
Abstract
To develop methods for the generation of insulin-producing β-cells for the treatment of diabetes, we have used GFP-tagged embryonic stem cells (ESCs) to elucidate the process of pancreas development. Using the reporter Pdx1GFP/w ESC line, we have previously described a serum-free differentiation protocol in which Pdx1-GFP+ cells formed GFP bright (GFPbr) epithelial buds that resembled those present in the developing mouse pancreas. In this study we extend these findings to demonstrate that these cells can undergo a process of branching morphogenesis, similar to that seen during pancreatic development of the mid-gestation embryo. These partially disaggregated embryoid bodies containing GFPbr buds initially form epithelial ring-like structures when cultured in Matrigel. After several days in culture, these rings undergo a process of proliferation and form a ramified network of epithelial branches. Comparative analysis of explanted dissociated pancreatic buds from E13.5 Pdx1GFP/w embryos and ESC-derived GFPbr buds reveal a similar process of proliferation and branching, with both embryonic Pdx1GFP/w branching pancreatic epithelium and ESC-derived GFPbr branching organoids expressing markers representing epithelial (EpCAM and E-Cadherin), ductal (Mucin1), exocrine (Amylase and Carboxypeptidase 1A), and endocrine cell types (Glucagon and Somatostatin). ESC-derived branching structures also expressed a suite of genes indicative of ongoing pancreatic differentiation, paralleling gene expression within similar structures derived from the E13.5 fetal pancreas. In summary, differentiating mouse ESCs can generate pancreatic material that has significant similarity to the fetal pancreatic anlagen, providing an in vitro platform for investigating the cellular and molecular mechanisms underpinning pancreatic development.
Introduction
P
In the developing mouse embryo, pancreatic commitment is foreshadowed by regionalized upregulation of Pdx1 expression within a restricted domain on the dorsal and ventral surfaces of the gut tube at embryonic day (E) 9 to 9.5 [6]. These Pdx1 + areas evaginate to form the dorsal and ventral pancreatic buds that subsequently expand in response to signals from adjacent pancreatic mesenchyme [7]. By E12.5, the developing anlagen is further elaborated to incorporate lobular outgrowths at the distal tips of the expanding pancreatic tree. This process of branching morphogenesis concomitantly continues with further cell specification and differentiation events that incorporate acinar and endocrine components into the developing organ. Over this period Pdx1 expression remains high in both the dorsal and ventral anlagen and is also detected in the posterior region of the stomach, the bile duct, and the duodenal epithelium [8]. Lineage-tracing experiments have demonstrated that Pdx1 expressing progenitors in the early embryo give rise to all 3 types of pancreatic cells from the exocrine, endocrine, and duct [9]. In the adult, Pdx1 is expressed at high levels in β-cells, where it activates insulin gene expression [6], while lower levels of Pdx1 expression can also be detected in the acinar tissue.
Embryonic stem cells (ESCs) are immortal pluripotent cell lines characterized by their ability to differentiate into specialized cell types representing derivatives of the ectodermal, endodermal, and mesodermal lineages [10,11]. The in vitro differentiation of these cells recapitulates many aspects of early embryonic development and is a valuable model for studying processes occurring during inaccessible periods of embryogenesis. As such, we and others have utilized this model to investigate the in vitro requirements for the formation of pancreatic endoderm from ESCs and their subsequent differentiation to endocrine and exocrine cell types [12 –26].
We had previously devised a serum-free differentiation protocol that fostered the generation of pancreatic bud-like structures from differentiating mouse Pdx1GFP/w ESCs and showed that GFP bright (GFPbr) buds harbored precursors of both the endocrine and exocrine lineages and could give rise to insulin+ cells [25]. In the present study, we further dissect the potential of these ESC-derived GFPbr buds by examining their ability to undergo branching morphogenesis in a Matrigel-based culture system and perform comparative analysis with pancreatic rudiments from the developing mouse embryo. Our study demonstrates that both the differentiating ESCs and the fetal pancreas pass through a transient stage during which they are competent to undergo branching morphogenesis. Indeed, comparative analysis showed parallels between the dynamics of branching in structures derived from Pdx1-GFPbr bud containing embryoid bodies (EBs) and embryonic pancreas explants. These results, coupled with gene expression analyses of branching cultures, suggest similarities in the cellular composition of pancreatic-committed progenitors present in the in vitro and in vivo derived cell populations. Overall, our studies highlight the utility of ESCs in the exploration of key events during pancreatic development.
Materials and Methods
ESC culture and differentiation
The mouse embryonic stem cell line Pdx1GFP/w
was maintained on irradiated primary mouse embryonic fibroblasts as previously described [18,25,27]. ESC differentiation was performed using a modification of the method previously described [27] and employed a “spin EB” platform originally developed for differentiation of human ESCs [28]. Briefly, individual spin EBs were formed by the forced aggregation of 350 cells/well in low attachment 96-well round-bottomed plates (Costar®) containing chemically defined medium (see Supplementary Table S1; Supplementary Data are available online at
Generation of Pdx1GFP/w targeted heterozygous embryos
Pdx1GFP/w embryos were generated by timed-matings of Pdx1GFP/w males [29] with C57/B6J females. The presence of the vaginal plug was designated as embryonic day 0.5 (E0.5) of gestation. Embryos of the desired age were dissected from the decidua and the pancreatic anlage, including both the dorsal and ventral pancreatic buds (including some surrounding mesenchyme), caudal stomach, and proximal duodenum was isolated (herein denoted as pancreatic/Pdx1+ rudiments). Images of embryos expressing GFP were captured with a Leica fluorescence microscope. All animal experimentation was performed under the auspices and approval of the Monash University School of Biomedical Sciences animal ethics committee (approval number SOBSA/MIS/2010/21).
