
Abstract
The earthworm, Eudrilus eugeniae, has a prodigious ability to regenerate lost segments. The skin of the worm has an outermost epidermal layer followed by a thick circular muscle layer and an innermost thin longitudinal cell layer. During the process of regeneration, the circular muscle layer decreased in thickness, and longitudinal cell layer increased. The histological analysis of the regenerated worm shows that the longitudinal cell layer forms the regeneration blastema. BrdU-labeling retention assay confirmed that the circular muscle and longitudinal cell layers have BrdU-positive cells, which migrate from the adjacent segments to the regeneration blastema. In addition, it was noted that the cells of the earthworm, E. eugeniae, have the property of autofluorescence. Autofluorescence was found in the cytoplasm, but not in the nucleus. It has been also found that the major source for autofluorescence is riboflavin. Further, it was also demonstrated that supplementation with riboflavin increases the rate of regeneration, while regeneration was hampered by reduced levels of riboflavin. The importance of riboflavin in regeneration was also confirmed by rescue assay. In addition, it was also identified that BrdU-positive cells are highly fluorescent compared to the surrounding cells.
Introduction
T
In animals, autofluorescence has been reported in the scorpion [5], spider [6], butterflies [7], jellyfish [8], frog [9], and many sea animals [10]. Chemical substances such as coumarin [5] or the modified amino acid dityrosine [11] have been reported as sources of autofluorescence in living systems. In humans, autofluorescence has been reported in the cytoplasm of the human hair follicle stem cell [12]. The fluorophore responsible for autofluorescence in the stem cell is not yet known.
Many animals have been used to study regeneration and stem cell biology. The salamander has been shown to regenerate its limb, tail, upper and lower jaws, ocular tissues such as the lens and retina, the intestine, and small sections of the heart [13 –16]. In axolotl, the transplantation of blastema from the amputated limb to the anterior chamber of the eye or a tunnel bored in the connective tissue of the dorsal fin leads to normal regeneration [17,18]. It was also reported that cells retain the memory of their tissue origins during limb regeneration in axolotl [19]. Zebrafish has the capacity for regeneration and renews the myocardium vigorously and restricts scar formation [20,21]. Axon regeneration in Caenorhabditis elegan reveals the regeneration of several neuron types, including motor and sensory neurons, upon injury [22]. The process of regeneration, from the planaria to amphibians, starts with the formation of regeneration blastema, which is a proliferative mass of undifferentiated progenitor cells from which new differentiated cells arise [23,24]. The earthworm has a prodigious regeneration capacity, and studies on regeneration have been reported in earthworms such as Eisenia andrei, Eisenia fetida, Lumbricus rubellas [25 –29], Ptychodera flava [30], and Enchytraeus japonensis [31]. In E. japonensis, regeneration is controlled by the nervous system [32]. In addition, it has been reported that in most of the oligocheate worms, a common morphological feature of the neoblast is the presence of undifferentiated cell types having a high nucleo-cytoplasmic ratio, a large nucleus with a large nucleolus and a basophilic cytoplasm [33 –37]. Even though the earthworm is easy, economical to rear, and a convenient model system, the process of regeneration has not been worked out in detail.
In the present study, it has been found that the longitudinal cell layer in the skin forms the regenerative blastema. The BrdU-labeling retention assay revealed that BrdU-positive cells are present in the boundary of the circular muscle layer close to the epidermis and in the longitudinal cell layer. The BrdU-positive cells migrated into the blastema during regeneration. Interestingly, it was found that these cells have a strong fluorescent property compared with the surrounding cells, and the major source of fluorescence is riboflavin. Riboflavin is important for normal regeneration of the earthworm.
Materials and Methods
Culture and maintenance of earthworm
The earthworm, E. eugeniae, was maintained in a tub containing soil, cow dung, and leaf litter at an ambient temperature [38].
Autofluorescence of coelomic fluid cells
To observe the autofluorescence of coelomic fluid cells, coelomic fluid was collected from the mature worm. It was smeared on a clean glass slide. The smear was fixed with 4% paraformaldehyde, briefly washed with 1×PBS, and examined under a Nikon Ti-S inverted fluorescence microscope.
Histology
To study autofluorescence in the worm and the patterns of tissue formation during regeneration, histology was performed. The worm tissues were fixed with 10% formalin for 24 h and allowed to dehydrate (by gradient isopropyl alcohol from 60% to 100%), followed by clearing of the isopropyl alcohol with xylene, and then embedded in paraffin wax (Purchased from HiMedia Laboratories Private Limited). The block was sliced into 6-μm-thick sections using a microtome (Besto), and the sections were mounted on slides and viewed under a Nikon Ti-S inverted fluorescent microscope for autofluorescence, after which the slides were stained with a hematoxylin–eosin combination. Then, the staining pattern was documented. Solvents used for histology were purchased from Merck Specialities Private Limited, India.
Sample preparation for thin-layer chromatography
Earthworm tissues were homogenized with sterile distilled water. The homogenates were centrifuged at 5,000 rpm for 10 min (Eppendorf Mini Spin Centrifuge), and the supernatant was resolved by thin-layer chromatography.
Thin-layer chromatography
The slurry was prepared by mixing silica gel and water in the ratio 3:2. It was applied on the glass plate at a thickness of about 0.25 mm and the plate was allowed to dry at room temperature for 15–30 min. Then, the plate was kept in a hot-air oven at 100°C–120°C for 2 h [39]. Samples were loaded on the plate, and resolved with a solvent mixture [butanol (7):chloroform (4):acetic acid (5):ammonia (1):water (1)] and visualized under UV light. The thin layer chromatography (TLC) plate was further stained with ninhydrin. The spot visualized under UV was scraped off using a surgical blade and the scraped material was transferred to a microfuge tube. For eluting the fluorophore and riboflavin from the silica gel, distilled water was added and mixed well. The tube was then centrifuged at 10,000 rpm for 10 min, and the supernatant was used for thin-layer chromatography and spectrofluorometric analysis. Solvents used for the TLC were purchased from Merck Specialities Private Limited, India.
