
Abstract
Autophagy is a lysosome-dependent degradation pathway that allows cells to recycle damaged or superfluous cytoplasmic content, such as proteins, organelles, and lipids. As a consequence of autophagy, the cells generate metabolic precursors for macromolecular biosynthesis or ATP generation. Deficiencies in this pathway were associated to several pathological conditions, such as neurodegenerative and cardiac diseases, cancer, and aging. The aim of this review is to summarize recent discoveries showing that autophagy also plays a critical role in stem cell maintenance and in a variety of cell differentiation processes. We also discuss a possible role for autophagy during cellular reprogramming and induced pluripotent stem (iPS) cell generation by taking advantage of ATP generation for chromatin remodeling enzyme activity and mitophagy. Finally, the significance of autophagy modulation is discussed in terms of augmenting efficiency of iPS cell generation and differentiation processes.
Introduction
P
Mechanisms of cellular homeostasis are important for preventing cellular injuries that could lead to impaired cellular function and ultimately cell death. One of those mechanisms is macroautophagy (hereafter called autophagy), a lysosome-dependent degradation pathway that allows the recycling of damaged or superfluous cytoplasmic content, such as proteins and organelles. During autophagy, an isolation membrane, or phagophore, hijacks portions of the cytoplasm, giving rise to the autophagosome. This double-membrane vesicle subsequently fuses with the lysosome, where the enclosed material will be released and degraded by lysosomal enzymes. Such process yields cell metabolic precursors that can be used for ATP generation or protein synthesis, for example (Fig. 1) [15 –19].
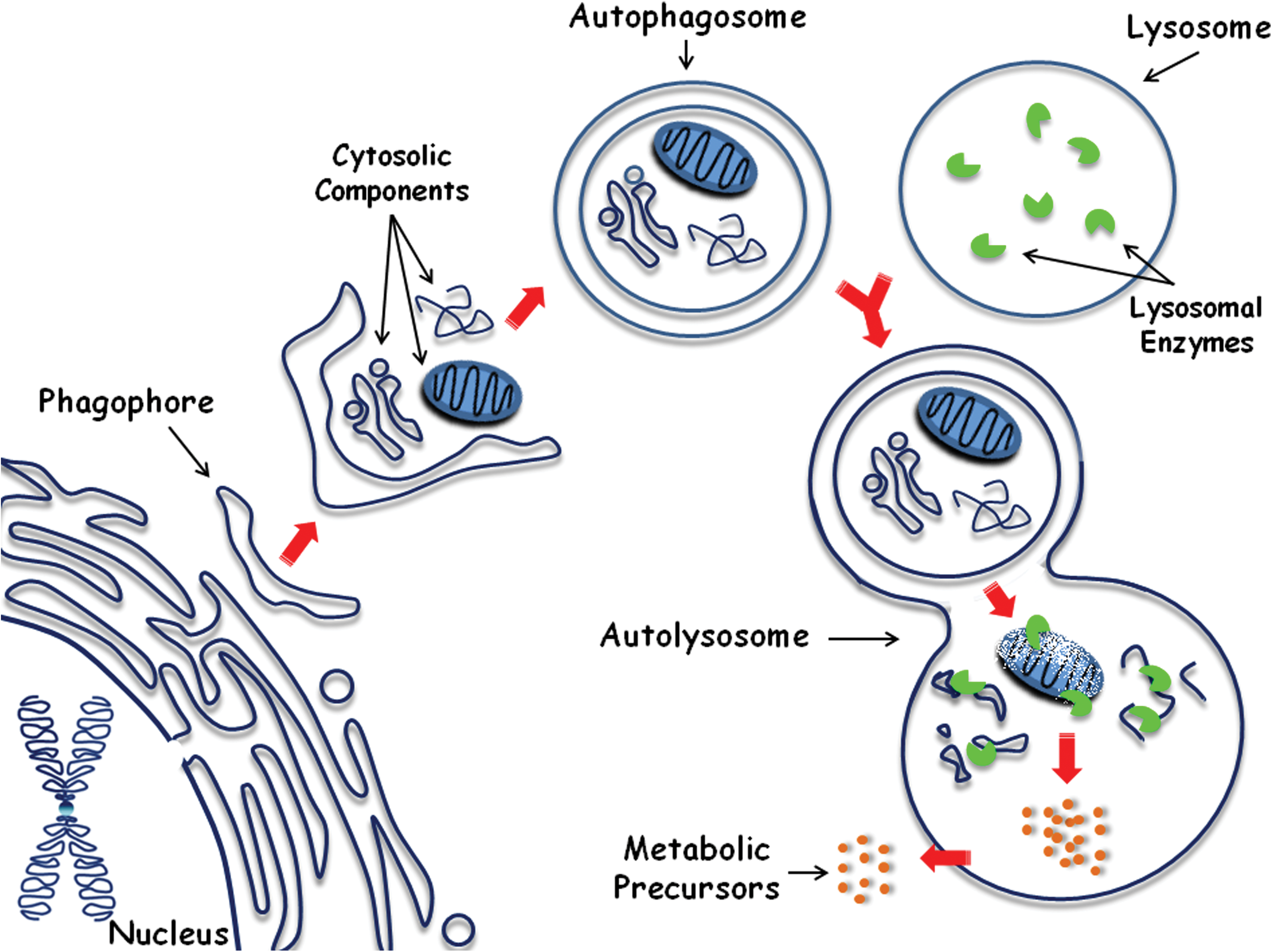
The autophagic degradation pathway. An isolation membrane (phagophore) sequesters cytosolic components into the autophagosome, a double-membrane vesicle that then fuses with the lysosome, originating the autolysosome. Inside of it, the captured material and the inner membrane of the autophagosome are degraded by lysosomal enzymes, thereby generating metabolic precursors that can be used for generating nutrients and energy (adapted from 15). Color images available online at
The autophagic pathway can be activated under different stimuli, such as starvation [17], endoplasmic reticulum stress [20], DNA damage [21], and reactive oxygen species (ROS) [22], thereby eliciting a cytoprotective response that helps cells to overcome those stressful situations. Deregulation of this pathway has been linked to several pathologies, such as neurodegenerative and cardiac disorders, cancer, and aging [15,23 –25].
