
Abstract
Human adipose-derived stem cells (hADSCs) are an attractive source for cell therapies, because they can be obtained from aspirated adipose tissues with the capacity of proliferation and differentiation into several mesenchymal lineages under certain conditions. Sulfated polysaccharides, including heparin, modulate osteogenic differentiation of stem cells through the regulation of growth factor binding and signaling pathways. Here, we examined the effects of the sulfated polysaccharide fucoidan on osteogenic differentiation of hADSCs. Strikingly, fucoidan treatment resulted in increased alkaline phosphatase (ALP) activity and alizarin red and von Kossa staining. At the molecular level, fucoidan treatment enhanced the expression of osteogenesis-specific marker genes, including ALP, osteopontin, type I collagen, Runt-related transcription factor 2, and osteocalcin. Furthermore, fucoidan also promoted the osteogenic differentiation of another mesenchymal cell lineage, human amniotic fluid stem cells. These findings strongly suggest that fucoidan enhances osteogenic differentiation of hADSCs and possibly other mesenchymal cell lineages, indicating that it may be a potential candidate for promoting bone regeneration.
Introduction
H
Differentiation of hADSCs into osteocytes is regulated by endogenous and environmental factors [14 –16]. Recently, several reports have demonstrated that sulfated polysaccharides, including heparin/heparan sulfate and chondroitin sulfate, influence osteogenesis [17,18]. These polysaccharides were able to stimulate fibroblast proliferation and extracellular matrix deposition in vitro. In particular, some reports have shown the modulation of osteogenic differentiation by heparin. For instance, in a recent study, Jiao et al. demonstrated that heparin sulfate modulates BMP2-mediated transdifferentiation of C2C12 myoblasts into osteoblasts [19].
Fucoidan is a sulfated, fucosylated polysaccharide found mainly in various species of brown seaweed that exhibits some heparin/heparan sulfate properties, including cell proliferation and differentiation [20 –23]. Many of these effects can be attributed to stimulation of hADSC differentiation ability. However, scant data are available on the osteogenic differentiation effect of fucoidan, which has a molecular structure and functions similar to that of heparin. In the recent study by Changotade et al., it was reported that low-molecular-weight (LMW) fucoidan extracts showed potential effects on bone biomaterial osteoconductive properties [24]. However, the functions and mechanisms in osteogenic differentiation of hADSCs by fucoidan treatment have not been previously reported.
On the basis of these findings, the present study was undertaken to investigate the potential effects of fucoidan on hADSC differentiation into osteoblasts through the analysis of matrix deposition and osteogenic marker gene expression.
Materials and Methods
Fucoidan
High-molecular-weight fucoidan (>30 kDa) was purchased from Haewon Biotech, Inc. (Seoul, Korea,
Isolation and cultivation of hADSCs and amniotic fluid stem cells
hADSCs were isolated from subcutaneous adipose tissue based on standard protocols. Tissue specimens were obtained using a protocol approved by the IRB of the Kangbuk Samsung Hospital. The raw adipose tissue was processed according to established methodologies to obtain a stromal vascular fraction. To isolate ADSCs from adipose tissue, samples were digested at 37°C for 30 min with 0.075% type I collagenase (Sigma, St. Louis, MO), followed by inactivation with an equal volume of complete medium containing dulbecco's modified eagle's medium (DMEM; Gibco BRL, Gaithersburg, MD) supplemented with 10% fetal bovine serum (FBS; Gibco BRL) and 1% penicillin/streptomycin (Gibco BRL). After centrifugation for 10 min at 1200g, cells were resuspended in complete medium and filtrated through a 100 μm strainer to remove debris. The hADSCs were maintained at 37°C in a humidified atmosphere containing 5% CO2. To prevent spontaneous differentiation, cells were maintained at subconfluent levels.
Amniotic fluid (AF) samples were collected from women undergoing routine amniocentesis for genetic testing from July 2001 to November 2005. After centrifugation of AF (10 mL) at 1800 rpm for 10 min, the AF pellet was added to 2 mL of CHANG media (Irvine Scientific, Santa Ana, CA) in a culture dish. Informed consent was obtained from all participants.