Estimation of riboflavin
The known concentration of riboflavin was resolved using thin-layer chromatography and visualized under UV light. By using imaging software NIS-Elements BR 3.1, the gray-scale intensity was measured. From the data, a standard graph was generated and the concentration of riboflavin in the different samples of worm was determined.
A solution of riboflavin in distilled water was scanned using a spectrophotometer and the absorbance maxima were determined. A standard graph was then plotted by taking different concentrations of riboflavin. Using the standard graph, unknown concentrations of riboflavin in worm samples were calculated.
Spectrofluorometric analyses
Spectrofluorometric analyses were performed using the lysate of earthworm. Earthworm tissues were homogenized with sterile distilled water, and the lysate obtained was centrifuged at 5,000 rpm for 10 min (Eppendorf Mini Spin Centrifuge). The supernatant was collected, and it was subjected for spectrofluorometric analyses with standard riboflavin, flavin mononucleotide (FMN), and flavin adenine dinucleotide (FAD) (purchased from HiMedia Laboratories Private Limited).
BrdU labeling retention assay
The BrdU (purchased from Sisco Research Laboratories Private Limited) stock solution was prepared at a concentration of 10 mg/mL in 1×PBS and each worm was injected with 10 μL of BrdU from the 20th segment to the 24th segments. The worm injected with BrdU was maintained for 30 days for chasing. On the 30th day the worm was amputated at the 11th segment and allowed to regenerate the lost part. The regeneration blastema appeared on the 4th day, and it was formalin-fixed, paraffin-embedded, sectioned, and stained with anti-BrdU antibody.
Immunohistochemistry
To visualize BrdU incorporation, paraffin-embedded earthworm tissue sections (6 μm) were de-paraffinized with xylene and hydrated. Endogenous peroxidase was inhibited by incubation of the sections for 30 min with freshly prepared 10% H2O2 and 10% methanol in 1×PBS. The sections were then treated with 0.1% trypsin in 0.1% CaCl2 at 37°C for 10 min. DNA was denatured by incubating the section with 2N HCL at 37°C for 45 min. Nonspecific staining was blocked by treatment with 2% BSA for 1 h at ambient temperature. The sections were then incubated overnight at 4°C with mouse monoclonal anti-BrdU antibody (Clone BU-33; Sigma) at a dilution of 1:200 in BSA. After incubation with primary antibody, tissue sections were washed and incubated for 1 h with goat anti-mouse IgG conjugated with horseradish peroxidase (Lot No: 062100; GeNei) at a dilution of 1:500. Staining was developed with Diaminobenzidine (DAB Kit; GeNei) substrate and the sections were counterstained with Ehrlich hematoxylin. The prepared slides were mounted with DPX and observed under a Nikon Ti-S microscope.
Statistical analysis
Statistical analyses such as standard deviation, P value for BrdU-positive cells versus a positive control, and regeneration growth kinetics were performed using Microsoft Office Excel 2007.
Results
The earthworm, E. eugeniae, is shown in Fig. 1A. The clitellum of the worm begins at the 13th segment and ends at the 18th [2]. The segments upstream (from the 1st to the 13th) and downstream (from the 18th to the last) of the clitellum are called the anterior and posterior parts, respectively.
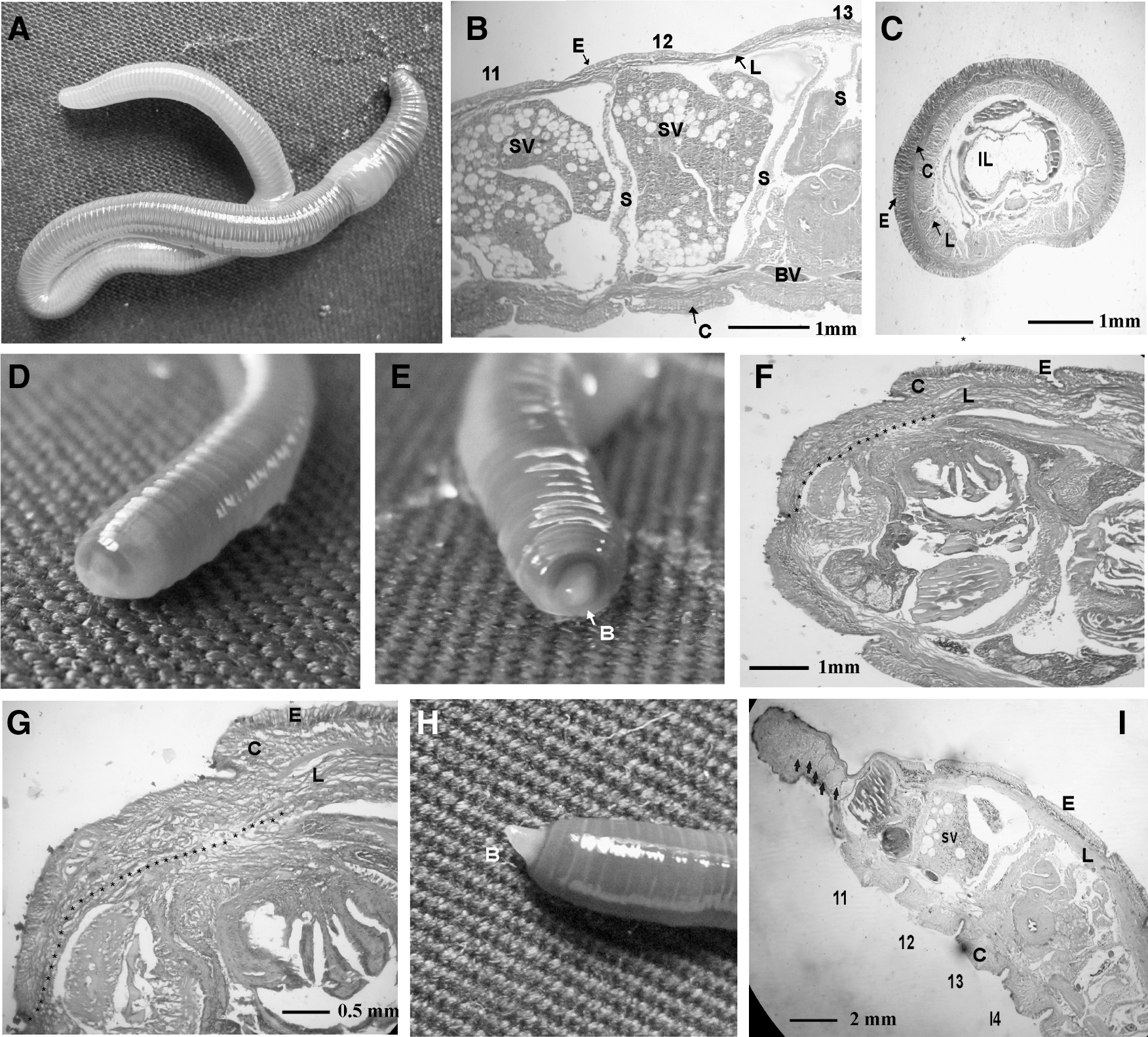
Regeneration studies in the earthworm, Eudrilus eugeniae.