In this article, we will summarize recent evidences showing that autophagy plays an important role in stem cell maintenance and differentiation. We will also discuss a possible role for autophagy during cellular reprogramming and iPS cell generation.
Autophagy Acts as a Cell Remodeling Mechanism During Cell Differentiation
As previously described, autophagy acts as an intracellular quality control mechanism through degradation of damaged or obsolete organelles and proteins. For instance, T lymphocytes deficient for the autophagy genes, Atg3, 5, or 7, show abnormal mitochondria accumulation and expanded endoplasmic reticulum due to impaired organelle homeostasis. As a result, there is an increase in ROS production and elevation in intracellular calcium that further impairs calcium influx, affecting the survival of T lymphocytes [26 –28].
Besides acting as quality control mechanism in differentiated cells, autophagy was also shown to participate in differentiation, as a cell remodeling mechanism that promotes morphological and structural changes. During adipogenesis, preadipocytes differentiate into adipocytes, a cell type that accumulates triglycerides inside a single and large cytoplasmic lipid droplet [29]. It was shown that preadipocytes deficient for Atg5 or Atg7, or treated with 3-methyl-adenine (3-MA), a pharmacological inhibitor of autophagy, originate smaller cells with decreased markers of adipocyte differentiation and reduced accumulation of triglycerides (Fig. 2A) [30]. Moreover, those triglycerides were stored inside small multilocular lipid clusters instead of a single large droplet. Authors noticed that those cells displayed a marked increase in mitochondria number and fatty acid β-oxidation levels, similar to what is found in brown adipose tissue cells (Fig. 2A). Similar results were observed by other authors who hypothesized that autophagy could be responsible for the cytoplasm remodeling activity that takes place during white adipocyte differentiation. Impairment of this pathway led to the accumulation of mitochondria, resulting in elevated rates of fatty acid β-oxidation. As a consequence, fatty acids were depleted, thus, impairing triglyceride synthesis that would accumulate inside the cell [31]. The authors also generated a mouse line carrying an adipocyte-specific Atg7 knockout allele, and observed that the animals were leaner, highly active, resistant to diet-induced obesity, and showed reduced white adipose tissue (WAT) mass. Low leptin levels and insulin resistance were also observed due to decreased WAT mass in these animals.