Immunophenotype of hADSCs
The specific surface antigens of ADSCs in the cultures of passages 5–6 were characterized by flow cytometry analysis. Cells were trypsinized and then incubated with fluorochrome-conjugated antibody on ice for 40 min in the dark. After incubation, the cells were rinsed twice with cold PBS containing 4% FBS and 0.03g/L sodium azide and then analyzed on an FACS Calibur device (BD Biosciences, San Jose, CA). A total of 104 events for each sample were acquired. The following human antibodies were used: HLA-DR (FITC), CD34 (PE), CD45 (PE), CD73 (PE), CD90 (FITC), CD105 (FITC), and CD166 (FITC) (BD Biosciences).
In vitro differentiation of hADSCs
To confirm the differentiation capacity of hADSCs, we induced differentiation into each lineage. For in vitro osteogenic differentiation, cells were incubated in osteogenic medium (OS) consisting of low glucose Dulbecco's modified Eagle's medium (LG-DMEM) supplemented with 10% FBS, 10 mM β-glycerol phosphate (Sigma), 100 nM dexamethasone (Dex; Sigma), and 0.5 μM ascorbate 2-phosphate (Sigma). For in vitro adipogenic differentiation, cells were incubated in adipogenic medium consisting of LG-DMEM supplemented with 10% FBS, 5 μg/mL insulin (Sigma), 1×10−6 M Dex, and 0.5 μM 3-isobutyl-1-methylxanthine (Sigma). Medium was changed twice a week for 14 days. For in vitro myogenic differentiation, cells were incubated in MCDB131 medium (Welgene, Daegu, Korea) supplemented with 1% FBS and 100 U/mL heparin (Sigma).
Treatment
For fucoidan treatment, hADSCs obtained from 5 passages were replated at 4×103 cells/cm2 in 12-well and 100 mm culture plates. When culture plates became 80% confluent, the culture medium of each group were replaced with each respective lineage induction medium. Fucoidan was added to fuco-groups at a final concentration of 100μg/mL in osteogenic or culture medium, whereas the control group was maintained without fucoidan. We removed and replaced experimental and control media every 3–4 days.
For Dex treatment, after primary culture in the control medium and expansion for 2 passages, the cells were trypsinized and replated into 6-well tissue culture plates at a density of 105 cells per well. The cells were incubated in the control medium for 24 h to adhere them to the plates and then placed in osteogenic medium containing 100 nmol/L of Dex. The osteogenic medium was changed every 2–3 days.
Mineralization assays
For the alkaline phosphatase (ALP) activity assay, cells were fixed with formaldehyde and stained with naphtol AS-BI alkaline solution (Sigma) for 15 min at room temperature. Extracellular matrix mineralization was confirmed with alizarin red S (ARS) staining. Briefly, cells were fixed with 70% ice-cold ethanol for 1 h at 4°C and then stained with 40 mM alizarin red (pH 4.2) for 10 min at room temperature. Von Kossa (VK) staining was used to confirm calcium deposits. Briefly, cells were fixed with 100% ice-cold ethanol, washed in distilled water, and then covered with 5% silver nitrate for 1 h under light. After extensive washing in distilled water, the cells were exposed to 5% sodium carbonate solution for 1 min. The area of mineralization appeared black.
RT-PCR analysis of osteogenic marker genes
About 106 cells were harvested from each group by trypsinization after incubation for 14 days, and the harvested cells were washed with cold PBS solution and centrifuged at 800 g for 10 min at 4°C (more than thrice). We then isolated the RNA from hADSCs and differentiated cells using an RNeasy kit according to the manufacturer's instructions (Qiagen, Valencia, CA). For RNA analysis, equivalent amounts of total RNA served as the template for cDNA synthesis using reverse transcriptase (Invitrogen, Carlsbad, CA). The first strand of cDNA was synthesized from 1 μg of total RNA using a SuperScript™ III First Strand Kit (Invitrogen) according to the manufacturer's instructions. Gene expression was determined by reverse transcription-PCR (RT-PCR). We confirmed the expression of osteogenic-specific marker genes, including ALP, osteopontin (OPN), type I collagen (Col I), and osteocalcin (OCN). Primers used for PCR reactions are shown in Table 1.