To study the regeneration process, 6 worms were taken and amputated at the junction of the 10th and 11th segments. The first 10 segments (1–10) contain the important organs as described earlier. The amputated worms were maintained in a rearing bed, and it took 24 h for the wound to heal at the posterior part of the worms (Fig. 1D). The regeneration blastema was formed at the 48th hour in all the worms (Fig. 1E).
The normal anterior segments (segments 1–10) and the regenerating blastema were subjected to histology analysis (Fig. 1B, C) and (Fig. 1F, G). The pattern (Fig. 1B, C) shows that the worms have an outermost epidermal layer followed by a circular muscle layer. The innermost layer in the skin is longitudinal cells. These layers are shown clearly in the longitudinal (Fig. 1B) and transverse section (Fig. 1C) of the worms, indicating that all 3 layers (epidermal, circular, and longitudinal layers) are not even in the body of the earthworm. They are thicker in the dorsal side and their thickness gradually decreases toward the ventral side, as shown in Fig. 1C.
During regeneration, there was no change in the epidermal layer. In contrast, the thickness of the circular muscle layer reduced from 170±3 to 98±2 μm at the mid-part of the dorsal side. In contrast, the longitudinal cell layer, which was 100±2 μm thick in normal worms, increased in thickness to 188±3 μm during regeneration. In addition, the section of the worm at 48 h of regeneration shows that the regeneration blastema is formed by the longitudinal cells (Fig. 1F, G).
In the 7th day blastema shown in Fig. 1H, the formation of septum (Fig. 1I) (indicated by the arrow) was observed. Based upon the data it is concluded that the regeneration blastema is formed by longitudinal cells, and that these cells divide further to produce different types of cells to regenerate the lost segments.
BrdU-labeling retention assay was performed in order to locate the stem cells. Six worms were injected with BrdU (100 μg/g of worm) into the ceolomic fluid from the 20th segment to the 24th. The BrdU-injected worms were maintained carefully for 30 days. On the 30th day, the worms were amputated at the junction of the 10th and 11th segments. The regenerated blastema of the 4th day was subjected to sectioning, followed by immunohistochemistry with the anti-BrdU antibody and a brief