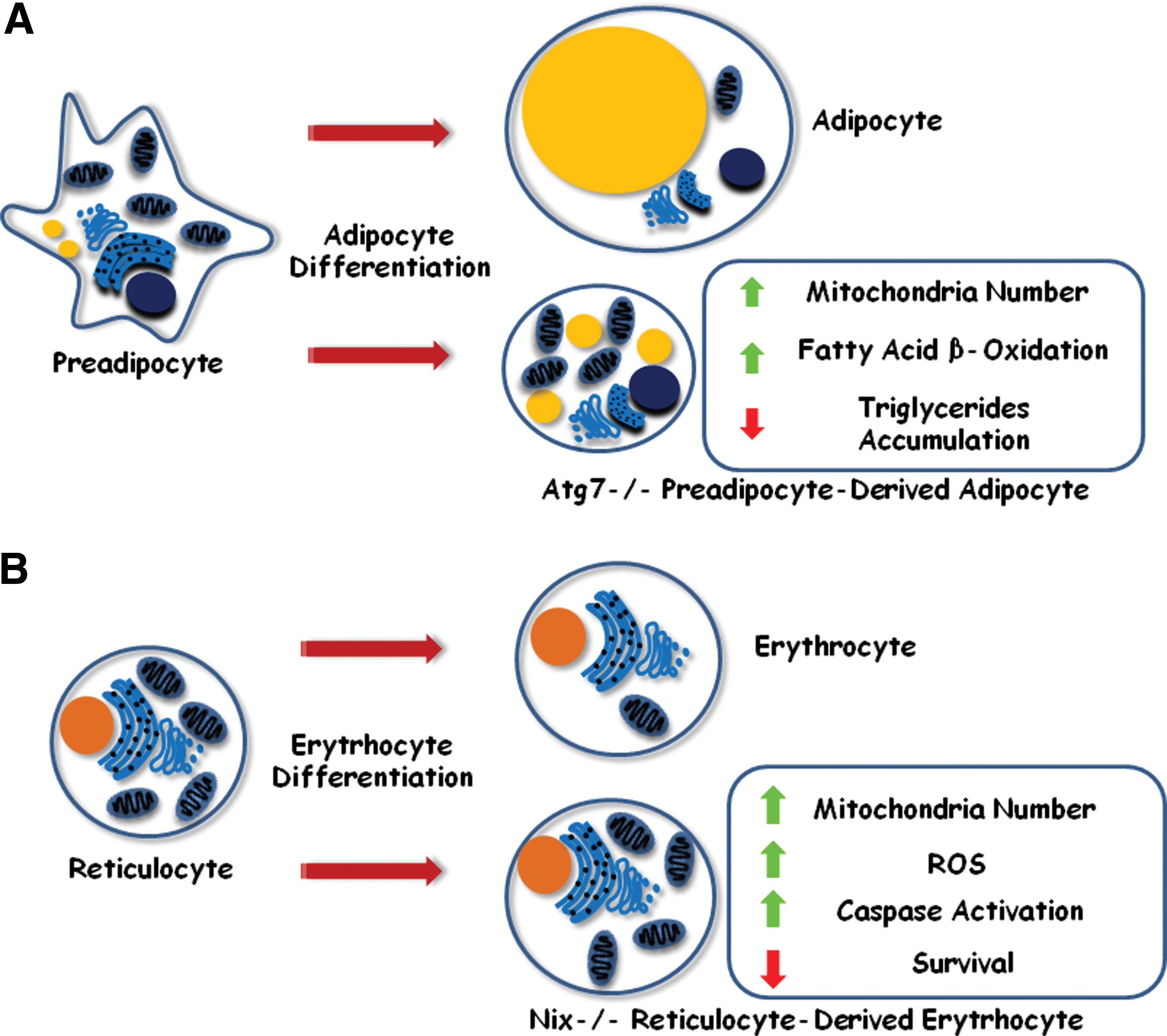
Autophagy is required during differentiation.
Similarly, it was also demonstrated that during terminal differentiation of reticulocytes into erythrocytes, mitochondria are eliminated in an autophagy-dependent fashion [32]. During this process, autophagy could selectively degrade mitochondria (a process called mitophagy) and that Nix, a BH3-only member of the Bcl-2 family, induces loss of mitochondrial membrane potential, an initial and critical step for selective engulfment of mitochondria into autophagosomes [33]. Because of defective mitochondrial removal, Nix−/− red blood cells (reticulocytes and erythrocytes) showed decreased survival in mice (probably resulting in the anemia observed in these animals), increased ROS generation, and caspase activation (Fig. 2B). The authors also described that pharmacological inhibitors of autophagy (3-MA, wortmannin, and chloroquine) also suppressed mitochondria removal in reticulocytes, confirming that this process is autophagy-dependent. Moreover, absence of Atg7 impairs mitophagy in erythroid cells, thereby leading to accumulation of mitochondrial superoxide in basophilic erythroblasts (Ter119+/CD71High), suggesting that defective mitophagy results in oxidative stress that may contribute to red blood cell death [34].