Explant cultures
Pancreatic explants were cultured using a Matrigel based culture system. Briefly, 100 μL of a solution comprised of 50 μL CDM (see Supplementary Table S1) and 50 μL Growth Factor Reduced (GFR) BD Matrigel™ Matrix (Becton Dickinson) was added to each well of a 4-well Chamber Slide™ System (Nunc Lab-Tek®). The GFR-Matrigel was allowed to gel at 37°C for ∼30 min. For the partial disaggregation of pancreatic rudiments or EBs, samples were collected and washed once in PBS. Samples were resuspended in 1 mL Trypsin/Versine containing 2% chicken serum and were incubated at 37°C for 4–7 min. Trypsin digestion was arrested with the addition of equivalent volume of fetal calf serum and the samples were gently pipetted to facilitate partial disaggregation. Following 2 rounds of washing with PBS, cells were resuspended in 500 μL of CDM per 5 fetal rudiments or per 12 disaggregated EBs. This cell suspension was then placed on top of the gelled Matrigel/CDM in each well of the chamber slide. Cultures were incubated at 37°C (5% CO2). The emergence of branching structures over an 8-day period was monitored by fluorescence microscopy.
Histology-fixation
Branching EBs and fetal pancreas rudiments derived from Matrigel explant cultures were fixed in 4% (w/v) paraformaldehyde (PFA) in PBS (pH 7.2) for 20 and 40 min at room temperature, respectively. Following fixation, samples were rinsed twice in PBS. Samples were then pelleted and embedded in 0.7% (w/v) low melt agarose. Agarose embedded samples were then dehydrated and embedded in paraffin wax. Blocks were sectioned at 5 μm, de-waxed, and processed for double indirect immunofluorescence labeling as previously described [30].
Immunohistochemistry
Following de-waxing of paraffin sections, heat mediated-antigen retrieval was performed by microwaving sections for 20 min in 10 mM Sodium Citrate, pH 6.0. Sections were allowed to cool for 20 min, followed by a brief wash in deionized water and rinsed twice in PBS. Sections were incubated for 30 min in 5% FCS in PBS containing 0.1% Tween and 0.5% BSA. The sections were incubated overnight at 4°C with primary antibodies as described in Supplementary Table S2. The chicken anti-GFP antibody (Abcam) was detected with an Alexa Fluor® 488 goat-anti-chicken IgG secondary antibody (Molecular Probes). The rabbit anti-Pdx1 (a generous gift from C.V.E. Wright, Vanderbilt University), rabbit anti-EpCAM (Abcam), rabbit anti-E-Cadherin (Santa Cruz Biotechnology), rabbit anti-Mucin1 (Abcam), rabbit anti-α-Amylase (Sigma-Aldrich), rabbit anti-Glucagon (Dako), and rabbit anti-Carboxypeptidase A (AbD Serotec) were detected with an Alexa Fluor® 568 goat anti-rabbit IgG secondary antibody (Molecular Probes). Nuclei were counterstained with 4', 6-diamidino-2-phenylindole (DAPI; Sigma).
Microscopy
Fluorescent and bright field images were captured on a Zeiss Axiovert 200 inverted fluorescence microscope (Carl Zeiss) and processed with Axiovision software. Confocal microscopy images were acquired using the Nikon C1 confocal laser-scanning microscope (40× and 63× oil immersion objectives). Data were processed using Image Processing and Analysis in Java (ImageJ) software.
Gene expression analysis
Total RNA was extracted and prepared using the RNeasy™ Mini Kit (Qiagen) according to manufacturer's instructions. E16.5 and E19.5 GFP+ sorted cells were dissected and isolated as previously described [29]. First-strand cDNA synthesis using random hexamer priming was performed with the SuperScript® III First-Strand Synthesis System for Real-Time-polymerase chain reaction (RT-PCR; Invitrogen). Quantitative gene expression assays were carried out according to the manufacturer's instructions using a 7500 RT-PCR System (Applied Biosystems by Life Technologies) with the following TaqMan® Gene Expression Assay probe sets as outlined in Supplementary Table S3. For each of the gene specific primer sets used, the signal was normalized against the expression of Gapdh as previously described [31], and the results referred to as relative gene expression. The results displayed are the mean±SEM derived from 3 independent experiments.
Statistical analyses
Data values obtained on the differentiation of EBs and pancreatic rudiments were subject to Student's t-test. Values of P<0.05, P<0.01, and P<0.005 were considered as statistically significant. Analysis of gene expression data was conducted using 1-way analysis of variance, and significant group differences were established with Tukey's post hoc comparison. Values of *P<0.05, **P<0.01, and ***P<0.005 were considered as statistically significant.
Results
Directed differentiation of mouse ESCs to pancreatic endoderm
We have previously described a multistep protocol for the differentiation of mouse ESCs to pancreatic endoderm cells in CDM [25,27]. This method entailed 4 days of treatment with low concentrations of BMP4 followed by a 24-h pulse of retinoic acid (RA) to induce the formation of Pdx1 + EBs. EBs developed over the 7 days with Pdx1-GFP+ cells being first visible at d8. In the current method, we replaced the BSA component within the CDM with recombinant human albumin (rHA), a modification that necessitated reassessment of the concentration and the timing of addition of growth factors required for reliable endoderm induction. In addition, we also included a low concentration of PVA, an additive we have previously shown to foster formation of EBs in 96-well low attachment plates (Fig. 1A). We observed that, unlike BSA containing CDM, rHA-CDM was unable to support efficient attachment of EBs transferred to adherent plates at differentiation d5. This difference could be obviated by addition of 10 ng/mL FGF2 at d5, a modification that promoted the outgrowth of an adherent stromal layer (data not shown). Although the role of this adherent layer was not specifically investigated, fluorescence microscopy demonstrated that, compared with controls, the overall percentage of Pdx1-GFP+ foci was ∼1.6-fold greater in FGF2 treated cultures (54.4%±3.7% vs. 33.9%±1.2%, mean±SEM, n=4 experiments) (Fig. 1B).
