussion
Senescence is the state or process of aging at the cellular level, and this process is thought to be involved in both age-related diseases and tumorigenesis [32]. In this study, we explored the role of miRNAs in senescence of hAT-MSCs. The data presented are consistent with a role for miR-486-5p as a natural inhibitor of SIRT1 during hAT-MSCs senescence. First, miR-486-5p expression progressively increases during repeated rounds of cellular replication, which is paralleled by a decrease in SIRT1 expression. Second, modulation of miR-486-5p is able to induce cellular senescence in young cells, whereas inhibition of miR-486-5p partially rescues aging phenotypes in old cells and increases proliferation. Third, luciferase assays demonstrated that miR-486-5p directly binds to the 3′UTR of SIRT1 in hAT-MSCs. Fourth, the downregulation of SIRT1 increased the replicative senescence of hAT-MSCs. SIRT1, an NAD-dependent deacetylase that regulates gene expression by deacetylation of modified lysine residues on histones, transcription factors, and some transcription cofactors [33], and is an important determinant of longevity that plays a role in life-span regulation in diverse species [34,35]. The relationship between SIRT1 and cellular senenescence has been supported by the findings that increased SIRT1 activity rescues cellular senescence by inhibiting p53 acetylation in primary mouse embryo fibroblasts [36], endothelial cells [37], and vascular smooth muscle cells [38].
The role of SIRT1 in cell proliferation is variable according to cell types. SIRT1 activation may increase cancer risk, based on observations that SIRT1 deacetylates and inactivates p53 and other tumor suppressors [39,40]. On the contrary, recent studies suggest that SIRT1 can serve as a tumor suppressor. SIRT1 plays a key role in recruiting RAD51 and NBS1 to DNA double strand breaks to initiate the DNA damage repair process, and SIRT1 overexpression suppresses age-related transcriptional changes [41]. Moreover, SIRT1 overexpression reduces the incidences of colon cancer in APC+/− min mice [42] and spontaneous cancers in multiple tissues of SIRT1+/−; p53+/− mice [43]. SIRT1 is also active in primary cells, promoting proliferation of endothelial cells [44] and skeletal muscle precursor cells [45]. In this study, we demonstrated that miR-486-5p regulates proliferation of hAT-MSCs. Overexpression of miR-486-5p inhibited proliferation of hAT-MSCs, whereas downregulation of miR-486-5p increased proliferation. These results indicate that miR-486-5p negatively regulates the proliferation of hAT-MSCs, like other primary cells [44,45]. Although miR-486-5p inhibitor-transfection inhibited replicative senescence in AT-MSCs (Fig. 3), the cells still proceed to senescent state during long-term culture. This may be the result of incomplete inhibition of miR-486-5p or the involvement of other miRNAs besides of miR-486-5p, because changes in multiple miRNAs expression occurred during hAT-MSCs senescence as observed in our microarray data (Table 1).
To provide a human pathological perspective of these findings, we analyzed the effect of high glucose media on miR-486-5p and SIRT1 expression. The results demonstrated that exposure of hAT-MSCs to high glucose inhibits cell proliferation, as observed in the previous studies [24,46], and is accompanied by increased expression of miR-486-5p-and subsequent downregulation of SIRT1. Recent studies indicate that SIRT1 levels are downregulated in EPCs by high glucose exposure [47] and in adipose tissues of diabetic patients [48]. Sirt1 gain of function improves metabolic efficiency, promoting longevity and disease resistance [49,50] and SIRT1 expression is correlated with insulin sensitivity and energy expenditure in type 2 diabetic patients [48]. Further, resveratrol, an activator of SIRT1 [51], improves insulin sensitivity [52,53]. All of these data demonstrate that glucose homeostasis is under tight control of SIRT1 and that SIRT1 is an important therapeutic target of metabolic syndromes, including type 2 diabetes. Our findings that high glucose results in miR-486-5p-induced downregulation of SIRT1 can provide insight regarding functional impairment of adult stem cells in diabetic patients. Diabetic patients show impaired function of EPCs [22], and resveratrol improves the neovascularization of bone marrow mononuclear cells from diabetic mice [54], suggesting that an impairment of EPC function can be related to decreased SIRT1 function.
The well known miRNA that is related to senescence is miR-34 [19]. In fact, we also found that miR-34 expression increased in replicatively senescent hAT-MSCs. However, the expression of miR-34 was not altered by exposure to high glucose media (Fig. 6A). Therefore, miR-34 is not responsible for changes in SIRT1 expression following high glucose-induced inhibition of proliferation.
Our data showed that inhibition of miR-486-5p action increased differentiation potential in addition to cell proliferation and inhibition of senescence without affecting surface marker expression in AT-MSCs, indicating that modulation of miR-486-5p function can be used for enhancing therapeutic efficacy of AT-MSCs, if safety issues that may be accompanied by genetic modification of miRNA are properly handled.
In conclusion, we have identified an endogenous inhibitor of SIRT1, miR-486-5p. This discovery opens new avenues in the search for mechanisms related to both the functions of sirtuins and their application in the treatment of metabolic disorders. Identification of novel potential targets in hAT-MSCs during altered glucose homeostasis may be of clinical relevance in the modulation of functional activities of hAT-MSCs in patients with various metabolic disorders.
Footnotes
Acknowledgments
This work was supported by the Bio-Scientific Research Grant funded by the Pusan National University (PNU-2008-101-102).
Author Disclosure Statement
No competing financial interests exist.