The importance of autophagy during cell differentiation is not restricted to mitochondria clearance, but may also extend to amino acid generation and possibly protein degradation. In support of this hypothesis, it was reported that during the oocyte-to-embryo transition, in the early mouse embryogenesis, autophagy is activated within 4 h after fertilization. Inhibition of this pathway (through Atg5 knockout) results in arrest at the four-to-eight-cell stage. The authors also noticed reduced protein synthesis (∼60% of that of the wild-type embryo), probably due to a lack of necessary amino acids provided by autophagy-promoted turnover of cytosolic content [35]. The authors also considered the possibility that autophagy could be responsible for degradation of oocyte-inherited proteins. In this case, defective autophagy would allow accumulation of maternally derived proteins and/or obsolete cytosolic material that could impair further zygote development.
Moreover, considering that some amino acids generated through the autophagic pathway may be used for protein synthesis, or even further processed in the tricarboxylic acid (TCA) cycle in order to provide intermediates that can be converted into nucleotides and sterols [16], it is tempting to speculate that autophagy may help cells undergoing differentiation to synthesize proteins and other cellular constituents, in addition to degrade proteins related to pluripotency maintenance that may impair differentiation.
Other studies support a clear role for autophagy during cell differentiation. It was described that hypoxia-mediated differentiation of RAW264.7 cells into osteoclasts is dependent on autophagy [36]. This process was activated in a HIF-1α/BNIP3-dependent fashion. Ablation of this pathway, using a specific HIF-1 inhibitor (YC-1), HIF-1α siRNA, or BNIP-3 miRNA plasmids, markedly prevented autophagy induction, as well as hypoxia-induced osteoclastogenesis. Autophagy inhibition using the pharmacological inhibitor 3-methyladenine or transfection with dominant-negative plasmids for Atg5 (DN-Atg5K130R) also resulted in abrogation of hypoxia-induced osteoclastogenesis.
Moreover, treatment with rapamycin, a drug commonly used to inhibit the mTOR complex (which regulates protein synthesis, cell growth, and also inhibits autophagy) or mTOR or raptor (regulatory associated protein of mTOR) silencing potentiates dbcAMP-mediated differentiation in the neuroblastoma glioma model NG108-15 cell line. The authors observed that rapamycin did not induce an increase in dbcAMP-mediated phosphorylation of p44/p42 ERK or CREB, although they have noticed an increase in cell cycle arrest, ATP generation, and autophagy upon treatment. Moreover, inhibition of the autophagic pathway by 3-MA or through silencing of the autophagy-essential gene beclin 1 abrogates the potentiation in NG108-15 differentiation mediated by rapamycin, revealing that autophagy might potentiate this process [37].
Autophagy upregulation was also noticed during keratinocyte fate commitment [38]. After inducing nonlethal stress by changing culture medium of HaCat (precancerous human keratinocyte) cells from KSFM™ (which is recommended for keratinocyte culture) to M199, the authors noticed dissociation of Beclin 1 from Bcl-X, an increase in LC3II, ATG5-ATG12 complex, and also in SIRT1, a NAD-dependent deacetylase that participates in cell differentiation and autophagy regulation. They also noticed an increase in the expression of the lysosomal enzyme cathepsin D, suggesting an increase in lysosome-dependent degradation activity during the process.
Although some autophagy genes and widely used pharmacological inhibitors of the autophagic pathway were shown not to be specific to this pathway [39], the data presented in here comprise different cell types, silencing of different autophagy-related genes, and use of different inhibitors. Thus, those results suggest that autophagy may play a pivotal role in a variety of cell differentiation processes.
Autophagy Protects the Genome and Helps to Maintain Hematopoietic Stem and Progenitor Cells
Stem cells need to protect their genome from damage to maintain their pool and self-renewal capacity [40]. It was also shown that intracellular ROS levels influence the long-term self-renewal capacity of hematopoietic stem cells (HSCs) [41,42]. In fact, the authors showed that HSCs deficient for Atm (a serine/threonine protein kinase involved in DNA damage response) exhibited high intracellular levels of ROS, reducing the long-term self-renewal capacity in a p38 MAPK-dependent manner. Moreover, long-term repopulating HSCs in murine bone marrow are highly positive for pimonidazole, a hypoxic marker, suggesting that low ROS levels help HSCs to maintain their self-renewal potential. [40] Embryonic stem (ES) cells exhibit fewer mitochondria and, consequently, lower reliance upon oxidative phosphorylation, resulting in fewer ROS generation (an important source of DNA damage) when compared with differentiated cells [43,44].