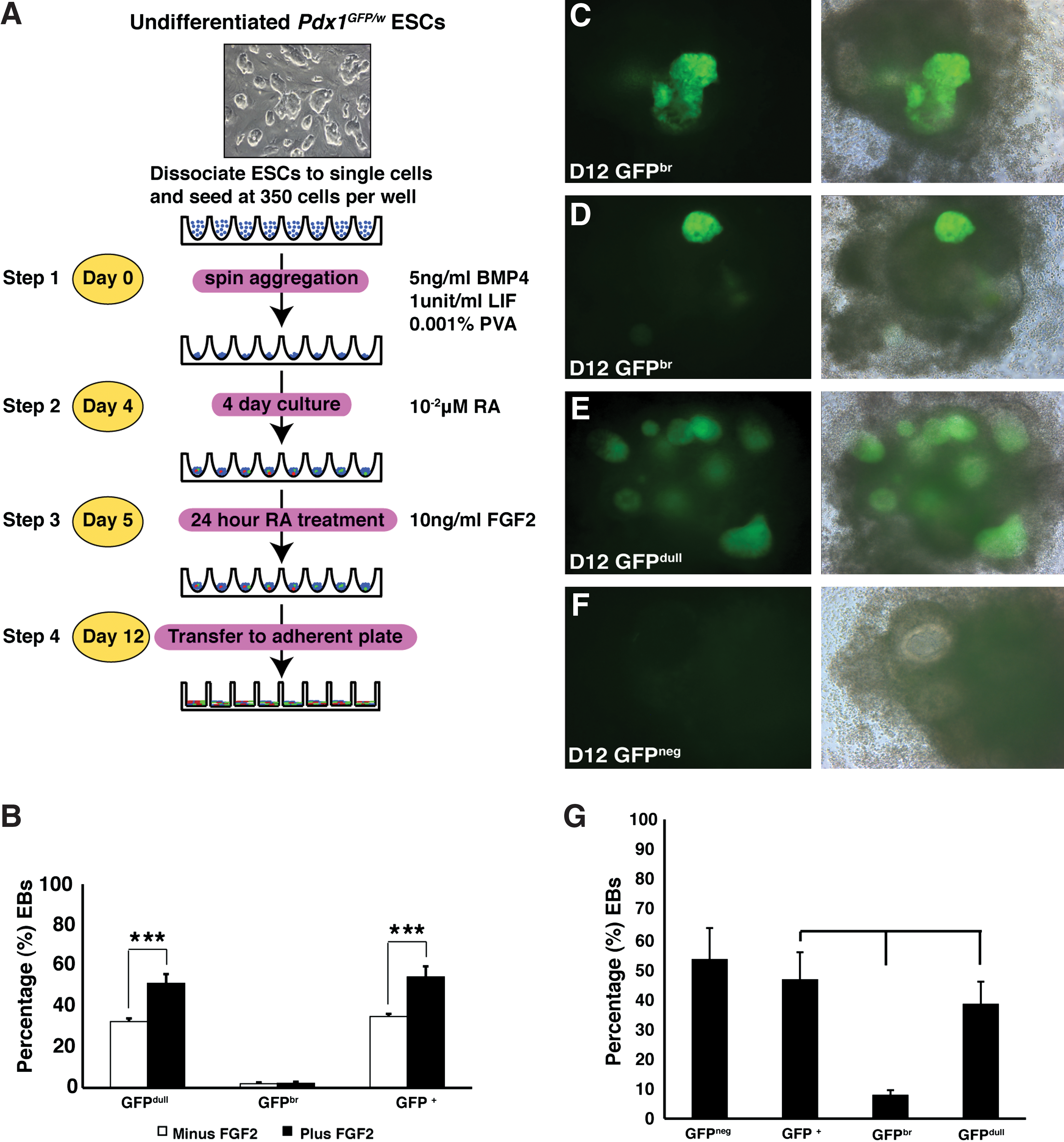
Protocol for the differentiation of ESCs to pancreatic endoderm.
In d12 Pdx1GFP/w EBs, 3 predominant morphologies were observed and subsequently scored using fluorescence microscopy (Fig. 1C–F); brightly fluorescing outgrowths designated as GFP bright buds (GFPbr, Fig 1C, D), diffuse GFP+ (Pdx1+) structures, referred to as GFP dull EBs (GFPdull, Fig. 1E) [25], and EBs that did not appear to contain any Pdx1-GFP+ endoderm (GFPneg, Fig. 1F). Using our modified protocol, the frequency of GFP+ EBs scored from d12 cultures was 46.6%±9% (mean±SEM, n=27 experiments) (Fig. 1G), similar to the frequency previously observed when permissive batches of BSA were available (53%) [25]. GFPbr buds and GFPdull structures were present in 8.0%±1.5% (mean±SEM, n=27 experiments) and 38.4%±7.4% (mean±SEM, n=27 experiments) of EBs respectively (Fig. 1G).
