
Abstract
To investigate age-related intrinsic regulation of the capacity of human fetal oligodendrocyte progenitor cells (OPCs) to myelinate, potential OPCs were selected from 15- to 23-gestational-week (gw) human fetal brain tissue based on the expression of gangliosides—recognized with the monoclonal antibody A2B5, which detects multipotent cells including OPCs—or platelet-derived growth factor receptor α (PDGFRα), an early marker of the oligodendroglial lineage. Cells were either cultured alone or cocultured with rat dorsal root ganglia neurons (DRGNs). When cultured alone, both the A2B5- and PDGFRα-selected cells exhibited age-dependent increases in early to mid-stage lineage markers, including sulfatides (O4 antibody) and the transcription factor Olig2, while the cell death rate correlated negatively with age. In coculture with neurons, cells also expressed the myelin components galactocerebroside (GC) and myelin basic protein (MBP), and ensheathed axons. In DRGN cocultures, A2B5+ cells derived from >19 gw produced more GC+/MBP+ cells compared with the 15–17-week cells. The number of GC+ cells making axonal contacts, and ensheathing axonal segments per cell increased proportionally to gestational age. This age-dependent difference in GC/MBP cell number and capacity to ensheath axons persisted when PDGFRα selection was used to enrich for the number of OPCs in cultures derived from younger ages. Addition of the growth factors brain-derived neurotrophic factor (BDNF) and insulin-like growth factor 1 (IGF-1) enhanced OPC differentiation under all conditions. These findings indicate that intrinsic regulatory mechanisms associated with the chronological age of the donor cells are key variables to assess when considering the myelination capacity of OPCs for cellular replacement therapy.
Introduction
M
The availability of human fetal tissues for in vitro studies of progenitor cells is largely restricted to first and early second trimester samples, thus preceding the time when myelination has begun or when significant numbers of oligodendroglial lineage cells can be detected in situ. Most studies of human progenitors have involved selection of cells based on expression of gangliosides recognized with the monoclonal antibody A2B5. This A2B5+ population from second trimester human fetal central nervous system contains only a small percentage of cells that have entered the oligodendrocyte progenitor lineage as determined with Olig2, PDGFRα, and O4 antibodies [9], consistent with the results of in situ studies [2]. The proportion of such cells increases in dissociated cell culture in the presence of growth factors [PDGF/fibroblast growth factor (FGF)] but remains a minor population <10% for up to 10 days [9]. Few maturing oligodendrocytes (GC and MBP) are detected in such cultures. As we demonstrated previously, in coculture with rat dorsal root ganglia neurons (DRGNs), a number of A2B5-selected cells differentiated into GC/MBP+ oligodendrocytes, which was accompanied by axonal ensheathment, an initial step in the myelination process [9].
In the current study, we analyze the properties of progenitor cells selected with A2B5 or PDGFRα antibodies from mid-gestational (15–23 weeks) fetal human brain in isolated cultures and in coculture with DRGNs. This combined approach was used to address the contribution of the chronological age on both early and late stages of oligodendrocyte progenitor cell (OPC) differentiation and their interactions with axons.
Materials and Methods
Human fetal OPC cultures
Human fetal brain tissue obtained from 15- to 23-gw embryos was provided by the Human Fetal Tissue Repository (Albert Einstein College of Medicine, Bronx, NY). Both the Albert Einstein College of Medicine and the McGill University institutional review boards approved these studies. A2B5+ cells were isolated immunomagnetically as previously described [9,10]. Similarly, PDGFRα+ cells were selected with a monoclonal antibody mouse anti-human CD140α (AbD Serotec, Kidlington, Oxford) followed by rat anti-mouse IgG1 magnetic beads (Miltenyi Biotech, Auburn, CA).