It was shown that beclin 1 +/− mice exhibit elevated incidence of spontaneous tumors in comparison with wild-type animals [45]. beclin 1 +/− or Atg5−/− cells engineered to express Bcl2 exhibited elevated DNA damage, gene amplification, chromosomal instability, and aneuploidy, suggesting a contribution for autophagy in maintaining genome integrity [46]. Also, autophagy-defective tumor cells accumulate p62 protein, leading to an increase in ROS levels, thereby contributing to tumorigenesis [47]. It was also proposed that autophagy may degrade defective mitochondria, preventing excessive ROS generation that could induce DNA damage and promote tumorigenesis (Fig. 3A) [48].
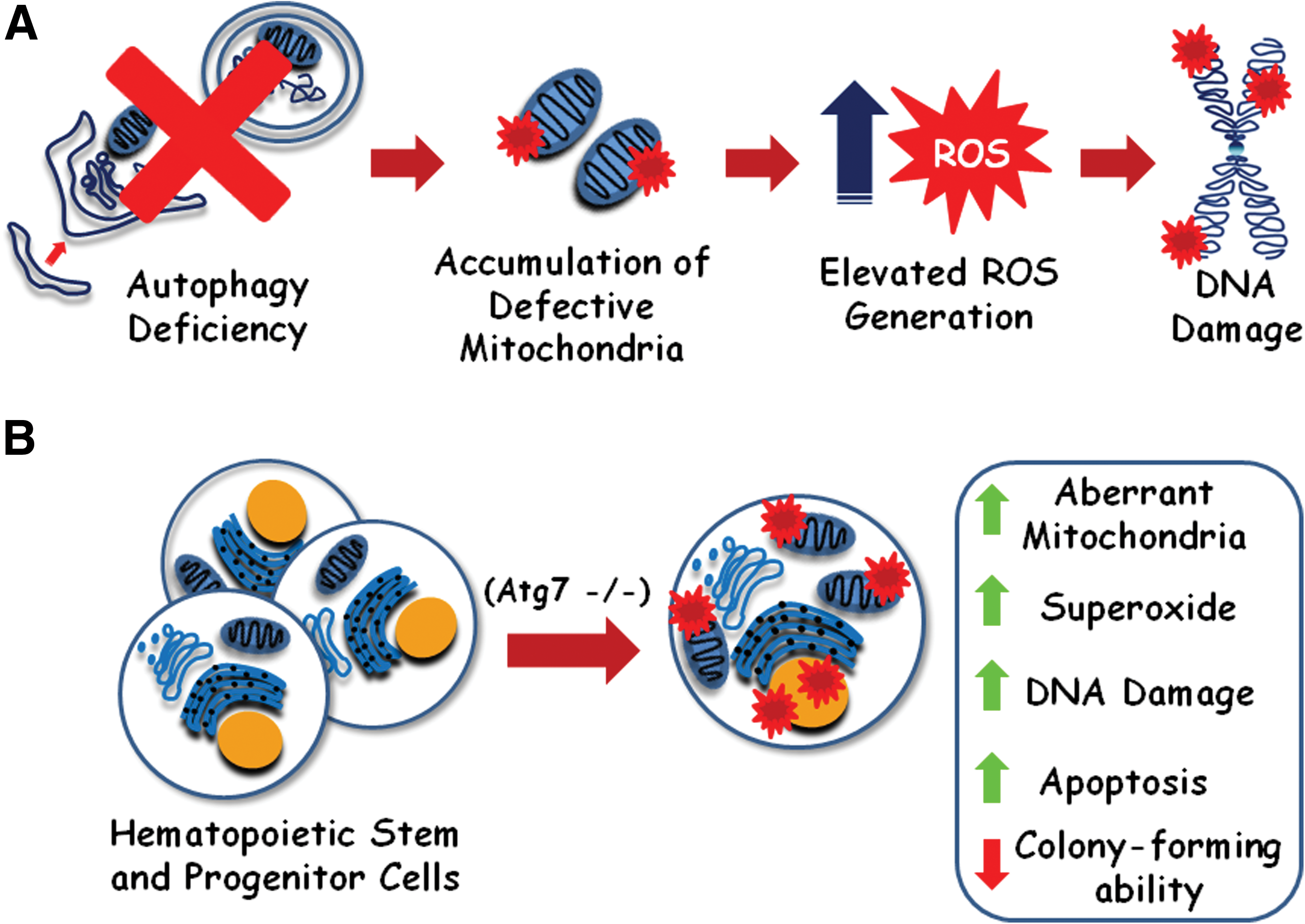
Autophagy is required for hematopoietic stem and progenitor cell (HSPC) maintenance.
