
Abstract
Rapid liver regeneration is required after living-donor liver transplantation and oncologic liver resections to warrant sufficient liver function and prevent small-for-size syndrome. Recent evidence highlights the therapeutic potential of mesenchymal stem cells (MSC) for treatment of toxic liver injury, but whether MSC and their secreted factors stimulate liver regeneration after surgical injury remains unknown. Therefore, the aim of this study is to investigate the effect of human liver-derived MSC-secreted factors in an experimental liver resection model. C57BL/6 mice were subjected to a 70% partial hepatectomy and treated with either concentrated MSC-conditioned culture medium (MSC-CM) or vehicle control. Animals were analyzed for liver and body weight, hepatocyte proliferation, and hepatic gene expression. Effects of MSC-CM on gene expression in a human hepatocyte-like cell line (Huh7 cells) were analyzed using genome-wide gene expression arrays. Liver regeneration was significantly stimulated by MSC-CM as shown by an increase in liver to body weight ratio and hepatocyte proliferation. MSC-CM upregulated hepatic gene expression of cytokines and growth factors relevant for cell proliferation, angiogenesis, and anti-inflammatory responses. In vitro, treatment of Huh7 cells with MSC-CM significantly altered expression levels of ∼3,000 genes. Functional analysis revealed strong effects on networks associated with protein synthesis, cell survival, and cell proliferation. This study shows that treatment with MSC-derived factors can promote hepatocyte proliferation and regenerative responses in the early phase after surgical resection. MSC-CM may represent a feasible new strategy to promote liver regeneration in patients undergoing extensive liver resection or after transplantation of small liver grafts.
Introduction
O
Recent studies describe the potential role of stem cells, especially mesenchymal stem cells (MSC), to promote liver regeneration after toxic injury and protect against fulminant hepatic failure [10 –13]. MSC were initially thought only to reside in the bone marrow, providing a supportive niche for hematopoietic stem cells. More recently, MSC have been identified in multiple tissue compartments, including lung, liver, and adipose tissue, and they are described to provide pleiotropic effects in response to tissue injury [14 –16]. MSC have the ability to differentiate into hepatocytes and cholangiocytes and have immunomodulatory and anti-inflammatory properties [17 –21]. Further, they are described to promote angiogenesis by upregulating the expression of pro-angiogenic factors [22,23]. There is an ongoing discussion on whether MSC contribute to liver regeneration by (trans)differentiation into liver cells or by paracrine effects of their trophic factors. Research by van Poll and colleagues shows a stimulating effect of bone marrow-derived MSC (BM-MSC)-secreted factors on liver regeneration in the setting of toxic liver injury and hepatic failure [24,25]. In a clinical setting, application of MSC-derived factors to improve regeneration might be more beneficial than the use of MSC, since there is no risk of rejection or possible malignant transformation. In addition, the factors can be produced in large clinical grade quantities. However, whether MSC-secreted factors have beneficial effects on liver regeneration after surgical injury is unknown.
Recently we showed that the adult human liver harbors a population of MSC [15]. These liver-derived MSC (L-MSC) were found to mobilize from liver grafts at time of transplantation and could be retrieved from the organ preservation solution. L-MSC were found to be highly similar to BM-MSC with regard to adipogenic and osteogenic differentiation potential and Wnt-stimulated proliferative responses. Moreover, their genome-wide gene expression profile was highly similar to that of BM-MSC, with less than 0.3% of genes showing a 2- or more-fold difference in expression. Like BM-MSC, these L-MSC have immunosuppressive capacities and the potential to differentiate into hepatocyte-lineages in a mouse model of liver injury. Therefore, L-MSC from adult human liver have regenerative and immune regulatory potential. The aim of the current study is to investigate whether trophic factors secreted by L-MSC stimulate liver regeneration after surgical liver injury.
Materials and Methods
Animals
Male C57Bl/6 mice (age 13–18 weeks) were obtained from Charles River (Maastricht, The Netherlands) and maintained in the animal facility on a 12/12 h light/dark schedule. The animals had free access to food and drinking water. All animal experiments were performed with the approval of the institutional animal welfare committee.