We have previously shown that GFPbr buds contained precursors of the exocrine and endocrine lineages [25]. In this current series of experiments, we asked whether these bright buds possessed another property of pancreatic primordia, namely, the ability to undergo branching morphogenesis in a Matrigel-based culture system [32]. To this end, we compared the branching ability of EBs containing GFPbr buds (d8, d10, and d12–d13) (Fig. 2A–D) and dissected embryonic pancreata from consecutive gestational ages (E11.5-E14.5) (Fig. 2E–H). Partially disaggregated EBs and fetal pancreata were cultured in GFR Matrigel diluted in rHA-CDM (Fig. 2). After 2 days in culture, cells from GFPbr EBs and fetal pancreas formed tight epithelial rings, which underwent a process resembling branching morphogenesis over the following 7 days. The ability of EBs containing Pdx1-GFP+ cells to undergo branching was optimal at specific stages of differentiation. For example, d8 GFPbr EBs (Fig. 2A) failed to branch, whereas d10 (Fig. 2B) and d12 GFPbr cells (Fig. 2C) formed convoluted GFP+ epithelia that increased in size and complexity over a 12-day and 7-day culture period, respectively. Finally, cells from d13 or older GFPbr EBs (Fig. 2D) possessed far more limited potential for branching (data not shown), and the structures that did develop displayed less intense GFP expression compared with that seen in those derived from d10 or d12 EBs. Correspondingly, in this culture system dissociated E11.5 derived Pdx1-GFP+ pancreatic rudiments (Fig. 2E) gave rise to poorly defined structures whereas E12.5 explants (Fig. 2F) had a propensity to form both cystic and duct-like structures, and exhibited some degree of branching. Pancreata from E13.5 Pdx1GFP/w embryos (Fig. 2G) underwent a process of growth and branching morphogenesis in which a highly elaborated tree developed over a 7-day period. Finally, E14.5 pancreatic explants had a reduced capacity to form branching structures and were more inclined to generate small clusters of brightly fluorescing GFP+ cells with limited potential for further growth (Fig. 2H).
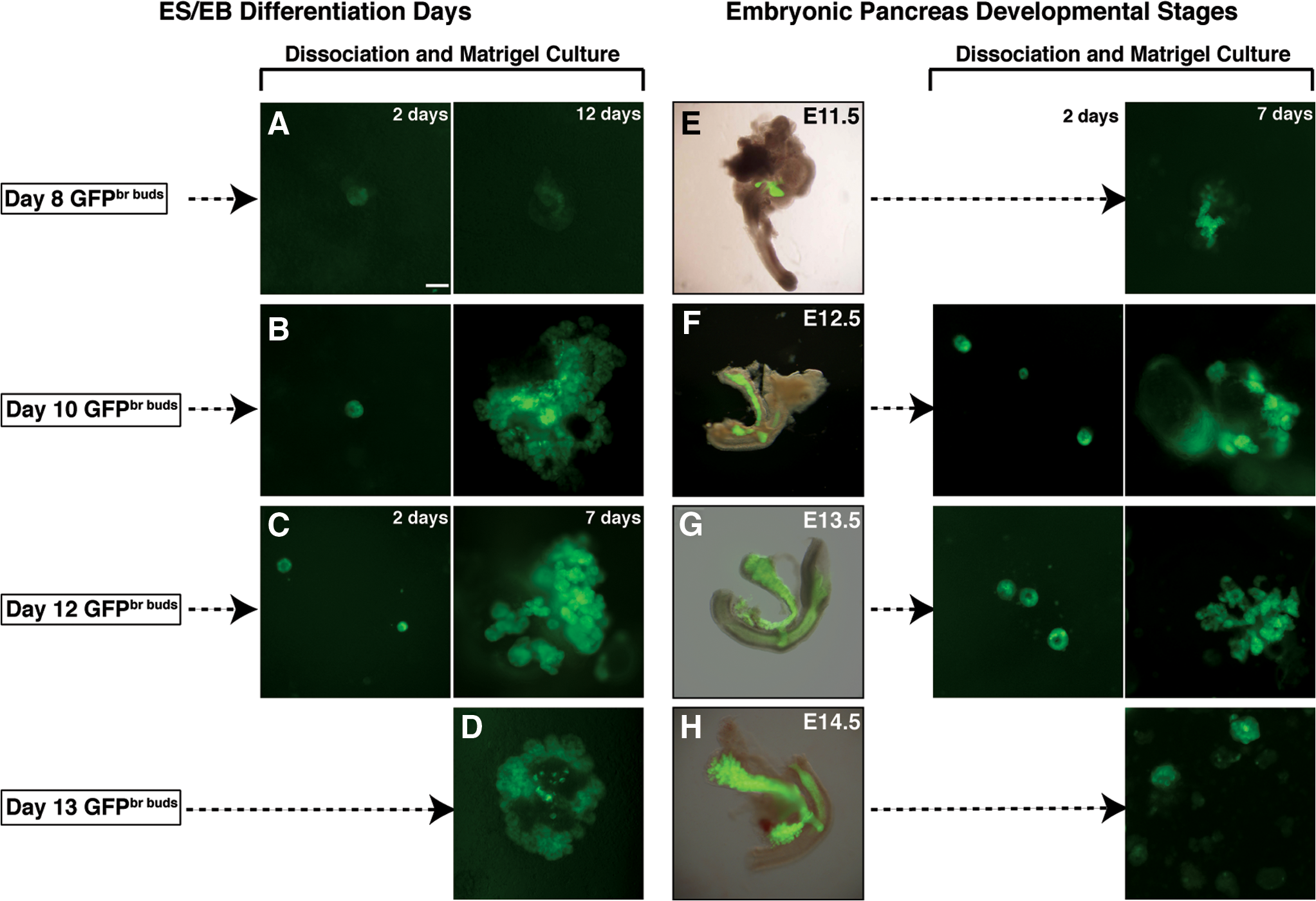
Comparison of branching potential of Pdx1GFP/w
EBs versus embryonic pancreata. Fluorescent images showing
The evolution of branching structures derived from GFPbr bud-containing EBs and pancreatic rudiments is further documented in Fig. 3. During the first 2 days following Matrigel culture, Pdx1-GFP+ cells from partially disaggregated E13.5 pancreatic rudiments and d12 GFPbr bud containing-EBs formed epithelial rings that were surrounded by GFPnegative (GFPneg) cells of mesenchymal appearance (Fig. 3A, D). On the fourth day of culture, small protrusions radiating from the epithelial clusters began to appear (Fig. 3A, D). In some instances, the tightly associated clusters of epithelial cells surrounded a small central lumen, which were indicative of the early signs of branching morphogenesis. From 6 to 8 days of culture, the epithelium expanded, with multiple branched structures forming a complex tree (Fig. 3A, D). Small independent lumina were always associated with these structures (Supplementary Fig. S1A–H). Beyond 8 days of culture, the fluorescence intensity of branching foci diminished and the epithelial clusters started to fragment, possibly reflecting exhaustion of nutrients within the culture medium. These same branching structures were not observed in cultures derived from GFPdull (Fig. 3E) and GFPneg EBs (Fig. 3F), nor did they arise from other Pdx1-GFP+ areas within the embryos, such as the distal stomach (Fig. 3B) and duodenum (Fig. 3C). Instead, explants of stomach and duodenum gave rise to tubular-like structures that were also surrounded by mesenchyme-like cells (Supplementary Fig. S2A–P). Moreover, the intensity of GFP expression associated with the structures that did develop was discernibly lower than that present in the pancreatic derived counterparts. In addition, the dimensions of these tubular-like structures, which were already pronounced by d4, continued to expand in size, such that by day 6, pulsating waves of peristaltic contractions could be observed (data not shown). This phenomenon was also apparent in explant cultures derived from the duodenum.

