
Abstract
Tissue-engineered skin has a long history of clinical applications, yet current treatments are not capable of completely regenerating normal, uninjured skin. Nonetheless, the field has experienced a tremendous development in the past 10 years, encountering the summit of tissue engineering (TE) and the arising of stem cell research. Since then, unique features of these cells such as self-renewal capacity, multi-lineage differentiation potential, and wound healing properties have been highlighted. However, a realistic perspective of their outcome in skin regenerative medicine applications is still absent. This review intends to discuss the directions that adult and embryonic stem cells (ESCs) can take, strengthening the skin regeneration field. Distinctively, a critical overview of stem cells' differentiation potential onto skin main lineages, along with a highlight of their participation in wound healing mechanisms, is herein provided. We aim to compile and review significant work to allow a better understanding of the best skin TE approaches, enabling the embodiment of the materialization of a new era in skin regeneration to come, with a conscious overview of the current limitations.
Introduction
S
Skin TE went through a difficult time early in the 20th century, but with the commercial success that several products have achieved, it has entered a new area of enterprise [3]. Skin analogues have the longest history of commercialization, and the clinical applicability of cellular skin substitutes such as Apligraf® and Orcel® is significant [4]. However, among the currently commercialized ones, the major hurdles encountered are high production costs along with the failure of keratinocyte performance and delayed vascularization. Angiogenesis, relying in most of the cases upon patient's wound bed condition, is in fact a critical aspect for the success of skin analogues, achieved so far in substitutes with 0.4-mm maximum thickness [5]. Although deficient, there has been an emergence of skin TE models involving the generation, prior to implantation, of a microvascular network that anastomose with the patient's blood vessels and consequently lead to a faster vascularization [6 –10]. Commercial skin models have also been considered far from what would be suitable in terms of immunogenicity, as those containing nonautologous adult cells can induce immune rejection [11]. It is now clear that another obstacle in the long-term function of skin equivalents is limited by the terminal differentiation of the grafted keratinocytes due to prolonged in vitro culture that reduces the number of highly proliferative units (holoclones) [12]. Thus, in order to meet the need for maintaining keratinocytes in an early differentiation state, the revolutionary approach in skin TE comprises the use of stem cells, guarantying also an unlimited source of biological material, crucial for large full-thickness skin defects.
This review aims to present the rationale for the use of stem cells in skin regeneration, showing the current drawbacks and the different angles whereby these cells can contribute with their unique features.
Stem Cells Contribution to Skin Relevant Lineages
Distinct sources of stem cells (SCs), comprising both adult and embryonic stem cells (ESCs), have unique intrinsic features that might represent an effective way to meet the challenge of skin replacement and its current major drawbacks as presented above. This is due to foremost characteristics such as the potential to provide an unlimited source of donor material for grafting along with the ability to switch into any cell phenotype in vitro, such as endothelial and epidermal. In fact, improved vascularization is required, generally to allow faster inosculation and specifically to promote healing of chronic skin wounds. Moreover, epidermal analogues created from epidermal stem cells or stem-derived epidermal precursors have the advantage, in relation to fully differentiated keratinocytes in specific clinical settings, of being available in a shorter time due to the cells' higher proliferation capacity and at an earlier differentiation stage, which is expected to contribute to a higher quality healing and to skin regeneration rather than repair.
Overall stem cells characteristics have been extensively reviewed [13 –17]. This section aims to provide a critical and organized perspective of the works that have been carried out with both adult and ESCs, which justify the valuable potential of SCs from different origins to be further explored in skin TE purposes.
Stem cell origins
Mesenchymal stem cells (MSCs), a considerable part of adult stem cells, integrate several mammalian tissues and are natural key players in tissue regeneration. Although their primary function is homeostasis maintenance, they also seem to still express pluripotency markers, having the valuable potential to switch into different cell types other than into cell connective tissue lineages [18, 19] and represent a compatible source to allogenic transplantation due to their immunomodulatory and immunosuppressive features [20]. In addition to most well-described bone marrow MSCs [21], other relevant MSCs such as human adipose-derived stem cells (hASCs) [22], due to their abundance, ready accessibility, and easy expansion in vitro; human amniotic fluid stem cells (hAFSCs), which represent an intermediate stage between adult and embryonic SCs (ESCs) [23,24]; and umbilical cord blood (UCB), with its easy accessibility [25] and primal nature [26], are highly attractive for the skin regeneration field. Skin also shelters dermal MSCs, which have high proliferative capacity and the ability to differentiate into mesodermal lineages [27]. Their plasticity has been reinforced by the presence of pluripotency-associated markers [19,28].