staining with hematoxylin. The data (Fig. 2A) show the presence of a cluster of BrdU-positive cells in the blastema. These cells are shown in the boxes of Fig. 2A. The regions containing the BrdU-positive cells were documented with the 40× objective (Fig. 2B–E). The data clearly show that the BrdU-positive and surrounding cells are stained dark brown and blue (by hematoxylin), respectively. The data show that the mass of slow-proliferating BrdU-positive cells is surrounded by fast-proliferating cells that are BrdU-negative. To study the presence of BrdU-positive cells in the upstream segments of the blastema, segments 11–13 were sectioned and processed for immunohistochemistry with the anti-BrdU antibody. The data (Fig. 2F) clearly show that BrdU-positive cells are present in the circular muscle layer, being particularly abundant in the circular muscle layer at the base of the epidermis. At the same time, a few BrdU-positive cells are also found in the longitudinal layer shown and marked by boxes in Fig. 2F. To study BrdU-positive cells in the normal worm, 3 worms were injected with BrdU (100 μg/g of worm) into the ceolomic fluid from the 20th segment to the 24th. These worms were maintained carefully for 34 days, after which they were processed for immunohistochemistry using the anti-BrdU antibody. The data from this study are shown in Fig. 2G, which clearly reveals the presence of a higher population of BrdU-positive cells in the regenerating worm (Fig. 2F) than in the normal worm (Fig. 2G). To verify these results, the experiments were repeated thrice and cells in the different layers of the skin and BrdU-positive cells were counted in each mid-part section of the 11th and 12th segments of regenerating and normal worms (Supplementary Table S1; Supplementary Data are available online at
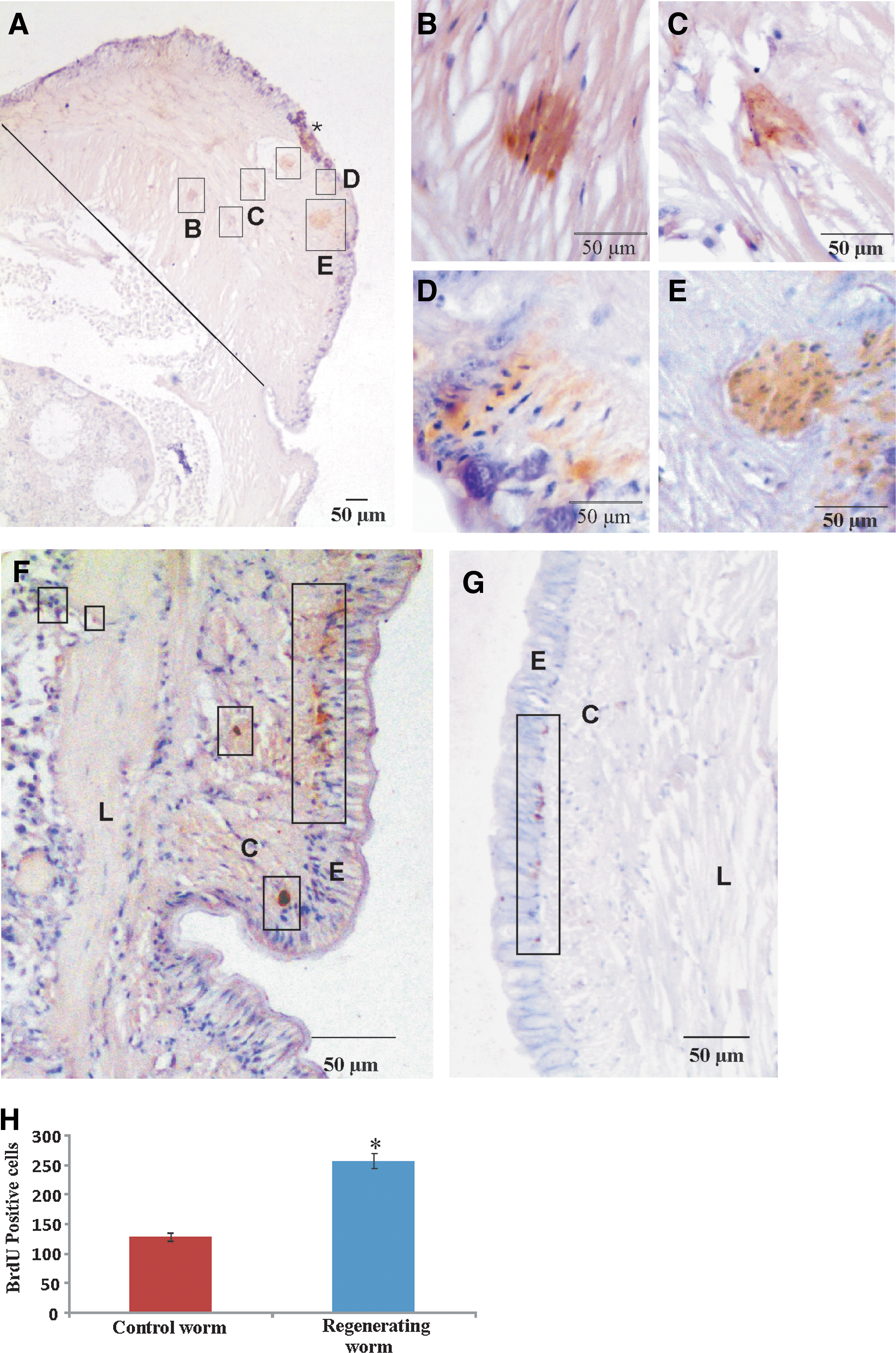
BrdU labeling retention assay.
As a negative control, earthworm tissue sections were processed with control anti-mouse serum and data from this experiment are shown in Supplementary Fig. S1. The data clearly show the absence of any reaction involving DAB, confirming that the earlier data with the BrdU antibody (Figs. 2 and 3E) are, in fact, signals corresponding to the reaction of DAB with the anti-BrdU antibody.
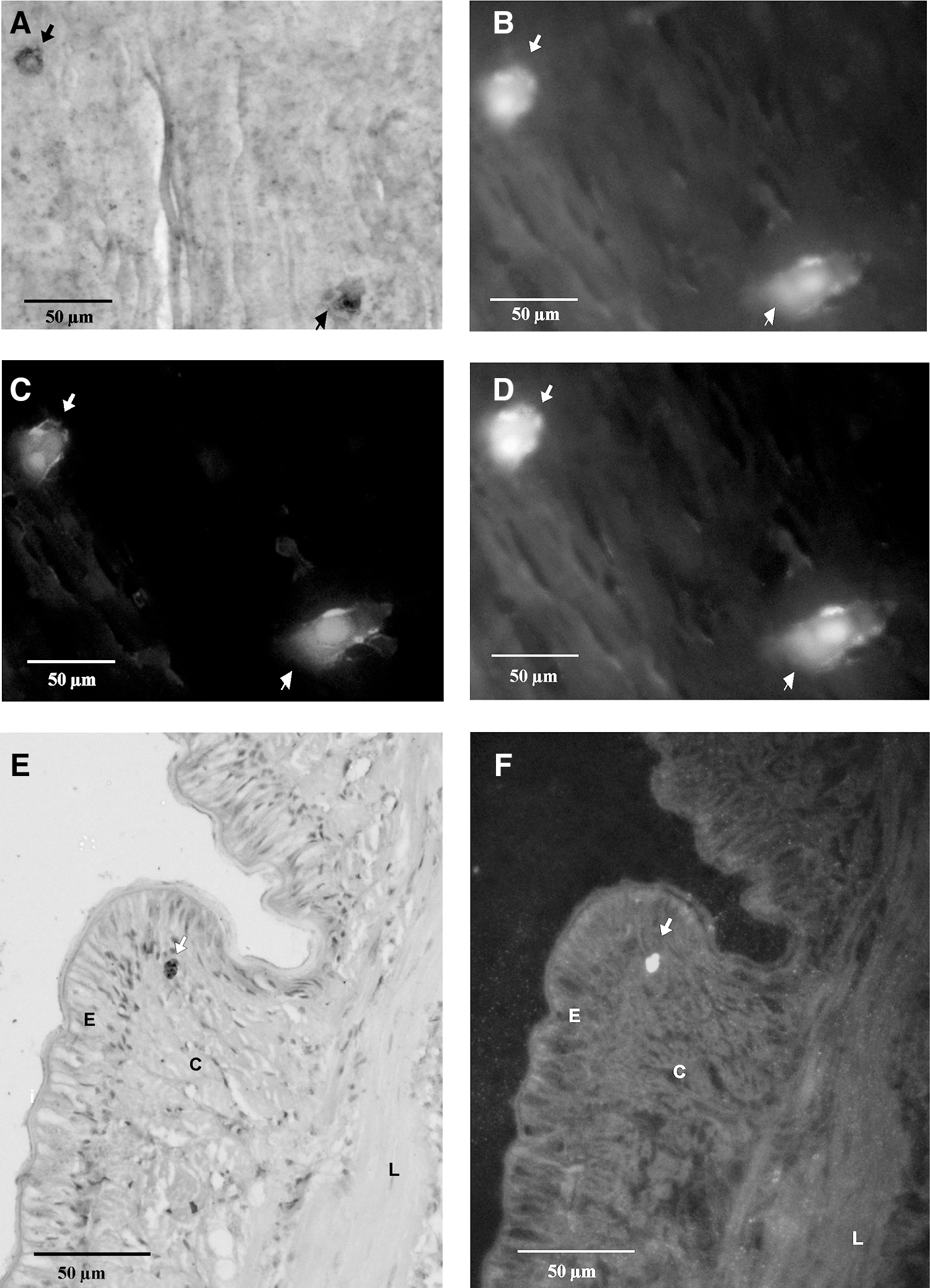
Autofluorescence of BrdU-positive cells.