Explant cultures derived from E13.5 Pdx1+ pancreatic rudiments and d12 GFPbr buds both undergo branching morphogenesis when cultured in Matrigel. Brightfield/GFP overlay image of an E13.5 Pdx1GFP/w
embryo showing expression of GFP is restricted to the dorsal and ventral pancreatic buds, the proximal duodenum, and the distal stomach.
The frequency of branching structures observed in cultures initiated with in vitro and in vivo cell populations is summarized in Fig. 4. These results indicate that Pdx1-GFP+ pancreatic rudiments formed on average, 51.6±27.4 (mean±SEM, n=4 experiments) branching structures (Fig. 4A), while none were scored for the proximal duodenum and distal stomach explants (Fig. 4A). By comparison, GFPbr bud containing EBs yielded ∼31±12.6 (mean±SEM, n=6 experiments) branching organoids (Fig. 4B). As indicated, the number of similar structures derived from GFPdull or GFPneg EBs was negligible. Overall, these analyses vindicate the assignment of separate identities to EBs containing different levels of GFP expression and strengthen the conclusion that only those containing GFPbr buds contain pancreatic progenitors.
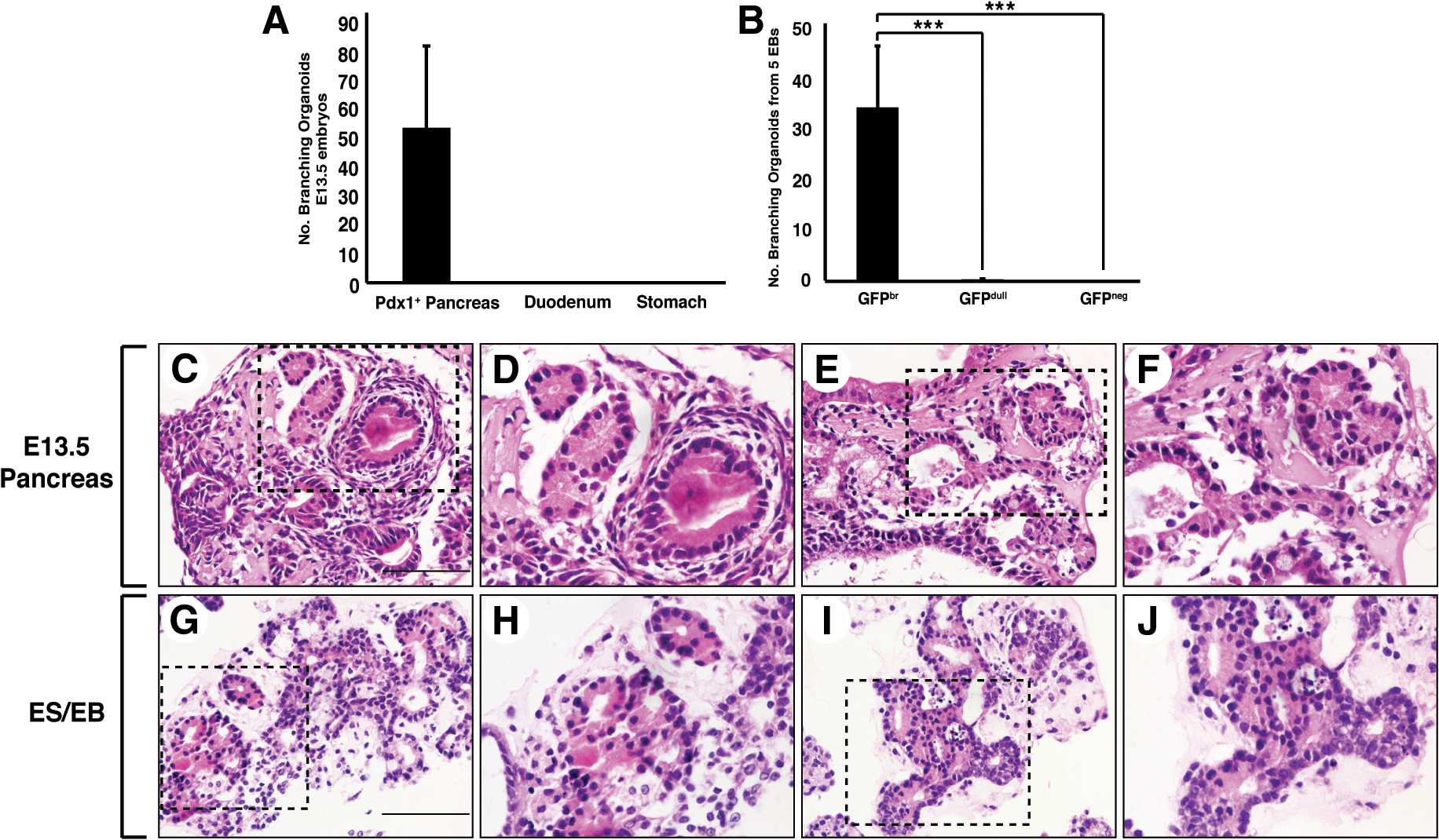
Branching frequency and histology of branching Pdx1-GFP+ foci derived from E13.5 pancreas and GFPbr EBs.