Human L-MSC cultures and conditioned media
L-MSC were obtained from the organ preservation solution (Viaspan, Bristol-Myers Squibb, Woerden, The Netherlands), collected after cold storage of human liver grafts for transplantations performed at the Erasmus Medical Center, Rotterdam, The Netherlands. The Medical Ethical Council of the Erasmus Medical Center approved the use of human donor material for medical research. Mononuclear cells were isolated from the collected preservation fluids by density gradient centrifugation using Ficoll-Paque Plus (GE Healthcare, Uppsala, Sweden) and put into culture as previously described [15]. Culture media consisted of DMEM (Lonza, Verviers, Belgium) supplemented with 10% fetal bovine serum (Sigma-Aldrich, Taufkirchen, Germany), L-Glutamin (Lonza), penicillin, and streptomycin (Gibco, Invitrogen, Paisley, United Kingdom). When L-MSC were cultured under serum-free conditions, culture media consisted of MEM-alpha (Gibco) supplemented with 0.05% bovine serum albumin (Sigma-Aldrich), L-Glutamin, penicillin, and streptomycin. The conditioned culture media were collected 3 days after medium change of L-MSC cultures from passage 6–10. MSC-conditioned culture medium (MSC-CM) was concentrated ∼25-fold by filtration with 3-kD molecular cutoff filters (Amicon Ultra, Millipore, Carrigtwohill, Ireland).
Partial hepatectomy and MSC-CM treatment
Liver regeneration was induced by subjecting C57BL/6 mice to a 70% partial hepatectomy (PH) as first described by Higgins and Anderson in 1931. Animals were anesthetized with isoflurane and, after a midline laparotomy, the left lateral and median lobes of the liver were ligated and resected. The peritoneum and skin were sutured separately. All procedures were performed under clean conditions. Animals were treated intraperitoneally with 100 μL concentrated MSC-CM in phosphate buffered saline (PBS) (Lonza; total volume 0.5 mL) or with PBS alone as vehicle control, starting 4 h prior to or at time of PH, and in both cases 24 h after resection.
In addition, a serum-free experiment was performed in which animals were injected at the time of and 24 h after PH with the concentrated MSC-CM collected from serum-free cultured L-MSC, to exclude possible effects on liver regeneration caused by fetal bovine serum. The control group in this serum-free experiment was treated with concentrated culture medium treated exactly the same as the serum-free MSC-CM (SF-MSC-CM), but without the presence of L-MSC [serum-free unconditioned medium (SF-UM)]. All animals were sacrificed 2 days after PH and their livers and blood were collected to further investigate the effects of MSC-CM on hepatocyte proliferation, hepatic gene expression, and serum markers of liver function and injury in the early phase of liver regeneration.
Weight calculations
The animals were weighed daily prior to treatment and the resected liver mass was weighed after PH. The initial total liver weight was calculated as follows:
resected liver weight/70*100 (g)
At time of sacrifice, animals and their regenerated liver mass were weighed and the percentage of reconstitution of the liver was calculated by:
regenerated liver weight/initial total liver weight*100 (%)
The liver to body weight ratio was calculated by:
regenerated liver weight/body weight at time of harvest*100 (%)
Immunohistochemistry
One hour prior to sacrifice, animals were injected intraperitoneally with 50 mg/kg 5-Bromo-2′-deoxyuridine (BrdU; B5002, Sigma-Aldrich). Livers were harvested and processed to 4 μm thick formalin fixed, paraffin-embedded sections. Antigen retrieval was achieved by boiling the slides in 0.01 M sodium citrate; pH 6.0 (microwave 1,000 Watt; 1×7 and 2×3 min). Endogenous peroxidase was blocked by 0.6% H2O2 in PBS for 30 min at room temperature, after which DNA was denaturated by incubation for 1 h at 37°C in 0.1 M HCl in aqua dest. Aspecific binding was prevented by 0.5% milk powder supplemented with 0.15% glycin in PBS (blocking buffer). Slides were incubated overnight at 4°C with monoclonal mouse anti-BrdU (Bu20a; DakoCytomation, Glostrup, Denmark; 1:80 in blocking buffer). The next day slides were incubated for 30 min at room temperature with polyclonal rabbit anti-mouse IgG/HRP (P0161; DakoCytomation; 1:1,000 in blocking buffer). After antibody incubation slides were incubated with DAB-solution and counterstained with hematoxylin. Per animal 4 high power fields (HPF; 400×) were analyzed for BrdU positive hepatocytes.
Serum analysis of enzyme levels
Blood samples were collected at time of sacrifice in heparin-coated micro tubes. After collection, samples were centrifuged (19 min, 1,800 rpm) to separate the serum, which was then further analyzed at the clinical chemical core facility of the Erasmus MC-University Medical Center to determine albumin, bilirubin, alanine transaminase (ALT), and aspartate transaminase (AST) levels.