It has been reported that stem cells of the human hair follicle have a capacity for autofluorescence [12]. This property of stem cells has been documented by different groups [40 –42]. In addition, cells that were differentiated from stem cells were not autofluorescent [43]. To check for the presence of autofluorescence in stem cells of the earthworm, two 6-μm sections were made longitudinally, and, first, it was examined by fluorescence microscopy (data shown in Fig. 3B–D, F). Then, the sections were immunostained with anti-BrdU antibody. One of the sections was developed with BCIP/NBT (5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium) and the other with DAB. The results are presented in Fig. 3A and E, respectively. The data show almost all cells in the section fluoresced. Interestingly, BrdU-positive cells are highly autofluorescent compared with their surrounding cells. As a negative control, a thin section of rat kidney tissue was examined under the fluorescence microscope. No cells with the property of autofluorescence could be observed (data shown in Supplementary Fig. S2). The BrdU-positive cells fluoresced 2.3-fold more than the surrounding cells. To understand the statistics of the strong fluorescence of BrdU-positive cells, fluorescence cells were counted in a total of 100 BrdU-positive cells randomly selected from the regenerated blastema, circular muscle layer and longitudinal cell layer of the downstream segments of regenerated blastema. It was found that 79±6 BrdU-positive cells were more fluorescent than the surrounding cells. The counting was performed thrice with 3 different mid-part sections.
It has been reported that the major source of autofluorescence in the earthworm E. fetida is riboflavin [44,45]. To find the source of autofluorescence in the earthworm E. eugeniae, the tissues of the worm were homogenized and centrifuged, and the supernatant was subjected to thin-layer chromatography. The TLC data clearly showed yellowish-green spots that were visible to the naked eye. Under UV light, the spots fluoresced brightly with a yellowish-green color. The fluorescent compound in the worm and riboflavin had the following common characteristics: (1) the color of the compound, (2) fluorescence under UV, and (3) Same R f values in the TLC. To verify the data further, riboflavin (10 μg), 10 μL of the earthworm extracts, and a mixture of the supernatant of earthworm extract and riboflavin (5 μg) were resolved by TLC. Figure 4A shows only one spot from the mixture with an R f value identical to that of riboflavin, confirming that riboflavin mixed with the supernatant has the same R f value as the other samples chromatographed. From the data, it was concluded that the major source of autofluorescence of the worm tissues is due to riboflavin.
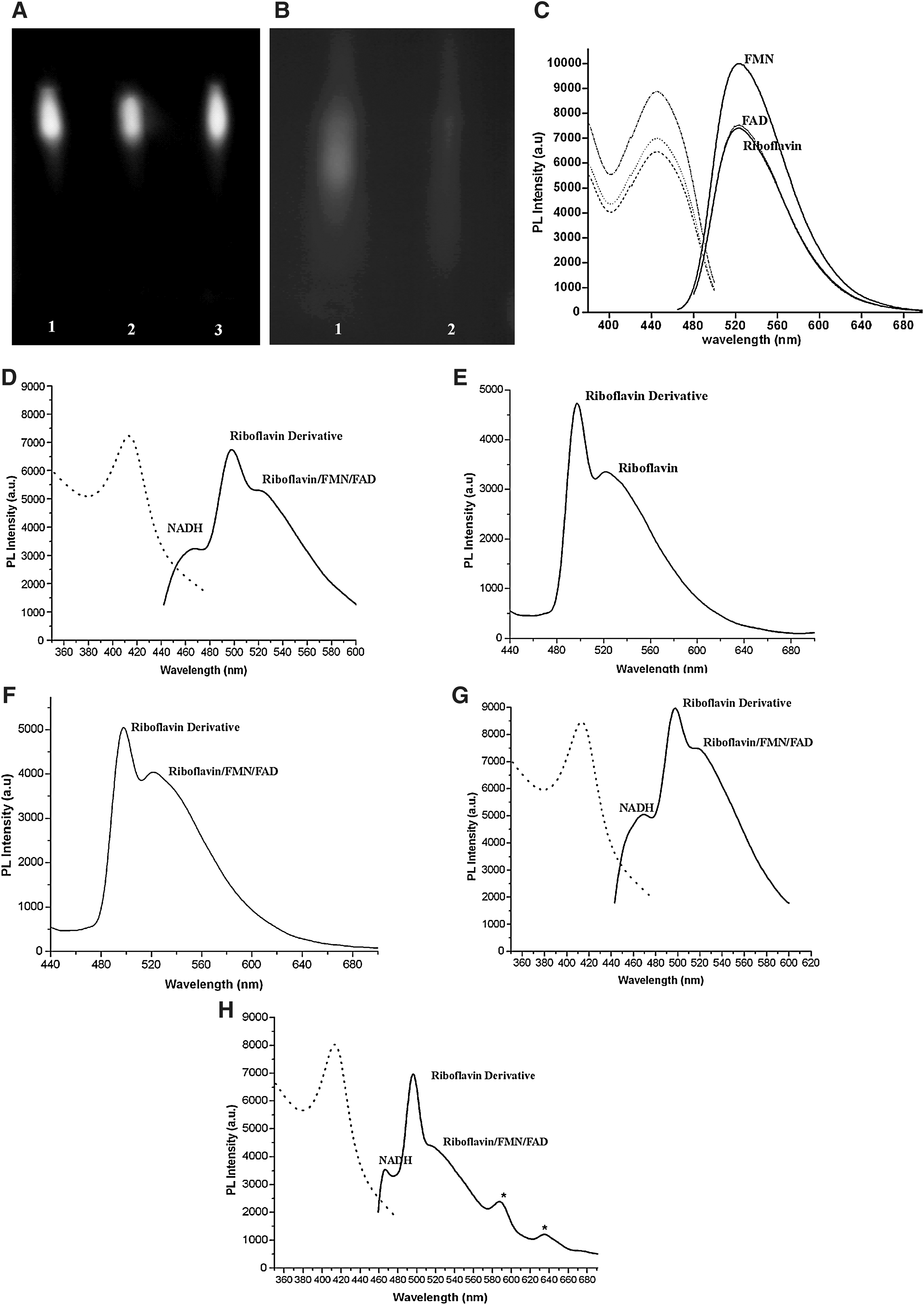
Thin layer chromatography and spectrofluorometry analysis of riboflavin, FMN, FAD, and earthworm extracts.