Histological analysis of GFP+ material collected from d12 Matrigel cultures indicated that branching structures contained cells of both epithelial and mesenchymal appearance. Branching organoids also characteristically contained multiple luminal epithelial ducts and adjacent to these, lobulated acinar-like structures (Fig. 4C–J). These small foci contained ductules with histological characteristics of acinar cells that were organized into discrete lobulated clusters containing abundant granular eosinophilic cytoplasm. These results, in combination with those described above, suggest that EBs containing GFPbr buds harbored precursors capable of giving rise to structures in Matrigel that resembled branching organoids generated from fetal pancreatic explants.
The results above suggested that d12 GFPbr EBs and E13.5 fetal pancreata contained pancreatic progenitors with a similar degree of developmental maturity. To explore this possibility further, we undertook a detailed analysis of branching structures derived from these 2 complementary experimental systems. In the first instance, we used immunofluorescence analysis to examine the expression of a cohort of pancreas-associated markers. Characterization of branching organoids derived from d12 GFPbr EBs and E13.5 fetal pancreata showed that the explant cultures contained cells expressing the epithelial markers EpCAM (Supplementary Fig. S3C, D) and E-Cadherin (Fig. 5A, B). These markers were expressed on both GFP+ and GFPneg cells, implying that Pdx1− epithelial cell types were present in the Matrigel cultures representing both in vitro and in vivo sources. Double staining of cells with antibodies recognizing GFP/Pdx1 (Supplementary Fig. S3A, B), Amylase (Fig. 5C, D), or Carboxypeptidase A1 (Cpa1) (Fig. 5E, F) revealed that cells expressing these exocrine markers formed localized foci that were integrated within the Pdx1-GFP+ epithelium. Branching structures derived from both EBs and pancreata also expressed Mucin1 (Muc1), an apical cell surface epithelial and ductal marker (Fig. 5G, H). Indeed, we observed that branching epithelial foci were always associated with small developing Muc1+ lumina (Fig. 5G, H). Similarly, scattered clusters of Glucagon+ cells were also detected in the GFP+ areas (Fig. 5I, J). Sparsely distributed single Somatostatin+ (SST) cells were only occasionally observed and these were not always confined to GFP+ epithelia (Fig. 5K, L). Significantly, no Insulin+ cells were detected in the branching explant cultures, even in instances where the beta cell differentiation agent, Nicotinamide, was added to the cultures (data not shown).

Branching organoids derived from E13.5 Pancreas and d12 GFPbr bud containing EBs express a suite of endocrine, exocrine, epithelial, and ductal markers. Confocal micrographs of branching organoids derived from either E13.5 pancreas or d12 GFPbr bud containing EBs were generated by immunolabeling with antibodies directed against E-Cadherin
The above immunofluorescence studies were complemented with quantitative (Q)-PCR gene expression assays, providing information regarding the overall gene expression profile of cells associated with selected GFP+ and GFPneg epithelial structures within the Matrigel cultures (Fig. 6 and Supplementary). Gene expression analysis of GFP+ and GFPneg cells isolated from E16.5 and E19.5 fetal pancreas demonstrates stage-specific expression profiles. Correlation of these profiles with in vitro generated cells enables elucidation of their representative developmental stage. Indeed, Q-PCR analysis of genes associated with pancreas development was enriched in E16.5 and E19.5 GFP+ sorted cells (Supplementary Fig. S4A–N). In this respect, the elevated levels of Foxa2 associated with structures derived from GFPbr bud containing-EBs most likely reflected the overall enrichment for endoderm when GFP+ areas were selected, rather than an increased frequency of endoderm within the cultures overall (Fig. 6A). Similar arguments can be made for the higher levels of Pdx1 expression observed in GFPbr bud and fetal pancreatic derived cultures (Fig. 6B). Collectively, these analyses demonstrate that criteria for selecting specific subpopulations within these mixed cultures enable the isolation of comparable material representing EB and fetal pancreatic derived cells.