However, the nature of MSCs is not fully clear; these cells are closely associated with perivascular niches [29] and coexpress many markers in common with pericytes in the microvasculature and adventitial fibroblast-like cells that surround the larger blood vessels [30,31,32], even though it remains possible that some MSCs originate in other cell subsets [33]. Furthermore, the dual origin of MSCs in a single tissue is suggested by the pericytes' capacity to differentiate into cells of mesenchymal origin, alongside with other MSCs of a nonpericytic origin [31]. In line with this, several studies revealed that human dermal fibroblasts, (hDFb) are difficult to distinguish from dermal MSCs, due to hDFb unexpected plasticity and immunoregulatory features [34,35]. Furthermore, it was recently reported that hDFb can generate, from a single cell (muse cell), others that form characteristic cell clusters expressing a set of markers related with pluripotency, raising the possibility to explore this feature benefiting from hDFb incapacity to form teratomas in vivo. Hence, despite the attempts that have been outlined for clarification [36], the limit to distinguish human dermal MSCs from hDFb ability is vague. For skin regeneration, the most important aspect relies on taking advantage of the classical features of fibroblasts and exploring the newly described qualities to achieve an improved skin model.
Besides the previously referenced skin MSCs, skin tissue also contains the skin resident naïve cells, representing a valuable cell source for skin TE. Their main niches include the apex of rete ridges [37] and the bulge of hair follicles [38] in the epidermis, and also the papillary dermis in the dermal compartment [39]. Epidermal stem cells (EpSCs) have a strong capacity for self-renewal and originate rapidly proliferating daughter cells—transient amplifying cells (TA)—that undergo several cell divisions before differentiating [40]. Despite the effort undertaken to characterize EpSCs and to assemble techniques that would allow better population purification [41 –44], specific markers of EpSCs are still questionable. While β1-integrin [42], the hemidesmosomal α6-integrin [44], and LGR5/LGR6 [45,46] are possible markers for stem cells in the hair follicle niche, early keratin markers such as K19, K14, and K15 are also shared with TA [47,48], though the distinct behavior of EpSCs and TA in vitro and in vivo have led to further define the retention of DNA label in vivo, and the in vitro clonal growth, as part of EpSCs' characterization criteria [49]. Within the dermis, hair-follicle sheath stem cells [50] and skin-derived precursors (SKPs) [51] have been recognized. While hair-follicle dermal sheath cells can be explored in a therapeutic context as an accessible option for improving the healing outcome of dermal or full skin equivalents, yet maintaining their role in the regeneration of hair follicles [52], SKPs, by sharing many characteristics with embryonic neural crest stem cells—including neural crest-like differentiation potential, in vitro and in vivo [39]—might be explored to promote the regeneration of skin nervous appendages. Nevertheless, optimization of protocols for human SKP isolation and differentiation into potentially clinically useful cell subtypes remains a challenging prerequisite to reach clinical translation [53].
ESCs are naturally the favorite candidates for regenerative medicine and TE due to their immortality, self-renewal ad infinitum, and highly versatile ability to differentiate into components of all embryonic germ layers (Fig. 1) [54], which might promote the use of these cells over adult stem cells. However, accurate and well-established differentiation techniques will be essential to the use of ESC-derived cells in future applications, namely in epidermis replacement [9,55 –61] and for improving vascularization [61 –66]. Further expectations rely on the development of efficient protocols to obtain melanocytes and promote regeneration of skin appendages, which would undoubtedly improve the overall functionality of skin substitutes [67].

KCL-002 human embryonic stem cell (hESC) cell line stained for pluripotency markers SSEA-4
Stem cells epidermal commitment potential
The commitment and differentiation of stem cells into epidermal/epithelial cells involve complex signaling pathways and multiple stimuli that occur naturally in an adequate microenvironment, but which have not been fully understood. Nevertheless, an effort to disclose this phenomenon has been explored in vitro [55,56,58 –60,68 –74] using distinct strategies, mostly focusing on MSC and ESC differentiation.
MSCs
In specific case of MSC differentiation toward keratinocytes, new methods has been explored for several stem cells such as bone marrow mesenchymal stem cells (BM-MSCs) [69,72], umbilical cord blood stem cells (UCBSCs) [68,71], and hASCs [73] and are mainly based on cytokine cocktails [56, 60, 68 –70], as well as on keratinocyte paracrine signaling [69,71,72,75] (Table 1).
Summary of the strategies employed to induce stem cells differentiation indicating the strategies employed and the main characteristics of the obtained cells.
ATRA, all-trans retinoic acid; BMP4, bone morphogenetic protein 4; EGF, epidermal growth factor; FGF, fibroblast growth factor; hESCs, human embryonic stem cells; HGF, hepatocyte growth factor; IGF, insulin growth factor; KGF, keratinocyte growth factor; MSCs, mesenchymal stem cells.