Isolated cell cultures
Cells were plated on a matrix of lysed human fetal astrocytes (astrocyte matrix) grown on poly-L-lysine–coated plastic coverslips (Nunc, Rochester, NY) as described previously [9,10]. The cultures were grown in Dulbecco's modified essential medium (DMEM)-F12 supplemented with N1 (Sigma, Oakville, ON); 0.01% bovine serum albumin (BSA); 1% penicillin-streptomycin, B27 supplement (Invitrogen, Burlington, ON); thyroid hormone (T3, 2 ng/mL; Sigma); PDGF-AA (10 ng/mL; Sigma); and basic FGF2 (20 ng/mL, Sigma).
DRGN: human OPC cocultures
Purified DRGN cultures were prepared from Sprague-Dawley rat embryos as described previously [11]. The cultures were maintained with 12.5 ng/mL nerve growth factor (NGF; Alomone Labs, Jerusalem, Israel) in serum-free N1-supplemented medium. Human fetal A2B5+ or PDGFRα+ cells freshly isolated from the total brain fraction were added at a density of 0.7×105 cells/cm2 to DRGN cultures in their third culture week. DRGNs are maintained for ∼3 weeks to allow establishment of a differentiated neuritic network and make axons competent for myelination.
Immunocytochemistry
For assessment of cell surface markers, cells were stained with monoclonal antibodies A2B5 (IgM, 1:50 [12]), O4 (IgM, 1:50 [13]), RmAb (IgG3, 1:50, against both sulfatide and GC [14,15]), and O1 (IgM, 1:25, against GC [13]) for 30 min at 4°C, fixed with 4% paraformaldehyde for 10 min at 4°C, and blocked with HHG (1 mM HEPES, 2% horse serum, 10% goat serum, and Hanks' balanced salt solution) for 10 min. Cultures were incubated with appropriate secondary antibodies conjugated with fluorescent dyes goat anti-mouse IgM-FITC or -Cy3 and IgG3-FITC (1:100; Jackson ImmunoResearch, Westgrove, PA) for 30 min at 4°C. Antibody isotype controls showed low nonspecific staining (data not shown). Olig2 (1:200; IBL, Gunma, Japan), neurofilament (NF, 1:1000; Sigma), or MBP (1:500; Sternberger, Lutherville, MD) antibodies were applied after the cells were fixed and permeabilized with 0.3% Triton X-100 followed by secondary antibodies goat anti-rabbit Cy3 (1:100; Jackson ImmunoResearch), goat anti-mouse IgG1-FITC (1:100; Molecular Probes, Eugene, OR), and goat anti-mouse IgG2b-TxR (1:100; Biosource, Camarillo, CA) and counterstained with Hoechst 33342. Terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling (TUNEL)-positive cells were revealed by staining with a commercial kit (Promega). Cell nuclei were stained with Hoechst 33258 (1:1000; Invitrogen) for 10 min at room temperature. For the studies of purified OPCs matured in culture, slides were mounted using gel/mount (Biomeda Corporation, Foster City, CA), and stains were visualized using epifluorescent microscopy (Leica, Montreal, QC) and OpenLab imaging software (OpenLab, Florence, Italy). For coculture studies, slides were visualized by confocal microscopy using a Zeiss LSM 510 (Zeiss, Oberkochen, Germany) with a 63× oil objective and analyzed using LSM 510 Image browser software (Zeiss). The optical thickness was less than 1.0 μm for each channel.
Data analysis
For studies of oligodendrocyte culture, the proportion of cells expressing specific lineage markers was determined by counting 400–600 cells in each of three high-power fields (25×) per coverslip. Data are presented as the mean values of three experiments performed. For coculture studies, the number of individual experiments performed and the number of cells counted for each investigation are provided in the “Results” section and/or corresponding figure legends. Results are presented either as the mean±SEM or as the fold or percent difference over control at the corresponding time point. Statistical significance was determined by one-way or two-way analysis of variance, followed by either Dunnett's or Bonferroni's multiple comparison tests. Probability values < 0.05 were considered statistically significant.
