
Abstract
A theoretical inverse relationship exists between osteogenic (bone forming) and adipogenic (fat forming) mesenchymal stem cell (MSC) differentiation. This inverse relationship in theory partially underlies the clinical entity of osteoporosis, in which marrow MSCs have a preference for adipose differentiation that increases with age. Two pro-osteogenic cytokines have been recently studied that each also possesses antiadipogenic properties: Sonic Hedgehog (SHH) and NELL-1 proteins. In the present study, we assayed the potential additive effects of the biologically active N-terminus of SHH (SHH-N) and NELL-1 protein on osteogenic and adipogenic differentiation of human primary adipose-derived stromal cell (hASCs). We observed that both recombinant SHH-N and NELL-1 protein significantly enhanced osteogenic differentiation and reduced adipose differentiation across all markers examined (alkaline phosphatase, Alizarin red and Oil red O staining, and osteogenic gene expression). Moreover, SHH-N and NELL-1 directed signaling produced additive effects on the pro-osteogenic and antiadipogenic differentiation of hASCs. NELL-1 treatment increased Hedgehog signaling pathway expression; coapplication of the Smoothened antagonist Cyclopamine reversed the pro-osteogenic effect of NELL-1. In summary, Hedgehog and Nell-1 signaling exert additive effects on the pro-osteogenic and antiadipogenic differentiation of ASCs. These studies suggest that the combination cytokines SHH-N+NELL-1 may represent a viable future technique for inducing the osteogenic differentiation of MSCs.
Introduction
M
Both Sonic Hedgehog (SHH) and Nell-1 have long been recognized as possessing osteoinductive properties. The activity of Hedgehog ligands resides in the N-terminus, and there are 3 known mammalian hedgehog ligands: sonic, Indian, and desert hedgehog. As the most studied, the N-terminal SHH (SHH-N) has been shown to have pro-osteogenic and antiadipogenic effects in numerous cell types, including adipose-derived stromal cells (ASCs) [12,14 –16]. Developmentally, Hedgehog ligands have been shown to be critical in skeletal ossification. For example, Indian Hedgehog (Ihh) null mice have a severe phenotype including foreshortened limbs, near absence of a cranial base, and cranial bones of diminished size [17 –20]. Overall, studies in the field of bone tissue engineering have suggested that the manipulation of SHH in ASCs may be of potential future therapeutic benefit [12,21 –23].
The growth factor NELL-1 has long been recognized as having osteoinductive properties. Similar to Hedgehog, NELL-1 is significant in skeletal development; NELL-1 is overexpressed in prematurely fusing (or craniosynostotic) calvarial sutures [24]. Follow-up studies demonstrated that Nell-1 overexpressing mice exhibit calvarial bone overgrowth and cranial suture fusion [25]. On the contrary, animals deficient in functional Nell-1 protein exhibit widespread skeletal anomalies [26,27]. Similar to Hedgehog signaling, NELL-1 has been observed as having potential utility from a bone tissue-engineering perspective, having induced bone formation in both small and large mammalian models [28 –32]. Recently, Nell-1 signaling has been shown to be antiadipogenic [33]. Moreover, recent evidence has found that NELL-1 can reverse the pro-adipogenic effects of high-dose BMP2 both in vitro and in vivo (data in submission).
In the present study, we investigate the potential additive effects of SHH and NELL-1 signaling in human primary ASCs (hASCs) in terms of both osteogenic and adipogenic differentiation. In our previous publication [33], we found that Nell-1 signaling positively regulates Shh transcript abundance as well as Hedgehog signaling activity in 3T3-L1 preadipocytes. We hypothesized that the 2 cytokines together may have additive effects in human primary ASCs.
Materials and Methods
Cell isolation
ASCs were isolated from human adult lipoaspirate as previously described [33]. Fat tissues were obtained from n=5 patients without medical comorbidities. The cell yield and cell viability were assessed via Trypan blue staining. These values are presented as cell yield per 100 cubic centimeters (cc) of lipoaspirate, viable cell yield per 100 cc lipoaspirate, and percentage cell viability (Table 1). Human ASCs were expanded in 100 mm dishes for subsequent experiments.