BM-MSCs were proven to give rise to epithelial-like cells, forming cobblestone pattern colonies and expressing both cytokeratin 18/19—when cultured with a growth factors cocktail that includes epidermal growth factor (EGF), keratinocyte growth factor (KGF), hepatocyte growth factor (HGF)—and insulin growth factor 2 (IGF-2) [72]. A higher expression of epithelial markers on the cells of clusters in the inner zone was observed, suggesting that most probably only these cells suffer differentiation due to the synergistic action of the growth factors and the stronger paracrine signaling, as described in other works [76,77]. Epidermal-like cells, expressing the early keratinocyte markers p63, cytokeratin 19, and β1-integrin and the late marker pan-cytokeratin were also generated from BM-MSCs [69]. In this work, cells seeded at higher densities demonstrated a more pronounced expression of those markers, implying that cell–cell contact, as expected, must be also considered as an important parameter to differentiate BM-MSCs into epidermal lineage cells. Although both studies [69,72] employed growth factors within the same concentration range, the role of each of these in epithelial and epidermal differentiation is yet undefined. Moreover, an analysis of stem cell markers, still lacking, would provide further information regarding differentiation efficiency of each strategy.
Findings with UCBSCs have underlined the potential of paracrine adult cell signaling and its effects on directing stem cell phenotype [71]. In this particular study, UCBSCs were co-cultured with adult keratinocytes on fibrin glue gel with encapsulated fibroblasts in vitro, and differentiation of UCBSCs into epidermal cells was detected. In addition, these results demonstrate that using this methodology the required initial number of both primary and stem cells is lower, as a mixture of both cell types was used, which might serve as a starting material for isolation and expansion of cells for transplantation in patients with large skin defects. A more recent work [68] also using hUCB as biological source of stem cells, investigated the use of a specific medium composed of a mixture of a basal medium and another that is normally used to expand keratinocytes, called the saigonese culture [68], to lead those cells into the epidermal lineage. After 7 days of culture, p63 and keratin1/10 positive cells were detected, meeting the postulated multipotential of UCBSCs. Nonetheless, issues such as isolation techniques employed, which result in low yields of the UCBSCs, and percentage of differentiated cells might be among the main concerns in the perspective of the clinical application of these cells.
A single in vitro study with hASCs showed that under the effect of all-trans retinoic acid (ATRA), cytokeratin 18 positive cells are generated, indicating hASCs' epithelial differentiation potential [73]. However, ATRA reduces cell viability in a concentration-dependent manner, and without the expression of typical keratinocytes markers, it is still insufficient to consider those cells for skin TE purposes.
All of these in vitro strategies have shown signal stem cells to change their gene expression patterns and transdifferentiate into epidermal/epithelial-like cells. However, in vitro assays to demonstrate this stratification capacity, as well as functional in vivo studies, are still lacking in most of the studies. Moreover, apart from a recent work that demonstrates that human umbilical cord lining epithelial cells are capable of generating a fully stratified epithelium [78], it is still necessary to verify whether the hMSC-derived cells have the same behavior as resident epidermal stem cells, notably in terms of multipotency and engagement in terminal differentiated/cornified cells to potentiate clinical application of stem cells in epidermal reconstruction.
ESCs
The possibility of human embryonic stem cells hESCs to differentiate towards the epidermal lineage has also been explored using distinctive strategies. Within embryoid bodies (EBs) or nodules produced in immunosuppressed mice [74,79,80], hESCs were able to differentiate along epidermal lineage, yet had a low proliferative capacity
A strategy based on the recapitulation of ectodermal development in vitro was proposed by Aberdam et al. [9,58,59,75,81]. Bone morphogenetic protein 4 (BMP4) effect along with the contribution of mesenchymal inducers (PA6 stromal cells) on ESC cultures was explored to generate cytokeratin 18/14 positive cells. The authors suggested this approach following the rationale of a coordinated development of the ectodermal and mesodermal layers during morphogenesis, leading the support and directing a number of biological functions including cell proliferation, migration, and differentiation by the reciprocal signaling of these two mutually dependent tissues [82]. This is consistent with the generation of keratinocytes from mouse ESCs seeded on matrix derived from human fibroblasts and exposed to BMP4 at different days of culture [83]. Knowing that epidermal and neural precursors are derived from the same neuroectodermal precursor [84], there was the need to clarify the successive development from ESCs to an ectodermal phase and the subsequent switching to epidermal precursors. For that, it was highlighted that the ectodermal commitment occurs through the activation of transcription factor ΔNp63, which seemed to play a dual role in early steps of epidermal precursor formation after BMP4 administration, acting as an epidermal inducer and an inhibitor of neural precursor formation [85]. In addition, ΔNp63 was demonstrated to be able to directly activate K14 promoter [86]. The importance of BMP4 in hESCs' differentiation pattern toward epidermal lineage, by blocking neural differentiation via Smad pathway [87] was also confirmed by obtaining a fairly pure keratinocyte population using retinoic acid along with BMP4, both in EBs and 2-dimensional hESC cultures (Fig. 2) [56]. The synergistic relation of the factors used softened the dilemma of neural versus epithelial specification among ectodermal precursors, giving rise to K14 positive cells. Moreover, the functional similarity of the generated keratinocytes to primary epithelium was demonstrated by showing that the obtained cells respond to microenvironment in a similar manner to adult keratinocytes [60]. Although some consistency has been demonstrated with the proposed strategies to obtain ESC-derived keratinocytes regarding comparable features with adult keratinocytes, at a protein level of expression and functionality, the reconstruction of a stratified epidermis is still lacking, with the exception of a study that used ascorbic acid and BMP4 for 40 days [57]. The authors claimed to mimic the long-term succession of biological steps that lead to epidermis formation during organogenesis generating functional basal keratinocytes with the ability to form a stratified epithelium that resembles normal human epidermis both in vitro and following grafting.