Real-time quantitative reverse transcription-polymerase chain reaction
At time of sacrifice, liver tissue was stored overnight at 4°C and thereafter at −80°C in Allprotect Tissue Reagent (Qiagen, Hilden, Germany) for RNA preservation. Total RNA was extracted using Trizol (Qiagen) and chloroform after mechanical disruption of the tissue. RNA was precipitated in 75% ethanol and dissolved in RNase-free water. RNA quantity and quality was analyzed using a NanoDrop ND-1000 (Thermo Fisher Scientific, Wilmington, DE). One microgram of RNA was reverse-transcribed to cDNA using an iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA). Polymerase chain reaction (PCR) primers (presented in Table 1) were synthesized by Isogen Life Science (Maarssen, The Netherlands) and Biolegio (Nijmegen, The Netherlands). Real-time quantitative reverse transcription-polymerase chain reaction (RT-PCR) was performed with a SensiMix SYBR and Fluorescein Kit (Bioline, London, United Kingdom) and MyIQ real-time PCR detection system (Bio-Rad Laboratories) according to the manufacturer's instruction.
Genome-wide gene expression profiling and proliferation of Huh7 cells cultured with MSC-CM
For genome-wide gene expression profiling, Huh7 cells were cultured for 48 h with MSC-CM or unconditioned medium (final concentration 50%; 6-well plate; 3 wells/condition; 2.5×105 cells/well). Total RNA from these cultures were isolated using the RNeasy Micro Kit (Qiagen) and used for gene expression analysis using Human Gene 1.0 ST Genechips (Affymetrix, Santa Clara, CA) according to the manufacturer's instructions.
The effect on proliferation of Huh7 cells was determined using an MTT assay, for which Huh7 cells were cultured up to 4 days with MSC-CM or UM (final concentration 50%; 96-well plate; 3 wells/time point/condition; 2×104 cells/well). At day 1, 2, 3, and 4 after starting treatment, the number of metabolically active cells was quantified by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, 0.5 mg/mL) colorimetry.
Statistical analysis
For genome-wide gene expression analysis, normalized signals were generated using RMAExpress. Class predictions were performed with Partek Genomics Suite software and functional analyses were performed using Ingenuity Pathway Analysis (Ingenuity Systems, Redwood City, CA). With these array analyses, P values<0.005 using a 2-way analysis of variance test were considered significant. All other data are presented as mean±standard error of the mean (SEM) and statistical analyses were performed using the Mann–Whitney test with GraphPad Prism software and P≤0.05 was considered statistically significant.
Results
MSC-CM treatment enhances hepatocyte proliferation and liver weight gain after PH
All treatment groups showed 100% survival. Hepatocyte proliferation, quantified by the percentage of BrdU positive hepatocyte nuclei, was significantly increased at day 2 after PH in animals treated at time of PH with MSC-CM compared to animals treated at time of PH with PBS (20.0% vs. 12.1%, P=0.02; Fig. 1A, B). Animals pretreated 4 h prior to PH with MSC-CM showed a similar trend compared to animals pretreated with PBS (19.3% vs. 14.7%; data not shown), however this difference did not reach statistical significance. Interestingly, the liver to body weight ratio was significantly increased in animals pretreated with MSC-CM compared to animals pretreated with PBS (3.1% vs. 2.8% of total body weight, P=0.05; data not shown). No significant differences were seen between animals treated at time of PH with MSC-CM or PBS.