Flow cytometry analysis
Cultured cells were labeled with the following commercial antibodies: anti-CD44-PerCP (BD Bioscience; 1:100), anti-CD73-PE (BD Biosciences; 1:100), anti-CD90-APC (BD Biosciences; 1:100), anti-CD105-PE (BD Biosciences; 1:100), anti-CD45-FITC (BD Biosciences; 1:100), or anti-CD31-APC (Biolegend; 1:100). Cells were then washed twice, and at least 50,000 events were acquired on an FACSCanto II cytometer (BD Biosciences). Unstained cells were used as a negative control. ASCs from n=2 patient samples were analyzed at passage 4.
Bromodeoxyuridine incorporation
Proliferation was assessed by bromodeoxyuridine (BrdU) incorporation assays as previously described [34], exposed to recombinant human (rh) SHH-N (250 and 500 ng/mL; R&D Systems) and rhNELL-1 (100 and 300 ng/mL). Dosages were used based on previous publications [12,33]. BrdU incorporation was quantified using a photometric ELISA (Roche Applied Science). n=6 per treatment group, experiments were performed from n=3 patient samples.
Osteogenic differentiation and assessments
Osteogenic differentiation and assessments were performed using hASCs as previously described [35]. rhSHH-N (25, 50, 250 and 500 ng/mL) and rhNELL-1 (25, 50, 100, and 300 ng/mL) were supplemented to osteogenic differentiation medium (ODM) and were replenished every 3 days. For select experiments, the Smoothened antagonist Cyclopamine was added (5 μM). To assess early osteogenesis, alkaline phosphatase (ALP) staining was performed as per the manufacturer's instructions (Sigma Aldrich; Cat. No. 864-1kt) on day 5 of differentiation; quantification was performed in each case normalized to total protein content in sister wells [35]. To assess bone nodule formation, Alizarin red (AR) staining and quantification was performed as previously described on day 10 of differentiation; quantification was performed by cetylpyridium chloride leaching and photometric quantification, normalized to total protein content [36]. A real-time polymerase chain reaction (PCR) was performed as previously described, performed in triplicate wells per RNA isolate [37]. Specific human runt-related transcription factor 2 (RUNX2) and osteocalcin (OCN) genes and primer sequences are listed in Table 2.
OCN, osteocalcin; PTC1, Patched1; RUNX2, runt-related transcription factor 2; SHH, Sonic Hedgehog.
In vitro adipogenic differentiation
For adipogenic differentiation, hASCs were seeded in 12-well plates at a density of 50,000 cells per well. rhSHH-N (250 and 500 ng/mL) and rhNELL-1 (100 and 300 ng/mL) were supplemented to adipogenic differentiation medium (ADM) and were replenished every 3 days. All assays were performed in triplicate wells. After attachment, cells were treated with ADM (Stem Cell Technologies, Inc.; Cat. No. 05501&05503). All experiments were performed in biological triplicate (cells derived from 3 separate patients), and each assay was performed in triplicate wells (n=3 wells per cell type per experiment, n=3*3=9 samples per condition). Assessments of adipogenic differentiation were performed as previously described [12,36,38]. Oil red O (ORO) staining was performed using a 0.2% solution in 60% isopropanol/40% deionized water.
Statistical analysis
Means and standard deviations were calculated, as depicted in the figures and figure legends. Bar graphs represent means, whereas error bars represent 1 standard deviation. Statistical analysis was performed using the appropriate ANOVA when more than 2 groups were compared, followed by a post hoc Student's t-test. *P≤0.01 was considered significant.
Results
Cell yield, proliferation, and cell surface expression markers of ASCs
Human primary ASCs were isolated and cultured from 5 donors after cosmetic liposuction procedures. First, the patient demographics, cell yield per unit volume, and cell viability were assessed across patient samples (Table 1). All patients were adults in good health, ranging in age from 28 to 59, and most were women (3/5 patients). In terms of cell yield, a narrow range of values was observed across samples. For example, an only 3.25-fold difference was observed from lowest to highest cell yields (0.8×107 to 2.6×107 viable cells/100 cc lipoaspirate).