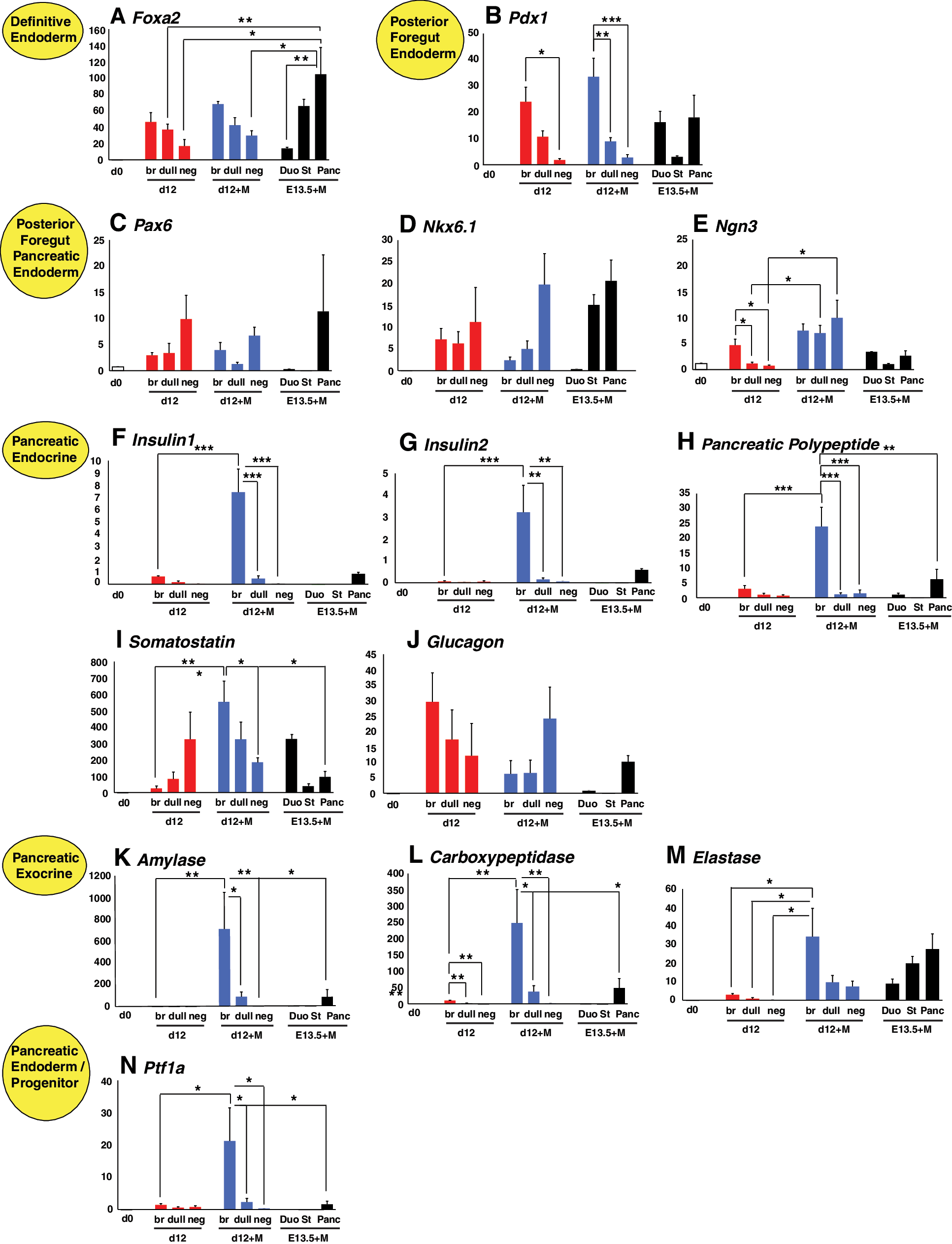
Quantitative-PCR analysis of genes associated with pancreas development. Genes that were assayed included markers of
Markers of pancreas differentiation, such as the transcription factors Pax6, Nkx6.1, and Ngn3 were also expressed at relatively low levels in both EBs and Matrigel samples compared with the levels observed in the E16.5 and E19.5 sorted fractions (Fig. 6C–E and Supplementary Fig. S4C–E). Interestingly Pax6 and Nkx6.1 were upregulated in GFPneg samples, perhaps suggesting that EBs contained other cell types in which these genes are expressed (Fig. 6C, D). Despite the fact that the expression of Insulin was not observed by immunofluorescence, branching organoids derived from GFPbr bud containing-EBs expressed detectable levels of Insulin1 and Insulin2. This result, combined with the observation that these organoids also expressed Pancreatic Polypeptide, Somatostatin, and Glucagon reaffirms the conclusion that pancreatic cells are enriched within the branching structures (Fig. 6F–J). This conclusion was also supported by the observation that expression of the exocrine markers Amylase, Carboxypeptidase, Elastase, and Ptf1a was also highest in samples representing GFPbr bud EBs (Fig. 6K–N). Statistical analysis of Q-PCR was also performed using analysis of variance (Supplementary Fig. S5). Interestingly, expression of exocrine markers in Matrigel cultures was substantially higher than that observed for the corresponding d12 EBs, suggesting that further culture in Matrigel resulted in enrichment or maturation of exocrine cells. Reassuringly, expression of these markers was also enriched in cultures derived from pancreatic rudiments relative to that observed in explants from the distal stomach and proximal duodenum.
Discussion
We have further refined a protocol for the differentiation of mouse ESCs to Pdx1+ pancreatic progenitors that arise in the context of structures that bear morphological similarities to the developing pancreas. Our method employed a chemically defined medium containing rHA, a modification that eliminated the high degree of variability associated with different batches of BSA previously used. Anecdotally, we had noted that some BSA batches were permissive for pancreatic differentiation while others were not. It is not clear whether these differences reflected pro-differentiative activities in the permissive batches or inhibitory factors in the nonpermissive batches. Nevertheless, in this current study we observed that FGF2 was required when using rHA-CDM to restore pancreatic differentiation to levels seen with “good” permissive batches of BSA-containing CDM.
Several previous studies have demonstrated the inductive role of notochord derived FGF2 during dorsal pancreas formation in chicken, mice, and humans [33 –35]. Consistent with these findings, addition of FGF2 in our protocol also increased the frequency of Pdx1-GFP+ endodermal cells. Although the mechanism of this effect was not investigated, we observed that FGF2 promoted the rapid outgrowth of mesenchymal cells after EBs were transferred to adherent plates (data not shown). It is therefore possible that this mesenchymal population produced factors that further enhanced the generation of Pdx1+ endoderm.