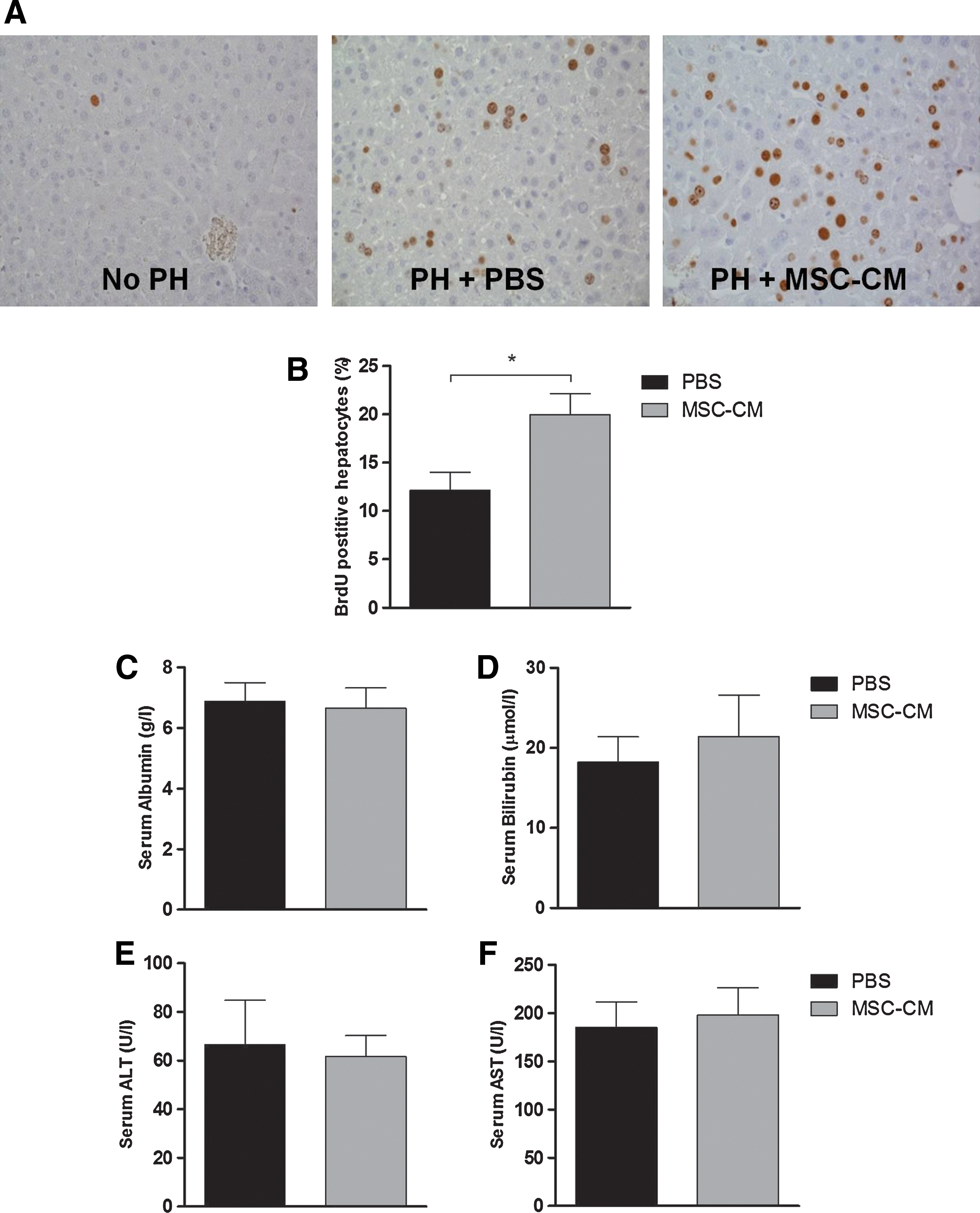
Effects of MSC-CM on hepatocyte proliferation, function, and injury.
Even though proliferation was stimulated, known to cause a shift away from metabolic functions, MSC-CM treatment did not cause changes in liver function compared to PBS treatment, as shown by serum albumin and bilirubin levels (Fig. 1C, D). Neither did MSC-CM treatment induce or reduce hepatocyte cell injury after PH compared to PBS treatment, as shown by ALT and AST levels (Fig. 1E, F).
MSC-CM induces expression of genes relevant for hepatocyte proliferation
Previous studies have shown that tumor necrosis factor-alpha (TNF-α) and interleukin 6 (IL-6) are crucial priming factors for hepatocytes to enter the cell cycle, whereas hepatocyte growth factor (HGF) is important in the proliferative phase [26 –28]. Downstream in their signal transduction cascades, the passage of hepatocytes from the G1 into the S phase is associated with upregulation of several cyclins including Cyclin D [29]. Transforming growth factor-beta (TGF-β) is known to be involved in the negative feedback on hepatocyte proliferation [30]. Figure 2A shows the hepatic gene expression levels of these factors and of proliferating cell nuclear antigen (PCNA), known to be upregulated during DNA synthesis. MSC-CM treatment at time of PH significantly upregulated hepatic gene expression levels of TNF-α (2.6-fold, P=0.02), HGF (1.9-fold, P=0.03), PCNA (4.8-fold, P<0.01), and TGF-β (2.1-fold, P=0.02) compared to PBS treatment. IL-6 and Cyclin D1 gene expression did not show a statistically significant upregulation.