ALP, alkaline phosphatase; OPN, osteopontin; Col I, type I collagen; OCN, osteocalcin; α-SMA, α-smooth muscle actin; PPAR-γ, peroxisome proliferator-activated receptor gamma; Runx2, runt-related transcription factor 2; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Results
hADSCs have immunophenotypes and differentiation capacities of typical mesenchymal stem cells
The expression of cell surface antigens was evaluated by flow cytometry on hADSCs. hADSCs showed positive expression of the mesenchymal stem cells (MSC)-specific surface markers CD73, CD90, CD105, and CD166, but did not express HLA-DR, CD34, and CD45, which are hematopoietic cell specific markers (Fig. 1A). It shows that hADSCs express the typical bone marrow-derived MSC immunophenotypes. Additionally, three of the hADSC cultures were tested for their ability to differentiate into other cell types. Beyond doubt, hADSCs were capable of differentiating into multiple lineages, including osteogenic, adipogenic, and myogenic cells. Moreover, we confirmed the specific marker gene expression [OCN for osteogenesis, α-smooth muscle actin (α-SMA) for myogenesis, and peroxisome proliferator-activated receptor-γ (PPAR-γ) for adipogenesis] of each lineage through an RT-PCR assay (Fig. 1B). These data indicated that hADSCs showed multi potentiality for differentiation to other cell types.

Stemness of hADSCs.
Fucoidan promotes osteogenic differentiation
Fucoidan is a highly branched, sulfated polysaccharide based mainly on L-fucose units with a similar chemical structure to heparin [25]. The ordered fucoidans mostly contain a linear backbone composed of (1→2)-α-L-fucose, (1→3)-α-L-fucose, and (1→4)-α-L-fucose [26,27]. A disaccharide repeat unit in the structure of fucoidan and heparin is diagrammed in Figure 2A.
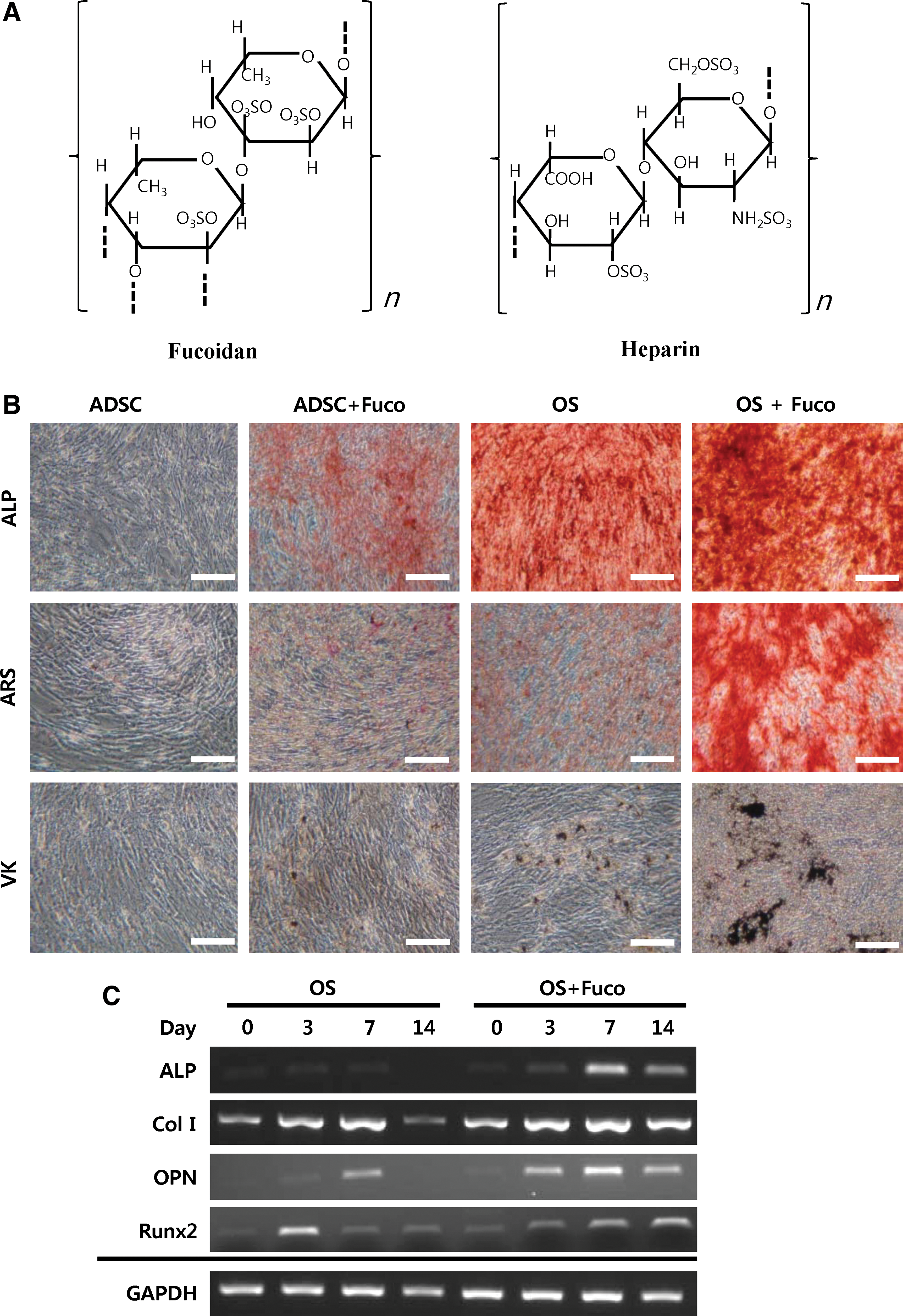