In fact, it was shown that conditional deletion of Atg7 throughout the hematopoietic system in mice (Vav-Atg7−/− mice) results in anemia and lymphopenia [34,49]. The authors showed that Atg7−/− hematopoietic stem and progenitor cells (HSPCs) significantly accumulate more aberrant mitochondria, show elevated mitochondrial superoxide levels, DNA damage (53BP1), and apoptosis (caspase 3 activation) (Fig. 3B). They also showed that those cells failed to form secondary colonies in vitro. Moreover, in an assay to evaluate the reconstitution ability of those cells, Vav-Atg7−/− BM cells were transplanted into CD45.1+-irradiated hosts, failed to sustain a long-term hematopoiesis, resulting in death of the hosts within 4 weeks. Collectively, these results suggest that autophagy seems crucial to sustain HSC activity. The authors also have noticed that the frequency of myeloid (CD11b+Gr1+) cells was increased in the spleen and bone marrow of Vav-Atg7−/− mice, suggesting an atypical myeloproliferation. In fact, those cells were positive for the proliferation marker Ki67, and myeloproliferative infiltrates were found in myeloid and nonmyeloid organs in all Vav-Atg7−/− mice analyzed. Those results suggest that impaired aberrant mitochondria removal in Vav-Atg7−/− HSPCs led to elevated ROS generation, followed by DNA damage and subsequently genetic alterations that could confer Vav-Atg7−/−HSPCs a malignant phenotype.
Those studies support a clear role for autophagy in preventing genomic instability and maintaining adult HSC population, preventing disorders such as anemia, lymphopenia, and also cancer.
A Possible Role for Autophagy-Driven ATP in ES Cell Maintenance and Differentiation
During recycling of cytosolic material, autophagy may provide the cells with amino acids that can be oxidized in the TCA cycle, allowing the generation of FADH2 and NADH for the electron transport chain, supporting ATP production [15 –17]. This phenomenon was shown to be responsible for maintaining viability of cells deprived of nutrients and growth factors. Moreover, this autophagy-driven ATP production was shown to prevent mitotic catastrophe in glioma cell lines treated with the DNA damage–inducing agents Temozolomide or Etoposide [21]. Autophagy was also shown to support glycolysis-mediated ATP production in apoptosis-deficient cells with depolarized mitochondria [50]. These results suggest a protective role for autophagy-mediated ATP generation in cells undergoing different stressful situations.
It was also shown that autophagy plays a crucial role in programmed cell death during mammalian morphogenesis [51]. In this study, the authors observed that embryoid bodies (EBs) derived from beclin 1 or Atg5-deficient mouse ES cells fail to cavitate, a process in which the proamniotic cavity is formed when the solid embryonic ectoderm undergoes apoptosis. Although apoptotic levels and cavitation signals were normal in these cells, authors noticed that dying cells activate autophagy to generate ATP, which will be used in an energy-dependent mechanism to generate the “eat me” (phosphatidylserine) and to secrete the “come and get me” (lysophosphatidylcholine) signals. Failure to express those signals led to impaired phagocytic removal of dead cells, resulting in their accumulation in the center of the EBs and, consequently, impaired EB cavitation.
In ES cells, the pluripotent state is maintained by different mechanisms, such as key transcription factors (Oct4, Sox2, Nanog, and Klf4), miRNAs, and also by chromatin remodeling enzymes (CREs) [52,53,54]. Those CREs act in an ATP-dependent manner, disrupting interactions between histones and DNA, thereby inducing nucleosome conformational changes. These modifications increase DNA accessibility, allowing gene regulators to induce (or repress) gene expression.
SWI/SNF, ISWI, and CHD are 3 well-characterized families of ATP-dependent CREs. For example, BAF complexes (one of two major subfamilies that compose the SWI/SNF family) are ATP-dependent nucleosome remodeling complexes that were shown to both express and repress gene expression. A specialized embryonic stem (es)BAF complex (defined by the presence of Brg, BAF155, and BAF60A, and absence of BRM, BAF60C, and BAF170 subunits) has been reported necessary for self-renewal and pluripotency of ES cells [55]. In this study, Brg, the core ATPase subunit, was depleted (using shRNA-mediated knockdown) in murine E14ES, resulting in marked reduction in self-renewal and lost of colony morphology and alkaline phosphatase staining.
Besides maintaining the pluripotent state in ES cells, BAF complexes may change its subunit composition and mediate ES cells exit from the self-renewal cycle, allowing differentiation of these cells into neuronal precursors, for example [56].