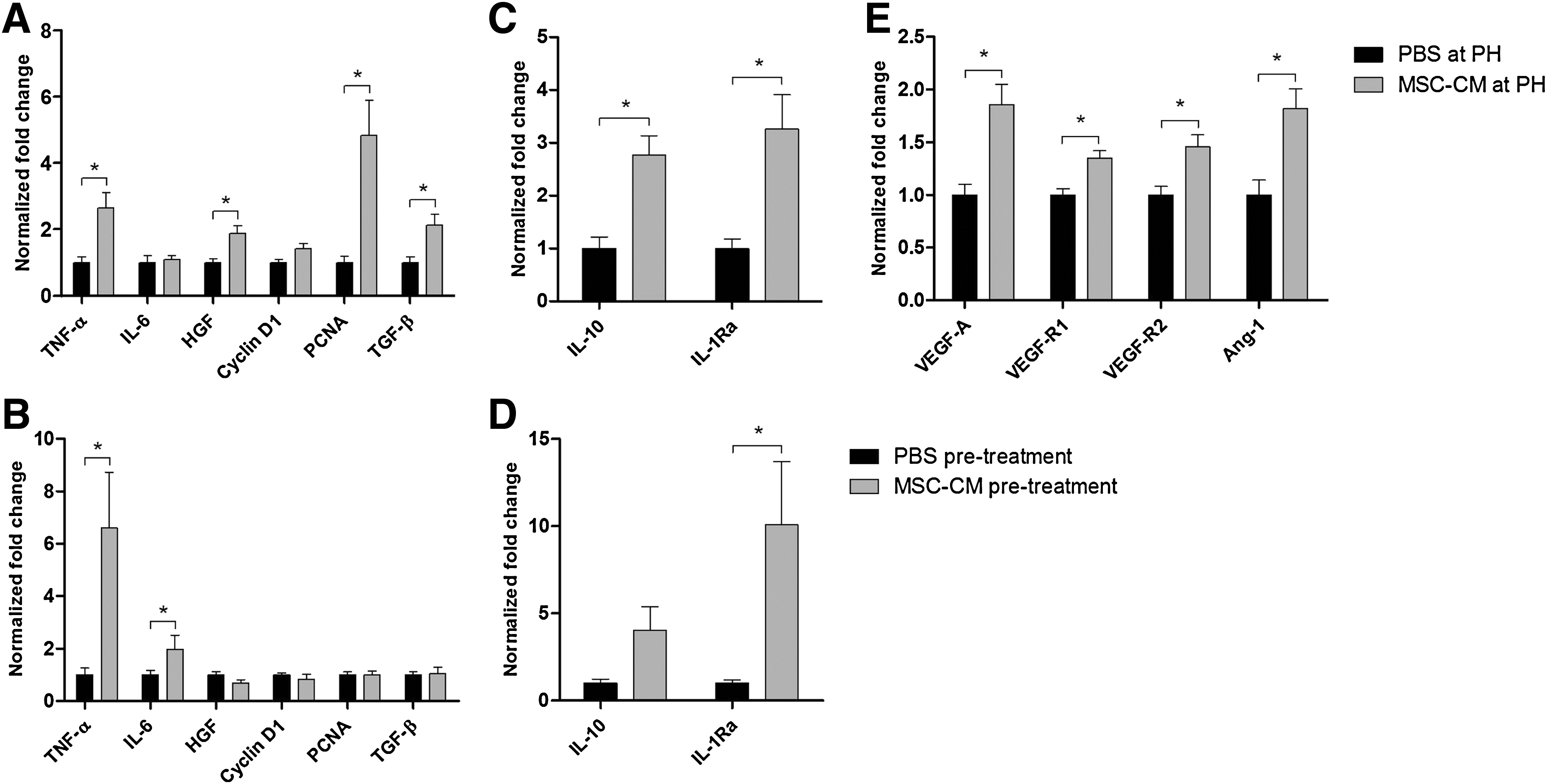
Effects of MSC-CM on hepatic gene expression. Hepatic gene expression levels were determined by quantitative RT-polymerase chain reaction and normalized against β-actin.