Next, baseline characteristics of proliferation, osteogenic differentiation, and cell surface expression were assessed between patient samples (Fig. 1). Cell proliferation was assessed by BrdU incorporation (Fig. 1A). Overall, BrdU incorporation was relatively similar across n=3 patient samples (Fig. 1A). Next, osteogenic differentiation was assessed by ALP staining for early differentiation, and AR staining for bone nodule formation (Fig. 1B). Only subtle differences were observed between n=3 patient samples. Finally, cell surface markers were assessed by flow cytometry (Fig. 1C). As shown, ASCs express 100% of typical MSC markers, including CD73, CD44, CD90, and CD105. In contrast, ASCs were negative for CD45 (a hematopoietic cell marker) and CD31 (an endothelial cell marker). Similar findings of a conventional MSC phenotype were observed across n=2 patient samples. Thus, hASCs from distinct donors showed relatively similar baseline characteristics of (1) cell yield and viability, (2) growth, (3) differentiation, and (4) MSC surface markers.

Baseline phenotype of human primary adipose-derived stromal cell (hASCs).
NELL-1 and SHH-N effects on proliferation
Next, the distinct effects of NELL-1 and SHH-N on BrdU uptake were assayed (Fig. 2). Dosages were used based on previous publications [12,33]. In agreement with other cell types [25,39], NELL-1 showed no effects on hASC proliferation. In contrast to NELL-1 and in agreement with previous publications [12], hASCs showed a slight mitogenic response to SHH-N. Thus, (1) hASC populations showed similar effects in cell proliferation on cytokine stimulation, and (2) the effects of SHH-N and NELL-1 on hASC proliferation were divergent.