Taking into account that autophagy provides cells with ATP for essential processes, and also for normal embryonic development, we hypothesize whether autophagy could provide ATP for the activity of those CREs.
Moreover, autophagy could also provide ATP for proper function of several other processes related to pluripotent-state maintenance. For example, ABCG2, an ATP binding cassette transporter of xenobiotic and chemotherapeutics agents that is highly expressed in hematopoietic cells and drug-resistant cancer cells [57], was shown to influence the self-renewal of mouse ES cells [58]. Authors described that inhibition of ABCG2 by Fumitremorgin C in these cells resulted in accumulation of protoporphyrin IX, which in turn leads to an increase in ROS levels, DNA damage, p53 and p53 phosphorylation (ser 18, 23, and 389), and downregulation of Nanog.
Although merely speculation, autophagy could help ES cells to maintain their self-renewal and pluripotency capacity, or also to differentiate, by providing ATP for CRE activity and ABC transporters, among other ATP-dependent processes.
Does Autophagy Play an Important Role in iPS Cell Generation?
In 2006, Takahashi and Yamanaka showed that introduction of 4 transcription factors (Oct3/4, Sox2, c-Myc, and Klf4) into mouse embryonic or adult fibroblasts was shown to reprogram differentiated cells to an embryonic-like state [6]. Those cells were named iPS cells, and exhibit ES markers and properties. For instance, ES cells have fewer mitochondria that are less active and less developed than those found in differentiated cells [43,44,59]. As a consequence, ES cells rely more on anaerobic than aerobic metabolism, generating fewer ROS that could induce DNA damage and, subsequently, lost of self-renewal capacity. Similar characteristics were recently described in iPS cells generated from human fibroblasts (Fig. 4), such as increased lactate production, reduced ATP generation, and decreased mitochondrial mass and number with an immature phenotype (round shape and underdeveloped cristae) [44,60,61].
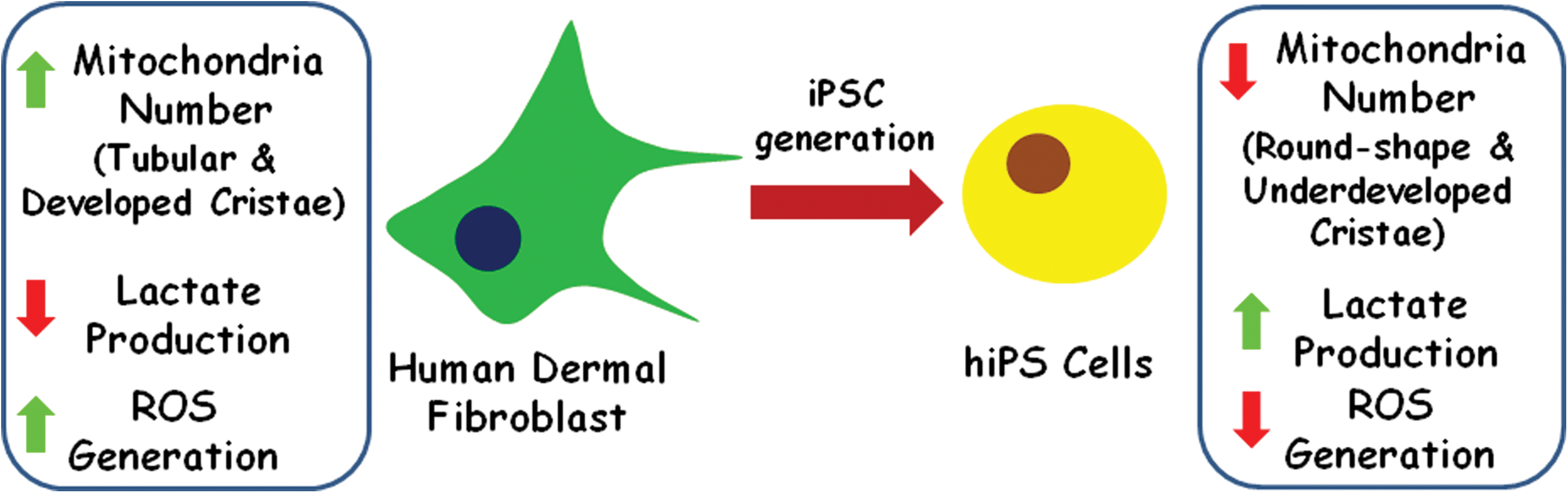
Variation of mitochondria content during genetic reprogramming to the pluripotent state. Induced pluripotent stem cells show reduced number of mitochondria, lower reactive oxygen species (ROS) generation, and increased lactate production than their respective somatic cells of origin. (Adapted from [61]) Color images available online at
The reduction in mitochondria number is likely to be responsible for the observed decrease in superoxide production in the generated iPS cell clones when compared with the fibroblast population from which they were originated. However, the process by which large amounts of mitochondria, present in the adult fibroblast, are eliminated during iPS cell generation is currently unclear.