Spectrofluorometry was performed to further confirm the identity of the fluorescent material and also explore the existence of any other fluorescent material in earthworm tissue. The excitation and emission spectra of riboflavin, FAD, FMN, and nicotinamide adenine dinucleotide hydride (NADH) was obtained for comparison. These spectra are shown in Fig. 4C. The excitation and emission maxima of riboflavin, FAD, FMN, and NADH are 450 & 524, 450 & 522, 445 & 523, and 340 & 466 nm, respectively. The data obtained were compared with the excitation and emission maxima of the above-mentioned vitamins and their cofactors, as shown in the Supplementary Table S2. This confirmed that the experimental data are within the range of the reported excitation and emission maxima of the above vitamins and their cofactors. Further, the lysate of the circular and longitudinal muscle layers of the worm was subjected to spectrofluorometric analysis, and data shown in Fig. 4D show that in addition to the 520–525 nm peaks, which are the emission maxima of riboflavin, FAD [51], and FMN [52], there is a peak at 497 nm. To investigate the source of the emission maximum at 497 nm, the lysate of the circular and longitudinal muscle layers (80 μg of protein sample) and pure riboflavin was subjected to thin layer chromatography. Under UV illumination, both riboflavin and the fluorophore of the worm lysate showed identical R f values (Supplementary Fig. S3A). The spots corresponding to the fluorophore were scraped from the plate, and the substance (fluorophore) eluted as described in Materials and Methods. The eluates (from lysate and pure riboflavin) obtained were again resolved by TLC, and the fluorophore eluted from the scraped material as above. The eluates from the silica gel were then subjected to spectrofluorometric analysis (Fig. 4E, F for pure riboflavin and lysate, respectively).
Comparison of the spectrofluorometric data of the eluted fluorophore from the lysate (Fig. 4F) with that from direct spectrophotometric analysis of the lysate (Fig. 4D) showed no great difference in the peaks at 497 and 522 nm. The data (Fig. 4E, F) clearly illustrate that both the eluted fluorophores from TLC materials (the lysate and the riboflavin) have an emission maximum at 497 nm. The experiment was repeated carefully 4 times and consistent data were obtained. The results confirm that during TLC, riboflavin undergoes modification. Hence, the emission maximum of eluted riboflavin shifted from 522 to 497 nm. Similarly, it has been reported by Chapman and Reid [48] that riboflavin can exist in various forms in the cell as follows: mostly, it is in the oxidized form in the cell. It is converted to the semiquinone by addition of a proton and an electron. The addition of one more electron to the semiquinone produces hydroquinone. Then, C4a-peroxyflavin was produced by oxidation of the hydroquinone by addition of 2 oxygen molecules to the carbon at C4. Finally, C4a-hydroperoxyflavin results by the addition of a proton to C4a-peroxyflavin. The report suggests that riboflavin exists in different forms in biological systems. In addition, as already known, carrier proteins bind with riboflavin for transport, and FMN & FAD as coenzymes bind with different proteins of the Krebs cycle [53], amino acid synthesis [53], and electron transport chain [53]. Hence, the reason for the 497 nm emission maximum of fluorescence of worm lysate could be protein interaction with riboflavin, FAD, or FMN. To confirm this, the presence of protein in the purified fluorophore of worm lysate was tested by the Bradford's method. The data showed the absence of protein in the eluted fraction of the fluorophore (data not shown). To confirm the data further, TLC technique was used as follows: The protein sample (80 μg) of circular muscle layer and longitudinal cell layer was loaded onto the TLC plate, and the protein was detected in purple at both the lysate loading spots. The path of the fluorophore on the TLC plate showed a purple color, and a trace of the color was also seen in at the fluorophore Supplementary Fig. S3A and B. The eluted fluorophore from 80 μg of protein lysate of circular and longitudinal tissue were resolved by TLC again and visualized under UV light (Supplementary Fig. S3C). The plate was then stained with ninhydrin (Supplementary Fig. S3D). The data show the absence of protein (Supplementary Fig. S3D) in the loading spot, and also in the running path of the fluorophore. The data suggest that the protein present in the eluate was effectively removed by the TLC separation and elution protocols. The absence of proteins in the eluted fluorophore could be due to the complete denaturation and precipitation of proteins by the mobile phase of TLC [butanol (7):chloroform (4):acetic acid (5):ammonia (1):water (1)], and the further centrifugation at 10,000 rpm for 10 min removed the trace amount of the protein contaminants from the fluorophore.
Taken together, the data shown in Supplementary Fig. S3 and the Bradford protein assay reveal that the fluorophore of worm lysate eluted from the TLC plate has no protein. Hence, it is clear that the peak for the 497 nm emission maximum is not due to the protein complex of riboflavin, FAD, or FMN. Thus, it is concluded that the fluorophore (from worm lysate) responsible for the 497 nm emission maximum is nothing, but a derivate of riboflavin. Further studies are necessary to understand the nature of the riboflavin derivates present in the tissues of the worm. The data also suggest that the tissues of the circular and longitudinal muscle have significant amounts of NADH (emission maximum at 466 nm).