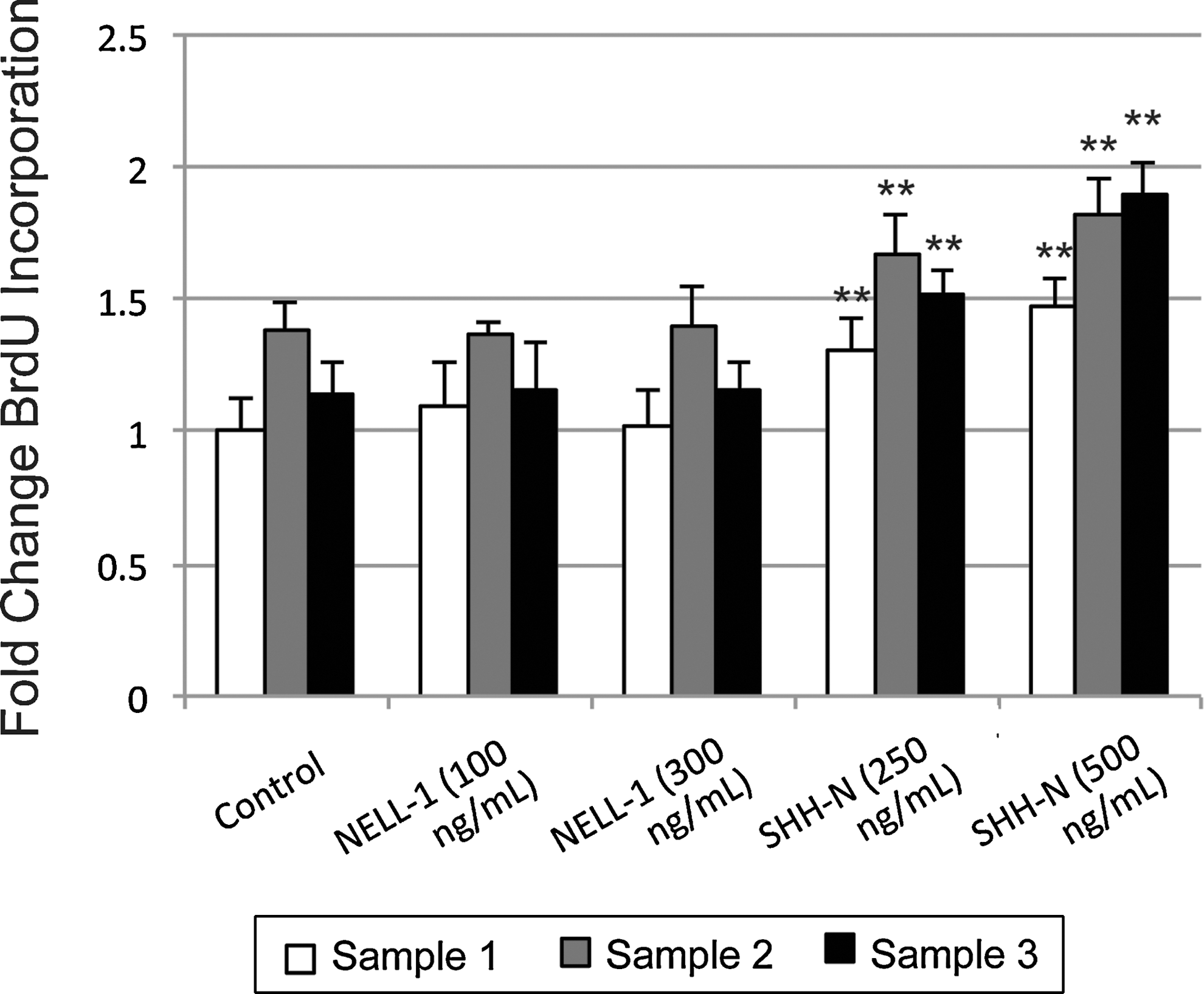
Cellular proliferation with NELL-1 or N-terminus of Sonic Hedgehog (SHH-N). BrdU incorporation assays at 4 days comparing baseline proliferation (control) with the effects of NELL-1 (100 and 300 ng/mL) or SHH-N (250 and 500 ng/mL). **P<0.01 in reference to identical sample under control conditions.
Endogenous Nell-1 and Hedgehog activity during osteogenic differentiation
We next sought to confirm that both NELL-1 and SHH signaling could be detected in hASCs, and whether endogenous expression levels would change over time in ODM. No recombinant proteins were added to standard ODM; gene expression was evaluated at 0 and 7 days differentiation. NELL-1 expression increased over time in all patient samples; however, the change in sample 3 was not statistically significant (Fig. 3A). SHH expression increased in all 3 patient samples, the fold change being greater than that of NELL-1 (Fig. 3B). Likewise, Patched1 (PTC1), a transmembrane receptor for Hedgehog ligands and a reporter for Hedgehog signaling activity, increased over time in differentiation medium (Fig. 3C). Thus, (1) hASCs from distinct patient specimens had similar changes in Nell-1 and Hedgehog gene expression over time in ODM, and (2) both signaling pathways increased in transcript abundance over time, with Hedgehog signaling showing a more robust increase.