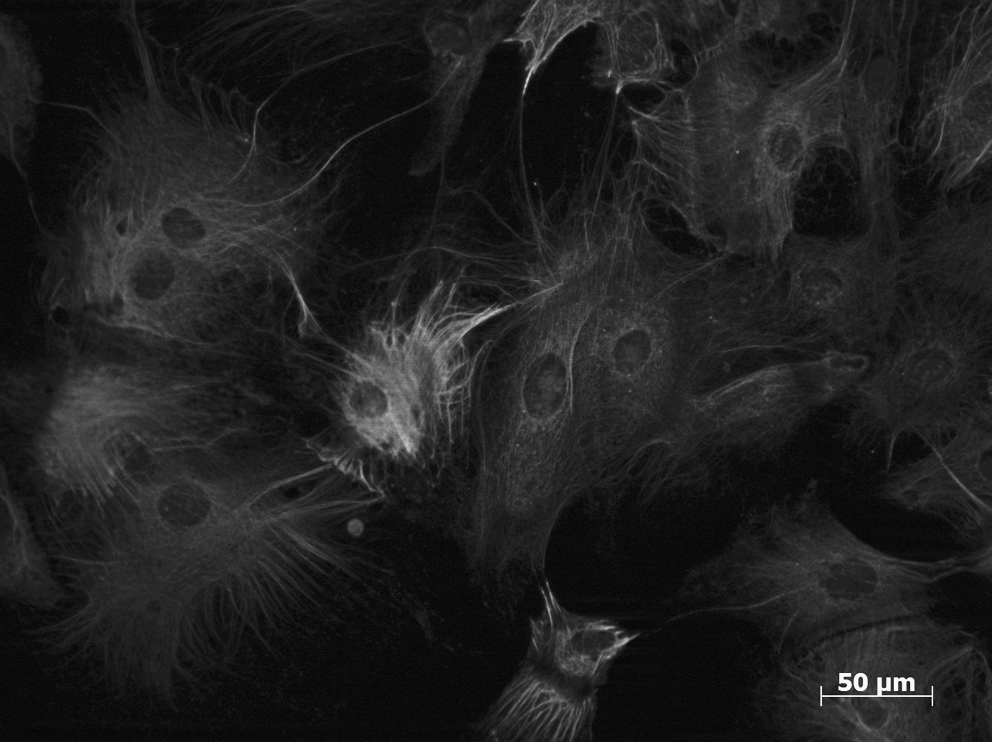
Keratinocyte precursors derived from KCL-002 hESC cell line, co-expressing keratin 18 and keratin 14, after treatment with retinoic acid and bone morphogenetic protein 4 for 25 days in culture.
Co-cultures of stem cells with adult cells, by taking advantage of the signal cues provided by these, are another promising approach to drive stem cell fate into a specific desired phenotype. Mouse ESCs directly co-cultured with mice keratinocytes were directed into the epidermal pathway as they were shown to express epidermal related markers, such as keratin 14 and k19, among others [88]. Despite the high potential of these studies, as the signaling provided by adult cells might be the key for phenotype switching, additional refinement of this methodology is required. Furthermore, transposing this approach to human cells together with functional tests, along with proteomics analysis, urges further exploration of the whole success of the strategy.
In what concerns epidermal fate irrespective of the SCs origin, there is still a long way to be explored in order to generate significant knowledge regarding the differentiation mechanisms involved. Moreover, the need to decrease the differentiation time-span to envision the clinical application of stem-derived epidermal/epithelial cells is still a reality.
Endothelial differentiation advances
Like for any other vascularized tissue, endothelial and endothelial progenitor cells also play a major role in skin regeneration, as they are responsible for the vascularization of the newly formed tissue. Together with pericytes, they ensure a stable and organized vascular network that supplies the whole tissue [89].
Despite the different approaches that have been employed to drive both adult and ESCs towards the endothelial lineage (see Table 2), no standardized protocols exist for generating stem cell–derived vascular cells [90]. The administration, in variable concentrations, of the most studied angiogenic cytokine, vascular endothelial growth factor (VEGF), [61,63 –65,91 –97] has been the most used route. However, distinct approaches such as co-cultures with other cells [98,99], SC exposure to mechanical stimulation [63,95 –97], and other microenvironmental cues [100 –103] also represent alternative attempts to accomplish this purpose.
bFGF, basic fibroblast growth factor; BMP4, bone morphogenetic protein 4; DiI-Ac-LDL, DiI acetylated low-density lipoprotein; ECGS, endothelial cell growth supplement; ECM, extracellular matrix; EGF, epidermal growth factor; EGM-2, endothelial growth media-2; hESCs, human embryonic stem cells; IL3, interleukin 3; IL6, interleukin 6; MSCs, mesenchymal stem cells; PECAM, anti-human CD31; PEG, polyethylene glycol; SVF, stromal vascular fraction; VCAM1, vascular cell adhesion protein 1; VE, vascular endothelial; VEGF, vascular endothelial growth factor; vWF, von Willebrand factor.