Results
Isolated cell cultures
Studies on A2B5-selected cells
To compare the temporal expression of oligodendrocyte-specific markers on brain-derived progenitors selected from different

Age-related differentiation of A2B5-selected cells in isolated culture. Cells selected from fetal brain samples at different gestational weeks (gw) were cultured for 6 days before immunostaining with Olig2, O4, and GC antibodies. Cell death was measured with TUNEL. An age-related correlation in number of cells expressing oligodendrocyte lineage markers
After culturing A2B5-selected human fetal brain cells for 6 days in vitro in growth factors (GF's) supplemented conditions, the number of O4-positive cells dying by apoptosis was examined using TUNEL, which assesses nuclear condensation and fragmentation. The rate of cell death ranging from 22%±1.2% to 8.7%±0.9% was inversely correlated with gestational age (15–20 gw) (Fig. 1C). Over all age ranges, omission of growth factors increased cell death rates to 29%±3.2% and 15%±1.4% for 15–17 and 19–20 gw, respectively (P<0.01).
PDGFRα-selected cells
Since A2B5 also recognizes cells outside the OPC lineage in the fetal brain [16], we selected cells using an antibody against PDGFRα, a relatively early marker of the oligodendrocyte lineage [17] and cultured these cells under the same conditions as applied to the A2B5-selected cells. The total cell number recovered using PDGFRα selection was 0.22%±0.05% at the youngest ages (15–17 gw, n=6), and increased to 0.94%±0.12% in older age samples (19–20 gw, n=3, P<0.05), corresponding to 0.6%–5% of that recovered using A2B5 antibody selection. After overnight in culture (a time needed to allow cell attachment to substrate), the PDGFRα-selected fraction contained 52.9%±2.6% Olig2+ and 51.1%±3.2% O4+ cells, and was higher than the A2B5-selected fraction (10.5%±0.5% for Olig2, 8.7%±0.2% for O4, P<0.05). After 6 days in culture, the PDGFRα-selected fraction contained significantly higher numbers of O4- and Olig2-positive cells as compared with the A2B5-positive fraction (Fig. 2A–F). In addition, the proportion of O4- (47%±1.2% vs. 27%±1.2%) and Olig2-positive (48%±1.7% vs. 32%±1.2%) cells was significantly increased (P<0.05) in the PDGFRα-positive fractions from the older samples (Fig. 2E, F). Similar to the A2B5-positive cells, a significant inverse correlation was detected between gestational age and the number of O4-positive cells dying by apoptosis (24.7%±1.6% and 12.5%±1.7% for 15–16 gw and 18–20 gw, respectively, P<0.05). The ability of the PDGFRα-selected cell population to differentiate into GC+ oligodendrocytes was higher in the presence of brain-derived neurotrophic factor/insulin-like growth factor (BDNF/IGF-1) (19%±2.5%) than with PDGF-AA/FGF2 (10.3%±1.3%, P<0.05), as previously reported for A2B5-selected cells.