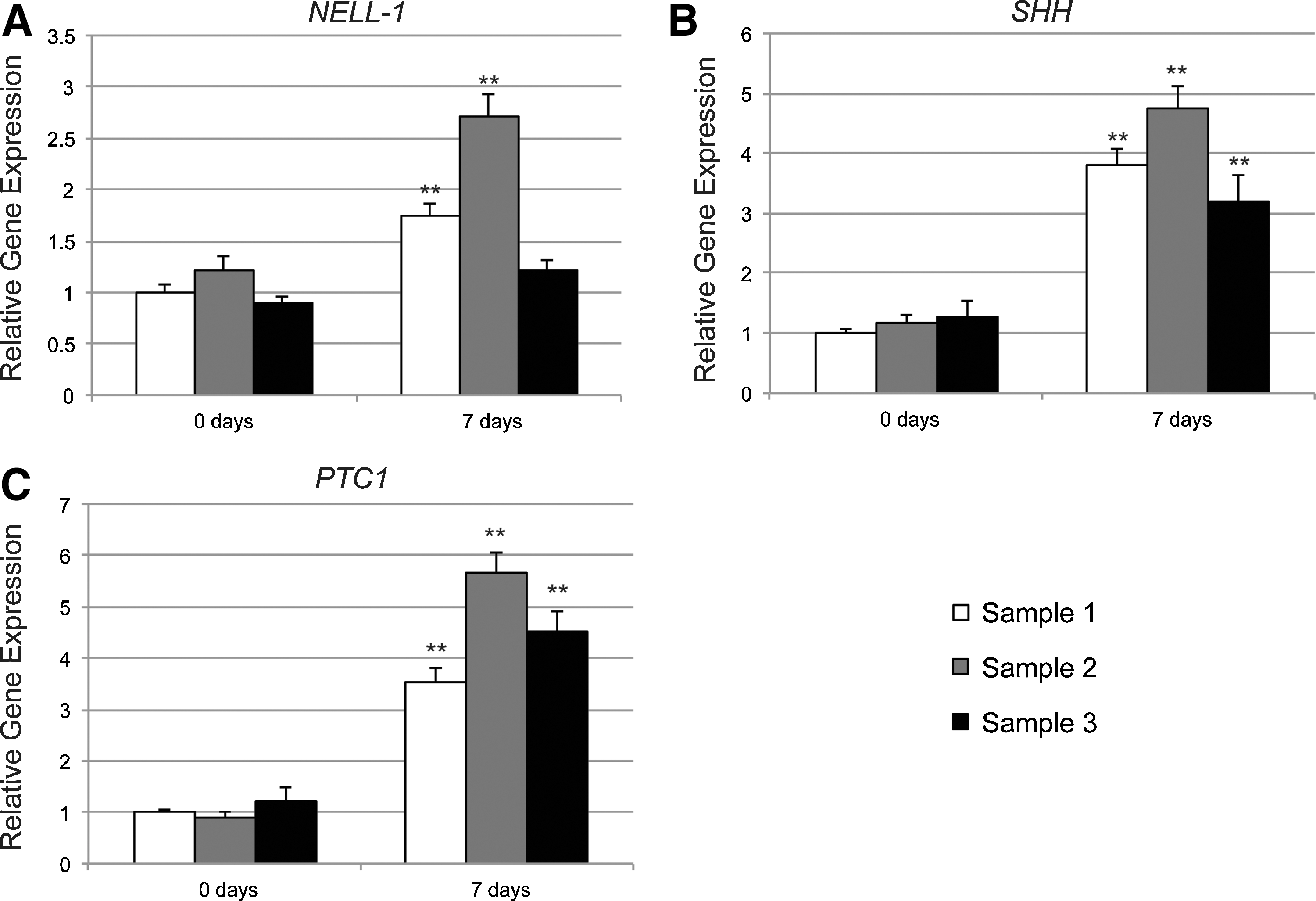
Endogenous Nell-1 and Hedgehog signaling. Human primary ASCs from 3 patient samples were assayed for Nell-1 and Hedgehog specific gene expression after 0 and 7 days of osteogenic differentiation by quantitative real time-polymerase chain reaction (PCR).
NELL-1 and SHH-N effects on osteogenic differentiation
While the positive effects of either NELL-1 or SHH-N alone on ASC osteogenic differentiation have been previously reported [12], the additive effects of both cytokines combined have not been addressed. After 5 days differentiation, ALP staining and quantification was performed, indicative of early osteogenic differentiation (Fig. 4A, B). Results showed that both NELL-1 and SHH-N, or SHH-N alone increased ALP activity. Moreover, the combination NELL-1+SHH-N showed a significant increase over either alone. Next, after 10 days differentiation, AR staining was performed to assay terminal differentiation (bone nodule formation) (Fig. 4C, D). In agreement with ALP activity, AR staining and quantification showed an additive increase with the combination treatment NELL-1+SHH-N. In parallel, osteogenic specific gene expression was assayed at 5 and 10 days differentiation (Supplementary Fig. S1; Supplementary Data are available online at

Osteogenic differentiation with NELL-1 and/or SHH-N. Recombinant human (rh) NELL-1 (100 and 300 ng/mL) or SHH-N (100 and 250 ng/mL) were supplemented to standard ODM.