Fucoidan induced osteogenic differentiation of hADSCs. Cells were cultured in DMEM control medium or osteogenic medium (OS) in the presence or absence of fucoidan (Fuco) for 14 days.
To investigate whether fucoidan has an influence on osteogenic differentiation of hADSCs, we performed various staining assays associated with osteogenic differentiation. As shown in Figure 2B, hADSCs cultured with fucoidan in osteogenic medium (OS+Fuco) were strongly positive for ALP, ARS, and VK staining assays, as compared with hADSCs cultured in osteogenic medium without fucoidan (OS). These data indicated that fucoidan treatment prominently induced osteogenic differentiation of hADSCs, showing strong boosting effects of fucoidan on osteogenic differentiation. hADSCs cultured in DMEM with fucoidan (ADSC+Fuco) almost did not show specific reactions in any staining assays. In contrast, hADSCs highly expressed osteogenic marker genes, including ALP, Col I, OPN, and Runt-related transcription factor 2 (Runx2), when cultured with fucoidan in osteogenic medium (OS+Fuco) for 14 days, as compared with hADSCs cultured in osteogenic medium without fucoidan (Fig. 2C). These data further supported the results just cited.
In addition, we examined whether fucoidan has a synergistic effect on hADSC osteogenic differentiation when co-treated with Dex in osteogenic medium. Dex has been known to be an inducer for osteogenic differentiation of hADSCs [8]. Fucoidan-induced osteogenic differentiation showed no difference in the absence (OS−Dex) or presence of Dex (OS+Dex) (Fig. 3). Cells cultured in fucoidan-containing osteogenic medium (OS) without Dex (OS−Dex) for 4 weeks were markedly positive in both ARS and ALP staining assays, compared with the case in Dex-containing osteogenic medium (OS+Dex) in the absence of fucoidan. These data indicate that fucoidan may be an ideal substitute for Dex in osteogenic differentiation of hADSCs.