We have previously discussed in this article that autophagy plays an essential role during differentiation of erythrocytes and adipocytes by promoting mitochondria degradation. Here, we hypothesize that autophagy could also play an important role in mediating remodeling of differentiated cells to a pluripotent state during iPS cell generation. In this scenario, autophagy would promote mitochondria degradation during iPS cell generation, allowing differentiated cells to reduce the amount of this organelle to an ES-like level. Moreover, autophagy could also promote degradation of proteins present in the differentiated cells that could impair (or slow) reprogramming, thereby eliminating proteins that should not be present in the pluripotent state. Autophagy-mediated turnover of proteins may also yield amino acids that could be used to boost protein synthesis. Taking into consideration that differentiated cells exhibit different proteome and organelle composition when compared with iPS (and ES) cells, it is tempting to speculate that autophagy may act as a cell remodeling mechanism not only during cell differentiation, but also during cell dedifferentiation.
A simple approach to tackle this problem would be to generate iPS cells from autophagy-deficient (Atg7−/−) adult dermal skin fibroblasts. If feasible, mitochondria mass and number, as well as presence of mature or immature mitochondria, and also superoxide levels, should be assessed and compared with the results previously described in this topic. An increase in developed mitochondria number and mass, as well as superoxide levels, in the autophagy-deficient iPS cells generated, would argue for a pivotal role for autophagy during reprogramming. Colony-formation assay followed by alkaline phosphatase staining [6,55,62] should confirm that autophagy-defective iPS cells may exhibit reduced self-renewal capacity due to increased ROS generation, for example. Protein synthesis rates could also be assessed, aiming at identifying whether autophagy-derived amino acids are being used in the newly synthesized proteins during the reprogramming process.
Moreover, human-derived iPS cells could be used as a tool to explore the importance of autophagy in various cell differentiation programs. The enormous variety of protocols describing methods to differentiate iPS cells into various cell types would allow us to study a more precise role of autophagy in several of these processes.
Concluding Remarks
Autophagy is a lysosome-dependent degradation pathway that allows cells to eliminate damaged or obsolete cytosolic components, such as proteins and organelles. Such mechanism provides the cells with metabolic precursors that can be used to maintain homeostasis during stressful conditions.
In this review, we discussed evidences showing that autophagy may also play an important role as a cell remodeling mechanism during cell differentiation and in HSPC maintenance. By degrading organelles and proteins, autophagy may help differentiating cells to degrade unwanted cytosolic material, such as mitochondria during adipocyte or erythrocyte differentiation. Moreover, by preventing accumulation of damaged mitochondria, autophagy contributes to keep low levels of ROS in HSPCs, thereby preventing cellular injuries that could contribute to loss of viability and self-renewal potential of these cells.
We also discussed a possible role for autophagy in mediating generation of ATP that could be used by ATP-dependent CREs, which participate in stem cell maintenance and in cell differentiation processes. Moreover, we suggest a set of experiments to investigate our hypothesis that autophagy may act as a cell remodeling mechanism during iPS cell generation, mediating organelle and protein degradation, thereby helping differentiated cells to achieve an ES-like composition. Finally, we suggest using iPS cells as a model to analyze the importance of autophagy in a wide variety of cell differentiation processes.
In light of the potential value of iPS cells for regenerative medicine, it is tempting to speculate that modulation of autophagy could become an essential component of protocols describing methods to obtain differentiated cells from iPS cells. In this scenario, tight regulation of autophagy during differentiation may have important outcomes, such as an increase in the efficiency of this process. In the same way, generation of iPS cells from differentiated ones may also be positively influenced by autophagy modulation. Finally, considering the differences observed between mitochondria of differentiated and ES/iPS cells, it is intriguing to imagine that transfer of iPS cell–derived mitochondria to somatic cells may also have a positive impact in reprogramming efficiency.
Footnotes
Acknowledgments
A.T. Vessoni would like to thank Dr. Carlos Frederico Martins Menck for the helpful comments on the manuscript. This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP-São Paulo, Brazil) and Conselho Nacional de Pesquisa (CNPq-Brasília, Brazil).
Author Disclosure Statement
No competing financial interests exist.