The ability of extracellular matrix to regulate the adhesion, migration, proliferation, differentiation, and maintenance of mature cell function has been well documented in the pancreas, most notably in β-cell development [36 –39], adult ductal and fetal pancreatic cells [40,41], and in ESCs [42]. In agreement with previous studies, we found that the commercial basement membrane preparation, Matrigel, proved an effective substrate for growth and expansion of Pdx1+ pancreatic progenitor cells via a process that resembled branching morphogenesis. The use of cultured pancreatic explants as a suitable model for studying branching epithelial morphogenesis has been meticulously documented [32,43 –48]. In the present study, we did not examine the mechanisms underlying pancreatic branching, however our observations of the branching dynamics for both ESC and embryo derived Pdx1+ progenitors were similar to those previously reported [32,43 –48]. We observed that disaggregated cell clusters derived from E13.5 Pdx1+ pancreatic rudiments and d12 GFPbr containing-EBs generated 3-dimensional epithelial structures that could be readily visualized and enumerated. In Matrigel, GFP+ cells formed clusters, which over time, progressively increased in size and differentiated into structures that appeared morphologically similar to acinar tissue.
Our EB explants contained a mixture of epithelial and mesenchymal cell types. Mesenchymal to epithelial signaling, mediated in part by soluble factors, plays a critical role in cell proliferation and differentiation of early pancreatic progenitor cells during pancreatic development [49,50]. Moreover, the correlation between mesenchymal signaling, epithelial morphogenesis, and cytodifferentiation were revealed in previous explant studies, in which there was a preferential allocation of cells to the endocrine versus the exocrine compartment depending on the presence/absence of mesenchymal signals [32,51]. These studies confirmed that mesenchyme surrounding the epithelium adopted an instructive role during pancreatic development. Our observations from branching experiments were similar to those previously reported by others, where depletion of mesenchyme resulted in the abrogation of branching [32,50,51] (data not shown). We have previously shown that day 12 GFPbr EBs treated with nicotinamide for 7 days are able to differentiate to insulin-producing cells in vitro [25]. However, using the Matrigel culture system described here, insulin expression was unable to be induced even in the presence of nicotinamide, indicating that these culture conditions are not permissive for the differentiation of insulin-producing cells (data not shown). This interpretation is supported by studies demonstrating that Laminin-1, a major component of Matrigel, promotes the commitment of multipotent pancreatic progenitors to the acinar cell lineage [47]. In addition, a more recent study by Semb and colleagues demonstrated that disruption of epithelial tube formation in the developing pancreas resulted in an increase in acinar differentiation at the expense of endocrine differentiation, largely as a consequence of changes in epithelial cell polarity and epithelial-ECM interactions [47]. In this light, the study of Semb and colleagues is consistent with our observation that our Matrigel culture favored acinar versus endocrine differentiation. Therefore, reduced endocrine differentiation may have resulted either from the pro-acinar affects of laminin-1 within Matrigel or from the disruption of epithelial tube formation during the dissociation of pancreatic explants and GFPbr EBs before Matrigel culture.
Although it was beyond the scope of the current study, future investigations will need to examine in greater detail the factors influencing the differentiation of these progenitors and whether the mesenchymal cells can be replaced by specific factors. In particular, Fgf10 [52] and laminin-1 [53, 54] have been shown to modulate the frequency and extent of branching morphogenesis in the absence and presence of an extracellular matrix.
Gene expression analysis of branching structures derived from both EBs and fetal pancreas indicated that our culture system favored the development of exocrine cell types. This was evident by the upregulation of markers such as Ptf1a/p48, Amylase, Carboxypeptidase, and Elastase, and the relatively low expression of genes associated with endocrine development. Although endocrine markers were present in branching structures, insulin+ cells were never observed even in explants derived from the fetal pancreas (data not shown and Fig. 6F, G).
In summary, we have described a system in which mouse ESCs are able to mimic aspects of the pancreatic developmental program that results in the differentiation of cells from embryonic endoderm to the generation of pancreatic progenitors of the endocrine, exocrine, and ductal cell lineages (Fig. 7). Our data supports the notion that developmental pathways that regulate differentiation of the pancreatic cell lineages in the mouse embryo may also be partially recapitulated during the in vitro differentiation of mouse ESCs. Insight and knowledge from our findings may extend existing strategies for pancreatic lineage specific genetic selection and provide a novel foundation for research into pancreatic differentiation and disease.

Summary model of branching morphogenesis from ex vivo derived E13.5 Pdx1+ pancreatic rudiments and in vitro cultured d12 GFPbr bud cells. At d0 of explant culture, small clusters of GFP+ cells coalesce, and these continue to generate bud outgrowths with the emergence of a central lumen. The continued proliferation of these bud outgrowths at d6, results in the formation of an arborescent epithelial mass with multiple lobules. By d8 the branching organoids contain precursor cells, which are representative of the endocrine, exocrine, and ductal cell lineages. Color images available online at
Footnotes
Acknowledgments
We would like to acknowledge Monash Micro Imaging Facility for the provision of instrumentation, training, and technical support from Mr. Stephen Firth, Mr. Chad Johnson, and Dr. Judy Callaghan and also Mr. Ian Boundy and Ms. Stefania Tombs at the Department of Anatomy and Histology, Monash University. We thank Professor Chris Wright for provision of the anti-mouse Pdx1 antibody. This work was supported by The National Health and Medical Research Council (NHMRC) of Australia and the Juvenile Diabetes Research Foundation (JDRF). AGE and EGS are both Senior Research Fellows of the NHMRC.
Author Disclosure Statement
The authors have no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.