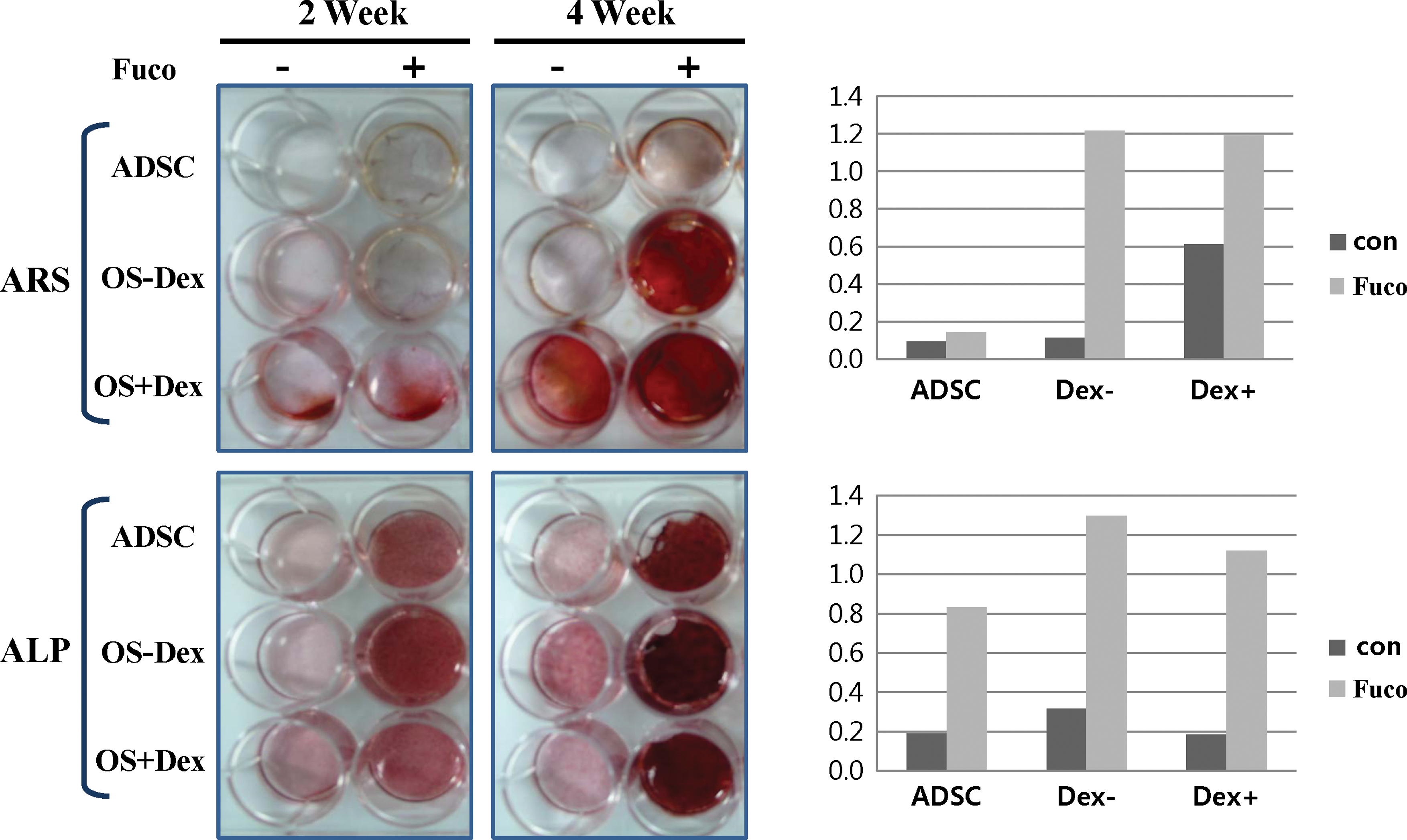
Effects of fucoidan (Fuco) on matrix deposition and ALP activity of hADSCs. Cells were cultured in DMEM medium either with or without fucoidan (100 μg/mL) or dexamethasone (Dex) (0.1 μM)/fucoidan-containing or not osteogenic medium (OS) for 14 or 28 days. hADSCs were stained by ARS or ALP for extracellular matrix mineralization. Color images available online at
Fucoidan also promotes osteogenic differentiation of other mesenchymal cell lineage, human amniotic fluid stem cells
Next, to confirm the beneficial effect of fucoidan on osteogenic differentiation of stem cells, we conducted the VK staining assay in human amniotic fluid stem cells (hAFCs). As shown in Figure 4A, hAFCs cultured with fucoidan in osteogenic medium (OS) were strongly positive in the VK staining assay over time, as compared with AF stem cells in osteogenic medium without fucoidan (Fig. 4A). These data suggested that fucoidan may promote osteogenic differentiation regardless of stem cell type. Additionally, to investigate the effect of fucoidan on differentiation into other lineages of hAFCs, we conducted the RT-PCR assay using either a myogenic α-SMA or an adipogenic PPAR-γ marker gene. As shown in Figure 4B, AF stem cells cultured in each induction medium containing fucoidan (Induction+Fuco) expressed the α-SMA gene to a similar level and the PPAR-γ gene to a slightly reduced level compared with AF stem cells in induction media without fucoidan (Induction). These results suggested that fucoidan may also induce osteogenic differentiation of other mesenchymal cell lineages.