The endothelial differentiation of hESCs has been predominantly achieved by spontaneous differentiation of EBs grown for 10 to 13 days, which closely recapitulate that of early in vivo embryogenesis [61,65,66]. Differentiated cells are then sorted for VE-cadherin/PECAM and cultured in VEGF-containing medium [61] or, alternatively, directly cultured in methylcellulose in a medium cocktail including ascorbic acid and insulin/transferring/selenium [66]. hESCs in monolayer were also led toward the endothelial phenotype after subculture in collagen 4 substrate and supplementation with VEGF [65]. More recently, a novel method without cell-sorting technique that consists of culturing the generated EBs using a differentiation medium with VEGF, BMP4, stem cell factor, interleukin 3 (IL), and IL6 was reported [64]. The obtained cells were capable of supporting subculture at least up to 10 passages without phenotypical depression, and more importantly, effective engraftment in vivo.
Like for the differentiation of ESCs into the epidermal lineage, some works that intended to shed some light on the signaling mechanisms that mediate development of early hemato-endothelial progenitors during human development have also demonstrated its usefulness as promising approaches to explore the endothelial differentiation of ESCs [98, 99]. Those studies have generated hESC-derived cells with hematopoietic [98] and endothelial potential [98,99] by co-culturing hESCs in inactivated BM stromal cells, though after selecting the generated CD34+ [99] and CD34brightCD31+Flk1+ [98] subpopulations.
Despite the discussions and attempts to clarify the potential of ESCs and the mechanisms involved in endothelial differentiation [62,90], the understanding of the role of secreted and cell-bound factors associated with stromal cells used to promote that switch is still in an early stage. Further investigation and advances regarding the hESC culture might also be relevant to progress toward the control of lineage-specific and in particular endothelial differentiation.
As mentioned above, the differentiation of adult SCs, namely BM-MSCs, UC-MSCs, hASCs, and hAFSCs, has been predominantly relying on the use of culture cocktails with distinct growth factors, but commonly including VEGF [63,92,95 –97]. The variability of the results obtained, naturally arising from the numerous factors involved, nonetheless represents the difficulty in achieving relevant results as well as the long way still to go to establish reliable and clinically acceptable methods for obtaining endothelial cells from adult SCs. In fact, the concentration of VEGF, ranging from 5 to 100 ng/mL [104], constitutes a major drawback of these approaches and an impairment for scaling up.
Local environment, namely extracellular matrix (ECM)-originated biochemical and mechanical signals, is known to directly influence MSC differentiation [101,100,102]. Mechanical stress [63,95,96] in particular was proven to promote the endothelial differentiation of adult SCs, as well as the culture of stromal vascular fraction from adipose tissue in methylcellulose [105]. The importance of the synergistic effect of shear and growth factors/culture medium such as VEGF, endothelial cell growth supplement, and endothelial growth media-2 in inducing the expression of some endothelial markers was demonstrated. While the acquired phenotype varied among the tested sources, also differently stimulated, a common feature was the nonhomogeneous character of each one of the obtained populations regarding the expression of endothelial-related markers. Furthermore, the expression of important markers, such as endothelial nitric oxide synthases, or von Willebrand factor, was either not assessed [63] or absent [95], which might significantly compromise the in vivo performance and functionality of SC-derived endothelial-like cells.
Additionally, the effect of differentiation signals provided by ECM produced by endothelial cells as well of MSC alteration of these signals over MSC endothelial differentiation was evidenced by Lozito et al. [106]. It was also demonstrated that the BM-derived MSCs cultured in the ECM produced by macrovascular endothelial cells, eliminated by cell lysis, differentiate into endothelial cells supposedly by the action of some factors provided by the matrix, including VEGF, platelet derived growth factor (PDGF), and fibroblast growth factor (FGF) as previously identified [103], showing evidence of a feedback system in which MSCs are able to alter the very matrix signals acting upon them, releasing EC-differentiation factors from the matrix. Furthermore, the influence of the combination of growth factors, cocktail administration, and 3D polymer scaffold environment has also been highlighted in cell differentiation, namely in the formation of a 3D vessel-like network in vivo [100].
Overall, the endothelial-generated cells (see Table 2) depicted an endothelial phenotype in vitro, as proved by the typical markers; however, in vivo functionality assessment has only been demonstrated for some of the differentiation strategies [61,64,65,95,100,102,105]. A significant explanation for this might be the low yield of cells; as most strategies do not accomplish the full population differentiation, selection methodologies have been used to achieve homogeneous cultures, consequently with lower numbers. Improved culture methodologies are needed to overcome this issue and to allow the validation of the results by proving the vascular performance and integration of the differentiated cells in vivo.