To extend the work further, lysate was prepared from the circular and longitudinal muscles of the regenerating worm, and the regenerative blastema, on the 4th day. They were subjected to spectrofluorometric analysis, and the data for these are shown in Fig. 4G and H, which clearly show that both the samples predominantly contain derivates of riboflavin (497 nm emission maxima), along with lesser amounts of riboflavin, FAD, and FMN. In addition to the above, a small amount of NADH is also present in both the samples. There was no significant difference between the normal and regenerating worm with respect to the fluorophores from circular and longitudinal cells. Interestingly, in the regenerative blastema, there were 2 unknown fluorophores: one with an emission maximum at 587 nm and another at 635 nm (Fig. 4H). Further studies are needed to characterize these unknown fluorophores.
To extend the study further, the coelomocytes of E. eugeniae were observed under the fluorescence microscope. It was found that coelomocytes have the capacity to fluoresce (Fig. 5A–D). The fluorescent property of the coelomocytes had been reported in many species of the earthworm. Cholewa et al. reported that only 11%–35% of coelomocytes in Allolobophora chlorotica, Dendrodrilus rubidus, E. fetida, and Octolasion sp. have the ability to fluoresce [3]. In contrast, it was found that all coelomic cells in E. eugeniae have the fluorescent property (Fig. 5A).

Autofluorescence of the different types of cells in the earthworm.
The data given in Fig. 5E, G, and H show that all cells do not fluoresce with the same intensity. Cells at the boundary of the epidermis and circular muscle layer fluoresce more strongly. In addition, a few cells in the longitudinal cell layer also exhibit a higher degree of fluorescence. The cells fluorescing at higher intensity were counted in the section of the 11th and 12th segments. It was found that 112±8 cells per segment had a 2.3-fold higher fluorescence than the others. Careful observation revealed that the fluorescence is from the cytoplasm, not from the nucleus (Fig. 5G, H). The data fit with the report that riboflavin is localized in the cytoplasm rather than in the nucleus [54]. Figure 5F shows thin sections of the above segments briefly stained with hematoxylin. In addition, it was also observed that solvents such as formaldehyde, isopropanol, and xylene used in histochemistry of the samples do not interfere with the fluorescent property of the cells.
To determine the amount of riboflavin in the worm, riboflavin in the tissues was estimated by TLC and spectrophotometry. It was found that the worm has abundant riboflavin, at a concentration of 477.5±15 μg/g. It would be of interest to understand the biological significance of riboflavin enrichment in the earthworm tissues.
To demonstrate the link between riboflavin enrichment in the worm's tissues and its regenerative capacity, 3 sets of 6 worms each was injected with a specified quantity of riboflavin once a day for 3 consecutive days. First of the sets was injected with 1.4 μg riboflavin/g of body mass, while the second set received 2.8 μg riboflavin/g of body mass. For control, the third set of the worms were injected sterile water at the same frequency. On the 3rd day, all 3 sets of worms were amputated at the 10th segment, thus blocking the intake of food as the mouth was removed. The kinetics of regeneration in the riboflavin-injected worms was compared with the control set of worms. The data (Fig. 6D–F) show that an increase in the dosage of riboflavin accelerates the regeneration process. On the 3rd day after amputation, 1±1 mm blastemas were observed with the lower-dose injection of riboflavin, while the blastema size increased to 2±1 mm in worms given the higher-dose injection of riboflavin; in control worms, the blastema size was 1±1 mm, similar to that of worms given the lower dose of riboflavin. Similarly, on the 5th day, blastemal size was 2±1, 4±1, and 2±1 mm in the lower-dose injected, higher-dose injected, and control worms, respectively. Likewise, on the 7th day, 4±1, 8±1, and 4±1 mm blastemal size was observed, respectively. These data clearly show that riboflavin promotes the regeneration process.

Regeneration kinetics upon injection of antibiotics, riboflavin (vitamin B2), and antibiotics+riboflavin on earthworm.
It is known that the riboflavin can be synthesized by microbes [55 –57] and plants [58 –60]. To understand the source of riboflavin production, a set of 6 worms was injected once a day for 3 consecutive days with a mixture of antibiotics at the following concentrations: penicillin, 10 U; amphotericin B, 0.025 μg; streptomycin, 0.010 mg. The control set of worms received an injection of 10 μL of sterile water at the same frequency. Penicillin and streptomycin have a broad-spectrum action against both gram-positive and gram-negative organisms, while amphotericin B is an anti-fungal agent. It was found that riboflavin production was reduced into 74±5 μg/g body mass after injection of antibiotics (Fig. 4B). The injection of antibiotics reduced the concentration of riboflavin to 1/7 the concentration in the control worms. The data confirm that microbes are the source of riboflavin in the earthworm. To understand the role of riboflavin in the regeneration process, it is necessary to reduce the riboflavin content in the worm. For this, 2 sets of 6 worms each were injected with a combination of penicillin, amphotericin B, and streptomycin. Two doses of the antibiotics (penicillin, 5 U; amphotericin B, 0.0125 μg; streptomycin, 0.005 mg) and (penicillin, 10 U; amphotericin B, 0.025 μg; streptomycin, 0.010 mg) were injected once a day for 3 consecutive days into separate sets of worms. The control set of worms received an injection of 10 μL of sterile water at the same frequency. On the 3rd day, all the 3 sets of worms were amputated at the junction of the 10th and 11th segments. The results of this experiment are shown in Fig. 6G–I. On the 3rd day after amputation, 1±1 mm blastemal size was observed at the lower-dose injection of antibiotics, while no blastema was observed in worms given the higher-dose injection of antibiotics. The control worms had a blastema size of 1±1 mm on the 3rd day. On the 5th day, 2±1, 1±1, and 2±1 mm blastemal size was observed in the lower-dose, higher-dose antibiotics, and sterile water-injected worms, respectively. Likewise, on the 7th day, 3±1, 2±1, and 4±1 mm blastemal size was observed in the lower-dose, higher-dose, and zero-dose antibiotics-injected worms, respectively. Taken together, the results showed that antibiotic injection reduces the concentration of riboflavin in the worm, and that antibiotic administration hampers the kinetics of blastema formation. The reduction of blastemal growth may also be due to the side effects of the antibiotics.