Fucoidan also enhanced osteogenic differentiation of human amniotic fluid stem cells (hAFCs).
Discussion
The potential of the sulfated polysaccharide fucoidan on hADSC differentiation into osteogenic lineages
Our results showed that fucoidan significantly induced osteogenic differentiation of hADSCs, increasing expression of osteogenic marker genes, including ALP, Col I, OPN, and Runx2. hADSCs from adipose tissue are defined as multipotential adult stem cells and can be differentiated into various types of osteoblasts, chondrocytes, adipocytes, muscle cells, and neural cells [4 –8]. Our data were consistent with the reports just provided (Fig. 1).
Sulfated polysaccharides such as glucosamine sulfate, heparin, and heparin sulfate are known to be macromolecules connected with the cell surface and the extracellular matrix [18,19,28]. These polysaccharides have been shown to interact directly with numerous growth factors, including BMPs, and to stimulate fibroblast proliferation [19]. Fucoidan is one of the representative sulfated polysaccharides extracted from marine brown algae. Recent evidence has shown that fucoidan has numerous biological properties, such as antioxidative, anticoagulant, anti-thrombotic, anti-inflammatory, antithrombotic, and anti-tumoral activities [27,29 –31]. Furthermore, fucoidan was reported to induce osteoblastic cell differentiation, and LMW fucoidan promotes human osteoblast proliferation and collagen fibrillation [24]. Here, we have demonstrated that high-molecular-weight fucoidan strongly influences osteogenic differentiation of hADSCs. hADSCs treated with fucoidan were markedly positive in ALP, ARS, and VK staining, and additionally significantly expressed all the osteogenic marker genes (Fig. 2C). ALP plays a crucial role as a phenotypic marker for early stage bone formation, and increased ALP expression correlates with increased bone formation [32]. Col I is an early osteogenic marker and the most abundant ECM protein found in osteoblasts. Additionally, OPN is a bone-related marker gene during osteoblastic differentiation. Runx2 has been identified as the primary transcription factor for osteoblastic differentiation [33]. In vitro studies have shown that Runx2 positively controls Col I, OCN, and OPN expression [34]. These reports infer that Runx2 might play a very important role in osteogenic induction processes of hADSCs.
Mineralization of extracellular matrix, which is the last phase in bone formation, was assessed by ARS and VK staining assays. According to these results (Figs. 2 and 3), fucoidan increased mineralized extracellular matrix and Ca2+ deposition. These findings indicate that fucoidan-stimulated hADSCs differentiate into osteoblasts. Dex-containing osteogenic medium is usually used for osteoblastic differentiation of MSCs and hADSCs [8]. In this study, we found that fucoidan further induced osteogenic differentiation of hADSCs (Fig. 3), compared with Dex. Recently, researchers have wondered about the efficacy of glucocorticoids such as Dex, which mediates the osteogenic process of skeletal progenitor cells and processed lipoaspirate cells [7,35]. In this regard, fucoidan might be an ideal inducer, enabling Dex to excel in osteogenic differentiation of hADSCs.
The efficacy of fucoidan on osteogenic differentiation using human amniotic fluid-derived stem cells
In addition, we also confirmed the efficacy of fucoidan on osteogenic differentiation using hAFCs. hAFCs cultured with fucoidan in osteogenic medium were strongly positive in the VK staining assay. These findings suggested that fucoidan may induce osteogenic differentiation regardless of cell type.
As shown in Figure 4B, fucoidan did not induce other differentiation pathways of AFCs, although it slightly suppressed adipogenesis. Kim et al. showed that fucoidan suppressed adipogenesis via the mitogen-activated protein kinase pathway in 3T3-L1 preadipocytes, a finding that is in part consistent with our current data [36].
In conclusion, we could demonstrate that fucoidan plays an important role in osteogenic differentiation of adult stem cells. On the basis of these findings, this study suggests that fucoidan could be possible source as a therapeutic agent for bone repair and regeneration.
Footnotes
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science, and Technology (2009-0076696). The authors are grateful to Professor Il Hwan Oh (Catholic University, Korea) for helping with the flow cytometry analysis of hADSCs.
Author Disclosure Statement
The authors have declared that no competing interests exist.