Stem Cells Impact in Would Healing
Wound healing in postnatal human skin is one of the most complex biological processes. It involves a highly coordinated interplay among cells, soluble factors, and extracellular matrix, aiming at an effective wound closure [107]. The purpose of skin wound healing comprehends skin restoration and reestablishment of its tensile strength and natural barrier function [108]. Dysfunctional healing leads quite frequently to lifelong disability, having significant consequences and economic impact.
Wound healing repair versus regeneration
The distinction between repair and regeneration is crucial to understanding desirable wound healing progression, as elucidated in Fig. 3. Wound repair commonly refers to the physiological adaptation of an organ after injury, in an effort to reestablish continuity without considering the functional replacement of damaged tissue, whereas accurate tissue regeneration comprehends replacement of damaged tissue as an exact copy, in which both morphology and functionality are completely restored [109].
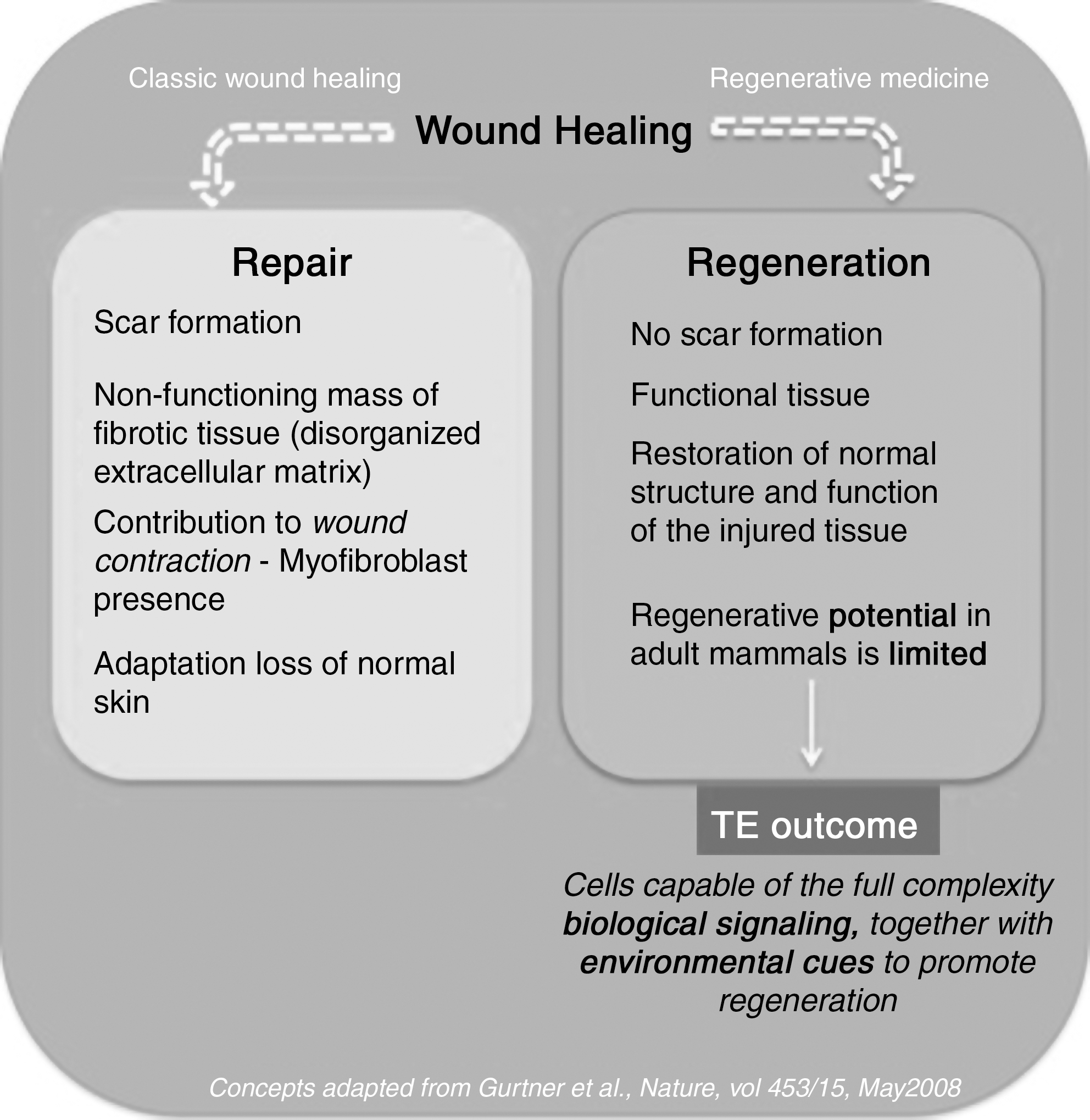
Schematic overview of wound healing, focused on the major events that distinguish repair and regeneration.
In adult skin, wound-healing responses can be physiological or regenerative, respectively leading to wound repair and wound regeneration. Although the understanding of how fetal skin shifts from a regenerative response in utero to a reparative response postnatally is still in its infancy [107], it might be the key for defining how adult skin healing may be redirected to the regenerative pathway instead of the wound repair–scar formation pathway.