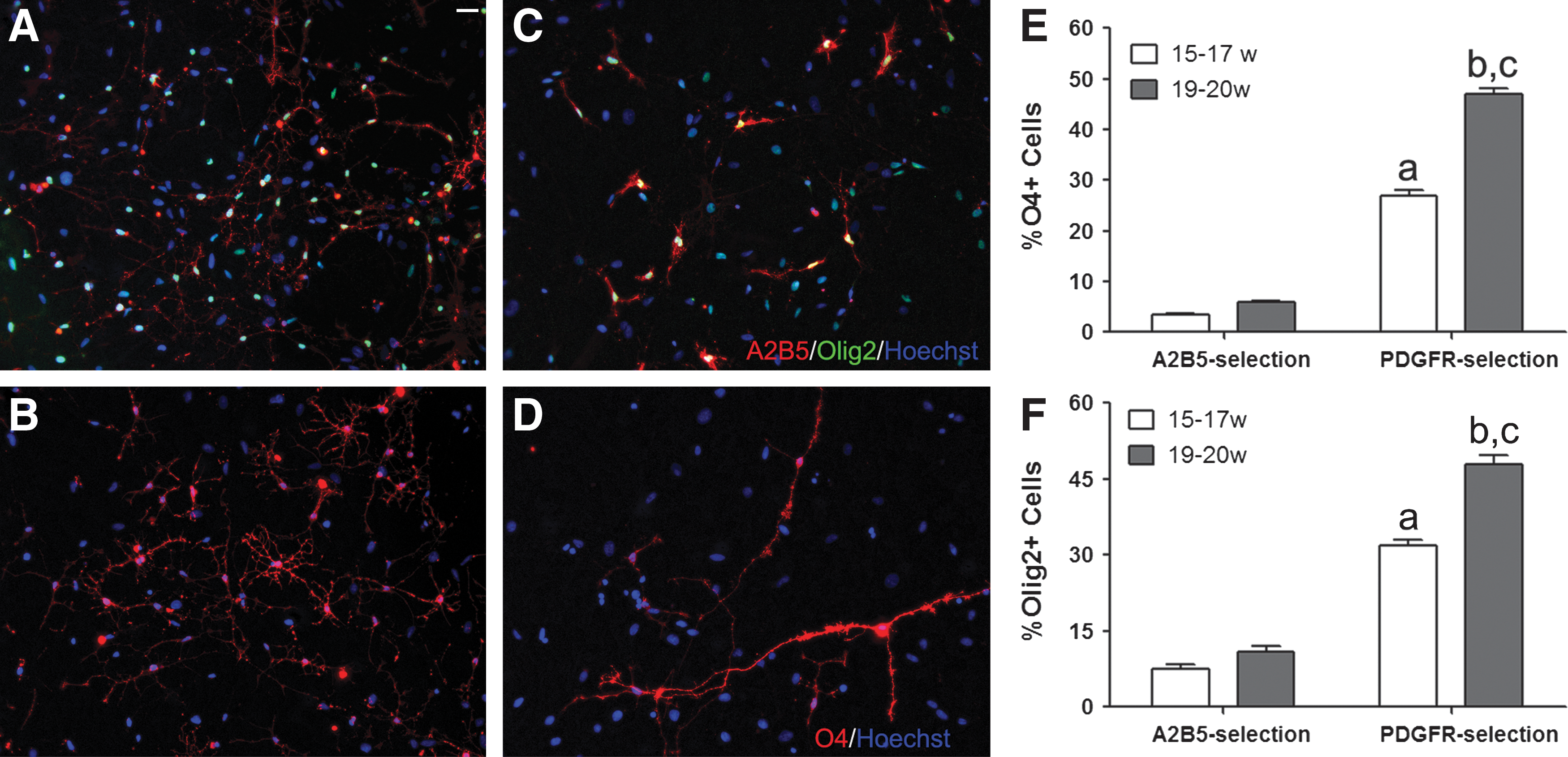
Comparison of age-related phenotypic properties of A2B5- and platelet-derived growth factor receptor α–selected cells in isolated culture. Cells from 17-gw brain were selected with A2B5
Coculture studies
A2B5-selected cells were cocultured with rat DRGNs for 4 weeks under defined conditions (NGF+/− BDNF/IGF-1) and examined for expression of GC (O1 antibody). In the presence of GFs, the number of GC-positive cells derived from the 19–20 gw A2B5-selected fraction (258±46.6 per coverslip) was significantly increased compared with those derived from the 15–17 gw samples (26±2.3 per coverslip, P<0.001, n=3) (Table 1). Although the number of MBP-expressing cells was lower than GC+ cells for both age groups, they were higher in the older group (84±8.7 vs. 5±0.7, P<0.001, n=3). Similarly, the percentage of GC+ cells making axonal contacts was significantly greater for the older A2B5-selected cells compared with the younger ones (34.7%±4.3% for 15–17 gw vs. 17.9%±1.3% for 19–20 gw, P<0.01, n=3) (Table 1). In addition, the number of GC+ cells making contacts with multiple (>10) axons was significantly higher (90%±1.7%) for the older (19–20 gw) than for the younger age samples (13.5%±7.8%) (as illustrated in Fig. 3A, B and quantitated in Fig. 3D). Supplementing the cultures with BDNF and IGF-1 resulted in a greater number of OPCs differentiated into GC+ and MBP+ oligodendrocytes and making axonal contacts for all groups (Table 1). Examination of contactin-associated protein and sodium channel proteins showed clustering of these two markers in the contacts between axons and GC+ oligodendrocytes, but the formation of nodal and paranodal axonal domains remained incomplete, suggesting that myelination is at the early stages (Fig. 3C).