Pretreatment with MSC-CM, compared to pretreatment with PBS, only resulted in significant upregulation of TGF-β gene expression (2.6-fold, P<0.01; data not shown). A possible explanation for these findings could be that pretreatment with MSC-CM might accelerate the regenerative response of the liver after surgical injury, with a shift forward in induction of abovementioned genes. Therefore, effects of pretreatment with MSC-CM on the hepatic gene expression at time of resection were evaluated. As shown in Fig. 2B, pretreatment with MSC-CM had no effect on HGF, Cyclin D1, PCNA, or TGF-β gene expression, but caused a significant upregulation of TNF-α (6.6-fold, P=0.02) and IL-6 gene expression (2.0-fold, P=0.05) at time of resection.
MSC-CM treatment induces an increased anti-inflammatory response
MSC are described to have anti-inflammatory capacities with an important role for interleukin 10 (IL-10) and interleukin 1 receptor antagonist (IL-1Ra) [21,31]. Therefore, effects on the anti-inflammatory response after PH were evaluated by analyzing gene expression levels of these cytokines. MSC-CM treatment at time of PH induced a significant upregulation of both IL-10 (2.8- fold, P=0.05) and IL-1Ra gene expression (3.3-fold, P=0.02) compared to PBS treatment (Fig. 2C). Pretreatment with MSC-CM, compared to PBS pretreatment, also showed a significant upregulation of IL-1Ra gene expression at day 2 after PH (3.5-fold, P=0.05), but not of IL-10 (data not shown). Upregulation of IL-1Ra gene expression was already observed at time of resection, 4 h after pretreatment with MSC-CM (10.1-fold, P=0.02; Fig. 2D).
Treatment with MSC-CM upregulates gene expression of pro-angiogenic factors
Besides hepatocyte proliferation, regeneration of damaged or lost vasculature plays an important role in liver regeneration. We therefore investigated the effects on hepatic gene expression of the pro-angiogenic factors vascular endothelial growth factor A (VEGF-A), vascular endothelial growth factor receptor 1 and 2 (VEGF-R1 and -R2), and angiopoietin 1 (Ang-1). Figure 2E shows that in animals treated with MSC-CM at time of PH, a significant upregulation in hepatic gene expression of the pro-angiogenic factors VEGF-A (1.9-fold, P<0.01), VEGF-R1 (1.4-fold, P<0.01), VEGF-R2 (1.5-fold, P<0.01), and Ang-1 (1.8-fold, P=0.02) was seen at day 2 after PH compared to PBS-treated animals. Pretreatment with MSC-CM, compared to PBS pretreatment, showed a similar induction in gene expression of these factors (VEGF-A 1.4-fold, VEGF-R2 1.9-fold, and Ang-1 1.5-fold), but differences were only statistically significant for VEGF-R1 (1.4-fold, P=0.03; data not shown). Pretreatment with MSC-CM did not cause significant changes in angiogenic gene expression levels at time of resection (data not shown).
Treatment of Huh7 cells with MSC-CM induces gene pathways involved in liver regeneration
To validate our findings in an in vitro model, a hepatocyte-like cell line (Huh7 cells) was stimulated with MSC-CM. MSC-CM treatment up to 4 days had no significant effect on cell viability, though there was a slight trend toward increased cell numbers, as determined by an MTT-assay (data not shown). Genome-wide gene expression analysis showed that in Huh7 cells exposed to MSC-CM ∼3,000 genes (<10% of the genome) were significantly differentially expressed compared to Huh7 cells exposed to UM. Functional analysis revealed that these genes are associated with pathways and networks relevant for protein synthesis, cell survival, and cell proliferation (Fig. 3).

Molecular and cellular functions affected in Huh7 cells cultured with MSC-CM. Genome-wide gene expression profiles in Huh7 hepatoma cells after 48 h culturing with MSC-CM were analyzed using Affymetrix genechips. Functional analysis using Ingenuity Pathway Analysis of the ∼3,000 significantly differentially expressed genes (P<0.005) after MSC-CM treatment revealed their involvement in pathways and networks relevant for cell growth, proliferation, and survival.