To confirm the results obtained on suppression of blastemal growth by antibiotics, a rescue assay was performed. A group of 6 worms was first injected with antibiotics (penicillin, 10 U; amphotericin B, 0.025 μg; streptomycin, 0.010 mg), followed by an injection with riboflavin (2.8 μg riboflavin/g body mass) after 12 h. The control set of worms received 10 μL of sterile water injection at the same frequency. This injection pattern was repeated for the next 2 days. On the 3rd day, all the sets of worms were amputated from segments 1–5, thus blocking the intake of food as the mouth was removed. Regeneration was observed on the 3rd day after amputation. The regeneration rates of the control worm and those injected with antibiotics and riboflavin are shown in Fig. 6A–C and J–L. The results clearly show that upon injection of antibiotics followed by riboflavin, the regenerative capacity of the worms, hampered by the injection of antibiotics, was revived by the subsequent injection of riboflavin. On the 3rd, 5th, and 7th day after amputation, 2±1 & 1±1 mm, 4±1 & 2±1 mm, and 6±1 & 4±1 mm blastemas were observed in worms injected with antibiotics followed by riboflavin, and control worms, respectively. Taken together, riboflavin is observed to reverse the inhibitory effect of antibiotics on blastemal growth. For example (Fig. 6M), the riboflavin-injected worm generated 2±1 mm of blastema on the 3rd day, while injection of antibiotics alone did not result in blastemal growth till the 3rd day. Similarly, the rescue effect of riboflavin was observed on the 5th day. In contrast, the inhibition of blastemal growth was not completely rescued even on the 7th day in worms injected with antibiotics alone. The difference in the size of the blastemas on the 7th day between riboflavin-injected worms and those injected with antibiotics followed by riboflavin is 1 mm. This difference might be due to the adverse effect of antibiotics. It is known that antibiotic injection causes such adverse effects [61]. Hence, the rescue assay data clearly confirm that riboflavin is not only the major source of auto-fluorescence but also the key factor for regeneration in the earthworm, E. eugeniae.
Discussion
Regeneration is the process of restoration of lost body parts. It was reported that during the regeneration process, a regeneration blastema was formed with a proliferative mass of undifferentiated progenitor cells, from which newly differentiated cells arise [23,24]. The wound of an amputated site was healed quickly at 24 h in the earthworm, E. eugeniae. The regeneration blastema formed at 48 h after amputation is soft and transparent. During regeneration, the reduction of cell density in the circular muscle layer and the increase of the longitudinal cell layer suggest that the circular muscle cells may be differentiated into longitudinal cells or may undergo apoptosis. The amputation at segments 10–11 removed not only vital organs such as the heart and the simple brain, but also the mouth. The removal of the mouth blocks the intake of nutrients. In this critical situation, apoptosis leading to loss of circular muscle cells may not occur. Hence, the removal of the circular muscle layer by apoptosis may cause additional energy loss, and differentiation of the circular muscle cells into the longitudinal cell layer could save the energy for the rest of the regeneration process. Hence, there is a greater possibility for differentiation of the circular muscle layer into the longitudinal cell layer.
The longitudinal cells further divide and form the blastema. The complete regeneration of the anterior part of the worm took 24 days. Hence, the experimental data suggest that cells of the blastema, which is formed by longitudinal cells, have the ability to differentiate into many different types of cells to regenerate the organs present in segments 1–10. The histological data of the 7th day regeneration blastema clearly show the formation of septa, but organs were not yet developed in the segments. The data suggest that the internal organs are developed after the formation of segments.
The presence of BrdU in the blastemal tissue (Fig. 2A), which is produced newly, confirms the migration of BrdU-positive cells from the adjacent segments to the blastema. Besides, the predominant niche of BrdU-positive cells in the 11th and 12th segments (upstream of the blastema) in the junction of the circular muscle layer and epidermis (Fig. 2F) further supports the stem cells to migrate from the 2 layers of skin to the blastema. The presence of more BrdU-positive cells in the 11th and 12th segments during regeneration (Fig. 2F, H) than in the control worm (Fig. 2G, H) might be due to either the migration of stem cells from the downstream segments to amputation site or renewal of stem cell.
In accordance with the previous reports [44,45], the data of TLC and spectrofluorometry (Fig. 4) show that the major source of fluorescence is riboflavin, and its derivates. It has been reported that riboflavin is an electron carrier in the Krebs cycle [53], electron transport chain [53], amino acid synthesis [53], lipid biosynthesis [53], and DNA repair [62]. In this study, it is proved that riboflavin is essential for regeneration too (Fig. 6J–L). In general, gut microbes produce ethanol, riboflavin, other vitamins, and secondary metabolites. In the scenario, Rawls et al. [63] reported that in a microbe-free (sterile) environment, the development of zebrafish was affected, leading to the formation of developmentally abnormal young ones. However, they did not find the substances whose absence was responsible for the developmental defects. The abnormal development of zebrafish in a sterile environment may be due to the riboflavin deficiency as the regeneration of organs from the blastema is comparable with the normal process of embryogenesis.
The data on strong fluorescence from BrdU-positive cells (Fig. 2) confirm that stem cells have the capacity to accumulate more riboflavin than the surrounding cells. The fluorescence of stem cells has been reported in rat and mice [40 –42]. Interestingly, Wun et al. reported autofluorescence of human skin stem cell. Hence, it is concluded that the fluorescent property of stem cells is conserved from worms to humans. The studies on riboflavin accumulation in the cell can shed light on stem cell and regeneration biology. The earthworm, E. eugeniae, is an ideal model to study the mechanism of riboflavin accumulation in stem cells, and the role of riboflavin in stem cell biology and regeneration.
Footnotes
Acknowledgments
This work was financially supported by Department of Biotechnology, New Delhi. We sincerely thank Dr. H. Nellaiah, Department of Biotechnology, Kalasalingam University, for critical reading of the article.
Author Disclosure Statement
The authors declare no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.