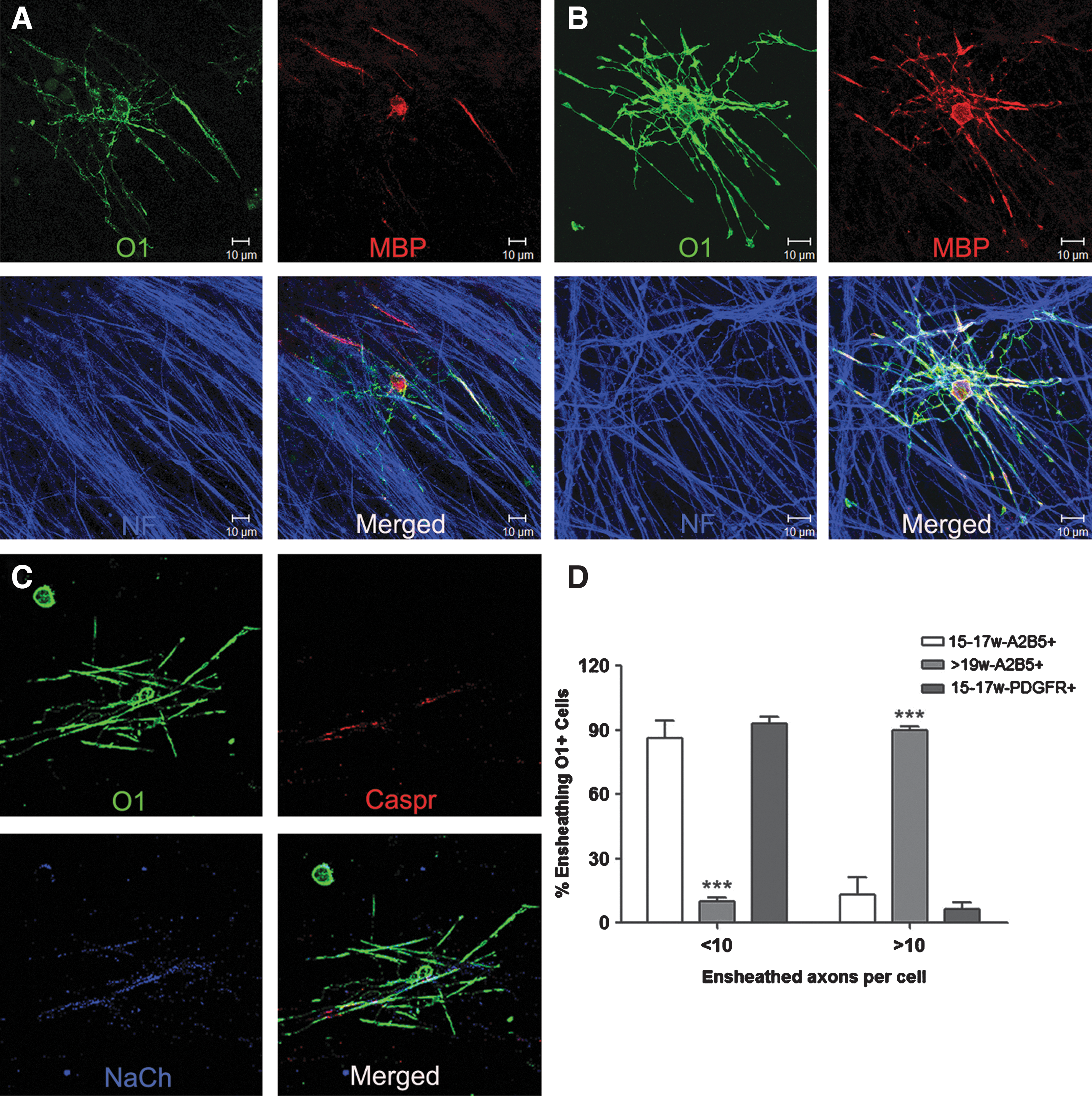
Differential capacity of oligodendrocyte progenitor cells to ensheath rat dorsal root ganglia neuron axons in culture. Cells selected with A2B5 from 16-gw
Cells of 15–17 and 19–20 gw were selected with A2B5 or PDGFRα antibodies and cocultured with DRGNs in the presence of NGF+/− GFs (BDNF+IGF-1) for 4 weeks. Cultures were fixed and immunostained for neurofilaments (NFs, far-red), MBP (red), and O1 (green) as illustrated in Fig. 3. The number of O1+ and MBP+ cells as well as percentage of O1+ cells making contact with DRGN axons was quantified. Data represent three independent experiments performed in duplicate for each condition (∼200 cells were counted per coverslip). Statistical significance was determined by two-way ANOVA, followed by Bonferroni's test. All values for conditions containing GFs are significantly greater than for conditions lacking GFs (P<0.001). For each corresponding category, P<at least 0.05.
Compared with 15–17 gw-A2B5+NGF.
Compared with 15–17 gw-PDGFR+NGF.
Compared with 15–17 gw-A2B5+NGF+GFs.
Compared with 15–17 gw-PDGFR+NGF+GFs.
CVS, coverslips; MBP, myelin basic protein; NGF, nerve growth factor; GFs, growth factors; PDGFRα, platelet-derived growth factor receptor α; DRGNs, dorsal root ganglia neurons; BDNF, brain-derived neurotrophic factor1; IGF-1, insulin-like growth factor-1; ANOVA, analysis of variance.