Low-concentration NELL-1 and SHH-N effects on osteogenic differentiation
In earlier dose curves, high concentrations of both NELL-1 and SHH-N had a pro-osteogenic effect alone. For example, a 75%–230% increase in mineralization was detected with the highest concentration NELL-1 (300 ng/mL) or SHH-N (250 ng/mL). We next examined the effects of much lower concentrations of cytokine under osteogenic differentiation conditions, either alone or in combination (Supplementary Fig. S2, 25–50 ng/mL each). Differentiation was again assessed by ALP staining and quantification (Supplementary Fig. S2A, B), and bone nodule formation was assessed by AR staining and quantification (Supplementary Fig. S2C, D). Overall, at all low concentrations, neither cytokine showed a consistent increase in differentiation reaching statistical significance. Exceptions include NELL-1 alone (50 ng/mL), which increased ALP quantification by 21% (P<0.01), and SHH-N alone (25 and 50 ng/mL), which increased AR quantification by 34%–37% (P<0.01). In general, however, the combination of low-concentration NELL-1+SHH-N led to a more consistent, statistically significant increase in comparison to either cytokine alone. This was most clear in ALP staining, where a slight dose-dependent increase in ALP staining was observed, with NELL-1+SHH-N (50 ng/mL each) leading to the highest 37% increase over control (P<0.01). Thus, (1) low concentrations NELL-1 and SHH-N alone lead to inconsistent increases in ASC osteogenic differentiation, and (2) the combination therapy NELL-1+SHH-N at low-concentrations showed more consistent pro-osteogenic effects.
Pro-osteogenic effects of NELL-1 are reversed by Hedgehog signaling inhibition
In our previous publication [33], we found that Nell-1 signaling positively regulates Shh transcript abundance as well as Hedgehog signaling activity in 3T3-L1 preadipocytes. First, we examined the extent to which NELL-1 increased Hedgehog signaling activity by measuring the receptor and reporter gene PTC1 (Supplementary Fig. S3). After 3 days, NELL-1 increased PTC1 transcript abundance in n=2 hASC samples. Next, could the pro-osteogenic effects of NELL-1 be antagonized by the addition of a Hedgehog signaling antagonist? To answer this question, Cyclopamine, a Smoothened antagonist, was added either alone or in combination with NELL-1 during osteogenic differentiation conditions (Fig. 5). Osteogenesis was assessed by ALP staining and quantification (Fig. 5A, B), AR staining and quantification (Fig. 5A, C), and RUNX2 expression as assessed by quantitative real time (RT)-PCR (Fig. 5D). By all markers, we confirmed again that NELL-1 showed a pro-osteogenic effect alone (black as compared with white bars, Fig. 5B–D). In contrast, the addition of cyclopamine to NELL-1 reversed the pro-osteogenic effect (gray bars, Fig. 5B–D). Thus, (1) NELL-1 treatment is associated with an increase in Hedgehog signaling activity in hASCs, and (2) NELL-1's pro-osteogenic effect can be fully antagonized by a Hedgehog inhibitor.

Osteogenic differentiation with NELL-1 and/or cyclopamine. rhNELL-1 (300 ng/mL) or the smoothened antagonist cyclopamine (5 μM) was supplemented to standard ODM.
NELL-1 and SHH-N effects on adipogenic differentiation
Finally, the converse experiments in ADM were next performed, with the addition of NELL-1 and/or SHH-N. Either cytokine alone resulted in diminished adipogenic differentiation as assessed by lipid droplet content (ORO staining) (Fig. 6A). Quantification revealed a similar reduction in ORO staining intensity when normalized to total protein content (Fig. 6B). The combination treatment of NELL-1+SHH-N resulted in a significant reduction in lipid accumulation in comparison to each cytokine treatment alone (Fig. 6A, B).