Stem cells action
Stem cells, besides their plasticity and self-renewal capability, are known to enhance wound healing and reduce inflammation [110]. Although the extent of stem cell involvement in cutaneous wound healing is complex and not fully understood, these cells play a significant role in promoting wound vascularization, reducing wound contraction, attenuating scar formation, and inducing keratinization, and thus in better skin healing [111]. In fact, resident stem cells from epidermis and dermis are thought to promote reepithelialization, vascularization, and extracellular matrix remodeling through paracrine communication with resident, inflammatory, or antigen presenting cells and by transdifferentiation [112,113]. A strong example of this is the innermost role of EpSCs, not only in reepithialization [114], but also in the differentiation along adnexal lines [115], thus providing a superior source of multipotent stem cells to bioengineer sebaceous glands, along with formation of hair follicles [116,117].
Paracrine signaling and interactions
Paracrine interactions designate neighboring cellular communications. Stem cells are able to interact with their microenvironment, releasing multiple wound healing factors that present several advantages when compared with administered factors [118]. Additionally, MSC paracrine signaling regulates the local cellular responses to injury [119]. During cutaneous healing, recruited and resident stem cells are sensitive to the disruption of this microenvironment and thus respond accordingly. Although the mechanism and the signaling pathways responsible for MSC homing to wounded and uninjured skin are not fully understood, they probably involve the complex interplay of adhesion molecules, cytokines, extracellular matrix proteases, and tissue inhibitors of matrix metalloproteinase, forming a sophisticated signaling cascade [120].
There is increasing evidence [110,121 –123] that leads to the conclusion that MSC signaling is the main mechanism responsible for enhancing wound regeneration. It is thought that extensive skin injury, after the local stem cells response [124], promotes the early trafficking to the wounded region of a unique subclass of cells from bone marrow. These are the circulating bone marrow–derived cells, which include fibrocytes [125,126] that originate the myofibroblast population, essential to the wound closure [127]. Injected BM-derived MSCs also release IGF-1, EGF, and KGF that promote the proliferation of keratinocytes [110], and proangiogenic cytokines such as VEGF-a and angiopoietin (Ang)-1, which trigger an angiogenic response by local cells [123]. hASCs transplanted into a full-thickness skin defect have also been shown to enhance vascular density through paracrine mechanisms, along with an autocrine loop functioning, since, as suggested by the authors, local ASCs are likely to produce angiogenic factors that act on themselves or neighboring ASCs to reestablish vascularization at the wound site [128]. Interestingly, MSC-conditioned medium (MSC-CM), not only from bone marrow, but also from hASCs, appears to accelerate epithelialization, in a similar manner to what was observed when MSCs were transplanted [121,122]. BM-MSC-CM has also been implicated in the enhancement of wound neovascularization, by exerting potent chemoattractive and mitogenic effects that lead to increasing numbers of positive cells for CD34, C-kit, or Flk-1, thus suggesting recruitment of endothelial and endothelial progenitor cells into the wound [129].
The understanding of the effect of paracrine signaling on individual cell types, envisioning to disclosure the complexity in wound site interactions, has been addressed with in vitro studies. Meeting conclusions from in vivo data, BM-MSC-CM was shown to act as a chemoattractant for macrophages and endothelial cells. [119] Also, it has been demonstrated that BM-MSCs and BM-MSC-CM are effective chemoattractants for keratinocytes and dermal fibroblasts, proving its importance in the recruitment of these two types of resident cells [130] Furthermore, MSC-CM stimulates their proliferation, along with endothelial cells, through secreted mitogenic factors [121,130,131].
Transdifferentiation
MSCs have the ability to differentiate into cell types other than mesenchymal cell lineages [69,72,132], including into tissue-specific cells, as a response to cues provided by the specific niche of different organs [39,118,133]. This transdifferentiation phenomenon has been observed in skin wound regeneration in which MSCs gave rise to critical skin-related cell lineages such as epidermal, endothelial, and pericytic [69,72 128,132,134 –136]. This was particularly detected when GFP transgenic mice BM-MSCs and hASCs seeded in silk-fibroin-chitosan scaffolds were respectively injected and transplanted into full-thickness mice skin wounds [128]. Moreover, labeled UCBSCs applied locally at the wound margin of cutaneous mice wounds were not only found to accelerate the speed of wound healing but also to transdifferentiate into keratinocytes, identified in the newly formed epidermis [69,71,72,136]. Yet, another study shows that BM naturally contributes to a significant percentage of dermal fibroblasts in the skin, mainly associated with hair follicles, but rarely to the newly formed epidermis [113].