To evaluate whether the lower values found with the younger age samples was due to the small proportion of A2B5-positive cells committed to the oligodendrocyte lineage [9], we assessed the differentiation potential of the PDGFRα-positive fractions derived from younger age samples in the coculture system. As shown previously in short-term cultures of purified oligodendrocytes, the PDGFRα-positive fraction produced a higher proportion of oligodendrocytes (sulfatides and Olig2) compared with the A2B5-positive fraction derived from samples of either age (Fig. 2). In cocultures, the number of cells in the PDGFRα-positive fraction (15–17 gw) expressing GC (119±10 per coverslip) or MBP (36±5.8 per coverslip) (Table 1) was significantly higher than for the A2B5 fractions derived from samples of the same gestational age (26±2.3 and 5±0.7 per coverslip for GC and MBP, respectively), but still significantly lower than from older samples (19–20 gw) (258±46.6 and 84±8.7 per coverslip for GC and MBP, respectively). Although the proportion of O1-positive cells from the PDGFRα fraction contacting axons (30%±2.7%) was comparable with that of GC-positive cells from the older age A2B5 cell fraction (34.7%±4.3%) (Table 1), the extent of axonal ensheathment by the PDGFRα fraction was significantly lower (>10 axons, 6.75%±3.1%; Fig. 3D).