Adipogenic differentiation with NELL-1 and/or SHH-N. rhNELL-1 (100 and 300 ng/mL) or SHH-N (100 and 250 ng/mL) was supplemented to standard adipogenic differentiation medium.
Discussion
Numerous multipotent cell types exist for tissue engineering applications, including traditional bone marrow MSCs and ASCs among others. Benefits for the use of human ASCs include that this cell type is easily obtainable by simple liposuction procedures and is readily expandable in vitro as compared with bone marrow mesenchymal cells [40,41]. Previous authors have convincingly observed that ASCs, whether derived from mouse or human origin, contribute to the osseous healing of mouse cranial defects [42,43]. Here, we sought to examine whether SHH and Nell-1 signaling would additively enhance hASC osteogenic differentiation for future tissue engineering applications. In fact, we found this to be the case: the combination therapy SHH-N+NELL-1 proved to enhance osteogenic differentiation in vitro over either cytokine alone. We also have provided evidence that the pro-osteogenic effects of NELL-1 can be abrogated by Hedgehog signaling inhibition.
It is interesting to note that both Nell-1 and Hedgehog ligands have strong associations with cranial sutures: the thin strips of tissue mesenchyme that separate cranial bones. Hedgehog signaling activity in the calvaria was first described as potentially due to SHH [44]; however, more detailed later studies have since then attributed sutural Hedgehog activity to be Ihh dependent [17,19,45]. Loss of Ihh in the mouse (Ihh null animals) results in a global reduction in expanse, thickness, and degree of mineralization of intramembranous cranial bones [17]. Interestingly, loss of function of Nell-1 in the mouse results in a strikingly similar calvarial phenotype with reduced bone thickness and increased sutural width due to defective mineralization [46]. These loss-of-function experiments suggest partially overlapping roles for both Hedgehog and Nell-1 signaling in intramembranous bone ossification.
While not addressed in the present study, it is important to note that both Hedgehog and Nell-1 signaling have been well studied in not only osteoblast but also chondrocyte development and differentiation. In fact, Ihh expression was first understood as important in skeletogenesis, as it is detected in chondrocytes of the early cartilage condensation, and later in the transition region from proliferating to hypertrophic cells [20,47,48]. Ihh regulates the rate of chondrocyte differentiation by paracrine induction of parathyroid hormone-related protein (PTHrP) [19,20,49], forming a negative feedback mechanism that serves to regulate chondrocyte hypertrophy. Recombinant NELL-1 has been shown to have stimulatory effects on chondrocyte proliferation and cartilage-specific extracellular matrix deposition in vitro, as well as in maintaining the differentiated state of rabbit chondrocytes [50]. Moreover, when placed in a rabbit articular cartilage defect, NELL-1 increases cartilage repair and regeneration [51]. In future studies, it would indeed be interesting to examine the potential additive effects of SHH-N+NELL-1 on hASC chondrogenic differentiation.
In summary, Hedgehog and Nell-1 signaling exert additive effects on the pro-osteogenic and antiadipogenic differentiation of human ASCs. These studies suggest the future therapeutic utility of modulating both Sonic and Nell-1 together for either skeletal or soft-tissue regenerative medicine.
Footnotes
Acknowledgments
This work was supported by the CIRM Early Translational II Research Award TR2-01821, NIH/NIDCR (grants R21 DE0177711 and RO1 DE01607), UC Discovery Grant 07-10677, Eli & Edythe Broad Center of Regenerative Medicine and Stem Cell Research at UCLA Innovation Award, a T32 training fellowship (5T32DE007296-14) to A.W.J., and a CIRM training fellowship (TG2-01169) to J.N.Z. and M.C.
Author Disclosure Statement
Drs. X.Z, K.T., and C.S. are inventors of NELL-1 related patents. Drs. X.Z, K.T., and C.S. are founders of Bone Biologics Inc., which sublicenses NELL-1 patents from the UC Regents.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.