Despite all those indications, it is still controversial whether MSCs significantly contribute to wound healing via transdifferentiation [132,137]; this uncertainty might be explained by the requisite of a suitable microenvironment that not only promotes specific transdifferentiation, but also sustains the viability of the transplanted cells, promoting the formation of whole biologically functional tissue. Another explanation for stem cell plasticity resides in the cell fusion phenomenon. [138,139] Unlike what frequently occurred ex vivo [140], there is evidence that fusion might not be the primary mechanism [141], and transdifferentiation is believed to be the predominant event in vivo [110]. Nonetheless, the phenomenon of cell fusion occurrence, versus direct differentiation, in stem cell therapy is not fully understood and the debate persists.
Clinical Trials and Stem Cells Contribution to Wound Healing
Several in vivo studies [142 –145] have highlighted the potential of stem cells to treat skin wounds by accelerating wound healing where other methods failed before, making the statement as a strong therapeutic alternative and raising advances towards clinical trials. This data collection motivated the arising of MSC-based therapies for treating skin tissue wounds, which is reflected by the 90 clinical trials currently listed in the United States National Institutes of Health registry [120]. Despite this enormous number of clinical trials currently ongoing, only a set of studies that represent the potential of distinct adult stem cell sources on the treatment of several range of skin wounds categories will be herein discussed.
Recent studies have reinforced the significant impact of BM-MSCs [146,147] in nonhealing wounds, as described in subsequent sections, showing their active contribution to wound closure, reepithelialization (by differentiating into keratinocytes), neovascularization, and appendages regeneration [110].
A major advance in the treatment of a chronic nonhealing wound (diabetic ulcer) has been made [148] using the patient's BM-MSCs in combination with autologous skin fibroblasts on biodegradable collagen membrane (Coladerm®). Wounds showed a steady overall decrease in size and an increased dermal vascularization and thickening of the wound bed along the treatment. Furthermore, a clinical trial with a BM-MSC CD90 enriched population was also undertaken in diabetic patients with critical limb ischemia [147]. Cells were administered intramuscularly or intra-arterially, leading to revascularization of the affected limb and local perfusion enhancement, demonstrating once more the relevant role of these cells in critical wound regeneration.
On another perspective, the potential of the EpSCs isolated from hair follicle to improve wound healing in full-thickness burn patients is also being explored in clinical trials following different approaches. In one strategy, hair follicles were implanted in the commercial available dermal substitute Integra™ [149] with a novel technique of micrografting that resulted in complete reepithelialization and a hair-bearing scalp without the need for a split-thickness skin graft. Another approach, in phase II clinical studies, uses fully differentiated autologous epidermal equivalent derived from keratinocytes/ EpSCs of the outer root sheath of plucked anagenic hair follicles to demonstrate the effectiveness of this split-thickness skin autograft in promoting healing and complete closure of recalcitrant vascular leg ulcers [150].
Other clinical trials currently underway make use of other sources of stem cells, such as autologous lipoaspirate [151] and umbilical cord [152], to obtain MSCs respectively to treat diabetic ulcers and diabetic foot condition, reflecting the rapid emergent technology that desperately needs to move forward to meet patient needs in skin regeneration.
Concluding Remarks and Perspectives
Stem cells function as the human body's building blocks that have to make “choices.” Numerous studies have begun to disclose their inner unlimited capacities, and unlike what was previously thought, they can be driven toward transdifferentiation and give rise to unexpected relevant cell lineages. Furthermore, stem cells have unique features that translate into the release of important growth factors that play decisive roles in therapeutic regenerative medicine approaches
Specifically in skin regeneration, the use of stem cells can contribute in an active way to revolutionize and overcome some of the limitations of the current approaches. Different methods have been designed and made progress in this field (eg, in the healing of chronic wounds). Stem cells, as part of skin TE constructs, interact with the resident cells providing the cues for the regeneration of a functional tissue, instead of fibrotic scar tissue. The strategies are limitless but can contemplate distinct concerns like in vitro differentiation or in vivo paracrine signaling control in different skin wound models. Either way, communication with the resident cell or even by autocrine signaling seems to be a common feature that triggers the whole therapeutic potential and takes advantage of the irreplaceable plasticity of stem cells. Thus, cell-signaling cascades seem to be the key for transplanted stem cells to efficiently engraft and subsequently to differentiate.
Beyond all the excitement in the stem cell field, technical and scientific obstacles should not be ignored, and the awareness that there are issues still to be addressed is extremely important when moving to clinical applications. In skin regeneration itself, stem cell therapeutic potential is a work in progress, where details of stem cell biology should always be present, and where the potential of off-the-shelf stem cell-based tissue engineered products is only just beginning to be realized. The great outcome of innovative stem cell-based models is opening a new era of skin regeneration, where achieving functional skin is the main concern, including the building of yet lacking appendages, such as nerves, sweat glands, and blood supply.
Footnotes
Acknowledgments
The authors express their gratitude to Stephen Minger for the collaboration with former Stem Cell Biology Laboratory, King's College London, where experiments illustrated in Figs. 1 and ![]() were performed, and also to Tírcia Santos for the discussion of wound healing–related concepts.
were performed, and also to Tírcia Santos for the discussion of wound healing–related concepts.
Author Disclosure Statement
No competing financial interests exist.