Discussion
Our findings demonstrate that intrinsic mechanisms exert a substantial influence on OPC differentiation and the initiation of myelination. In short-term isolated cultures of either A2B5- or PDGFRα-selected cells, which are well suited for the analysis of oligodendrocytes at early stages of differentiation, we show an age-dependent increase in the proportion of cells expressing the earlier lineage markers sulfatides and Olig2. The proportion of cells expressing sulfatides and Olig2 was significantly greater for the PDGFRα selection as compared with A2B5 selection, supporting the conclusion that PDGFRα is a more specific marker for OPCs during human fetal brain development. The relatively low number of OPCs found in the earlier brain developmental stage may be due to lack of environmentally derived survival signals as has been suggested by rodent studies; these include direct contact with axons [7,18] or soluble (trophic) factors secreted by astrocytes and neurons [19]. The mid-second trimester cells used in our study are derived at a time prior to axonal maturation [20]. We show that even in the presence of growth factors, there is significantly higher cell death in the younger samples whether the cells were selected with A2B5 or PDGFRα antibodies, raising the possibility that enhanced apoptotic elimination may be a characteristic of the younger samples. This possibility is supported by a previous study that revealed higher levels of the proapoptotic genes bax and procaspase-3 expressed in oligodendrocyte progenitors versus mature oligodendrocytes [21]. Interestingly, a recent study using transgenic mice reported that knocking out bax increased the survival of oligodendrocyte lineage cells [22].
In our OPC–DRGN cocultures, which are well suited for the analysis of cells at relatively late stages of differentiation, we show that the number of cells expressing GC or MBP derived from the A2B5-positive fraction purified from >19-week brain tissues was significantly higher as compared with A2B5-selected cells derived from 15- to 17-week samples, as were the proportion of GC+ cells that made contacts with axons. These gestational age–related differences in the number of GC/MBP cells and the capacity to ensheath axons persisted even when cocultures were initiated with PDGFRα-positive fractions derived from younger brain samples, indicating that the differences could not be ascribed only to the small number of oligodendrocyte progenitor lineage cells present in the A2B5-positive fractions. Comparison of the A2B5-positive and PDGFRα-positive fractions indicates that the number of axonal contacts increases as one enriches for cells committed to the oligodendroglial lineage.
We further noted that the number of ensheathed axonal segments per cell was markedly reduced with the younger (90%<10 axons per cell) versus older cell samples (90%>10 axons per cell) even when the number of axonal contacts was relatively comparable (PDGFRα younger samples vs. A2B5 older samples). Heterogeneity in the number of myelin sheaths made per oligodendrocyte has been well documented, and this has been linked to the diameter of the axons myelinated, with an increased number of sheaths per cells for those oligodendrocytes that ensheath small axons [23]. All of our coculture studies presented here utilize the same type of neuronal cultures. Previous studies using rat OPCs in this coculture system, performed with cells obtained at the time of spontaneous ongoing myelination, show that the axons can support complete myelination [11]. We cannot exclude the possibility that progenitors derived from tissue samples of different ages may represent distinct populations, as has been suggested by the rodent literature [24].
In conclusion, our combined results obtained from isolated cell cultures and neuronal cocultures—while demonstrating the expected influence of extrinsic signals on OPC development—strongly implicate the presence of intrinsic regulatory mechanisms throughout oligodendrocyte development. Potential intrinsic regulators include stage-specific expression of activators [9,25,26] and repressors [22 –24] of transcription. Other important factors that regulate oligodendrocyte development include epigenetic signals [27,28], and signal transduction molecules [29 –32]. Modulation of these intrinsic regulatory mechanisms could be used to enhance the effectiveness of cellular replacement therapy in a range of human dysmyelinating conditions, as well as providing means to promote endogenous remyelination in multiple sclerosis.
Footnotes
Acknowledgments
This research was funded by a postdoctoral fellowship from the Multiple Sclerosis Society of Canada (MSSC) to Q. Cui and by operating grants to J. Antel and G. Almazan from the National Multiple Sclerosis Society and the MSSC.
Author Disclosure Statement
No competing financial interests exist.