SF-MSC-CM causes similar effects on liver regeneration as serum-containing MSC-CM
The use of bovine serum in the culture media of MSC will not be suitable for clinical applications of MSC-CM in humans, because of concerns regarding prions, zoonosis, and xenogenic immune responses. Therefore, the effects of concentrated SF-MSC-CM were investigated in liver regeneration after PH and compared to the effects of concentrated SF-UM. Similar to treatment at time of PH with serum-containing MSC-CM, no significant changes in body weight, liver to body weight ratio, or serum levels of albumin, bilirubin, ALT, and AST were seen in animals treated with SF-MSC-CM compared to animals treated with SF-UM (data not shown). SF-MSC-CM treatment caused a significant stimulation of hepatocyte proliferation at day 2 after PH compared to SF-UM treatment (17.3% vs. 9.1%, P<0.01; Fig. 4), which was similar to the effect of serum-containing MSC-CM. These results show that MSC-CM can be produced under conditions suitable for clinical application in humans and furthermore that positive effects of MSC-CM on liver regeneration are not caused by factors present in serum.

Effects of SF-MSC-CM on hepatocyte proliferation. Two days after PH, livers were processed for immunohistochemistry on BrdU to quantify hepatocyte proliferation after SF-MSC-CM and SF-UM treatment. n=7 in the SF-MSC-CM group and n=5 in the SF-UM group; *P≤0.05 versus SF-UM. SF-MSC-CM, serum-free MSC-CM; SF-UM, serum-free unconditioned medium.
Discussion
In this study we found that treatment with the concentrated conditioned culture medium of human L-MSC stimulates the already robust liver regeneration in mice after 70% PH. Hepatocyte proliferation was increased by 65% and the tightly regulated liver to body weight ratio by 11%. These effects are in line with findings of van Poll et al., who observed a 3-fold increase in the number of proliferating hepatocytes in animals treated with MSC-CM from BM-MSC after toxic liver injury [24].
Possible underlying molecular mechanisms responsible for the stimulating effects of MSC-CM could be altered expression levels of cytokines and growth factors involved in liver regeneration. Indeed, MSC-CM upregulated gene expression levels of the cytokine TNF-α and growth factor HGF, known to be required for initiation of and progression through the hepatocyte cell cycle [1,26,28]. MSC-CM furthermore upregulated gene expression of the pro-angiogenic factors VEGF-A, VEGF-R1 and -R2 and Ang-1, suggesting that MSC-CM promotes neo-angiogenesis after liver resection. This is in line with findings of Lai and co-workers, who found increased capillary density and better heart function after MSC-CM administration to pigs with a myocardial infarction [32].
Another interesting effect is the induction of an anti-inflammatory response, as shown by upregulated gene expression of the cytokines IL-10 and IL-1Ra. These findings are in line with previous reports suggesting the immunomodulatory effect of MSC can be attributed to their stimulating effects on IL-10 expression and their production of IL-1Ra [21,31,33,34]. The anti-inflammatory response may prevent adverse effects of the pro-inflammatory cytokine TNF-α, which is needed for the initiation of liver regeneration, but also mediates cell death [35]. This is supported by the findings of Cosgrove et al., showing that IL-1Ra enhances TNF-α-stimulated hepatocyte proliferation [36].
Overall, our results show that MSC-CM stimulates liver regeneration by influencing signaling pathways relevant for hepatocyte proliferation, angiogenesis and reduction of inflammation. This is supported by mass spectrometry analysis of the MSC-CM, which revealed the presence of IL-6 and TGF-β, among other extracellular and cytoplasmic components known to be related to cellular growth, proliferation and cellular homeostasis (see Supplementary Methods and Supplementary Table S1; Supplementary Data are available online at
The most critical phase in patients undergoing partial liver resection or transplantation of a partial graft is during the early postoperative phase. Potential new treatment strategies to enhance or accelerate liver regeneration should therefore act in this early postoperative period. For that reason, we chose to investigate the effects of MSC-CM at day 2 after PH, known to show the early peak in hepatocyte proliferation in mice. Furthermore, both treatment with MSC-CM at time of PH as well as pretreatment 4 h prior to resection were investigated. Based on hepatocyte proliferation and gene expression data, treatment at time of PH seemed most effective in stimulating liver regeneration. Interestingly however, we found a significant increase in liver to body weight ratio after pretreatment, whereas no significant difference was found if animals were treated at time of resection. We hypothesize that MSC-CM pretreatment shifts the regenerative response of the liver after surgical injury forward and thus accelerates liver regeneration. Gene expression data collected at day 2 after PH in both treatment groups, combined with data collected at time of PH from pretreated animals, gave insight in the (timely) underlying mechanism by which MSC-CM seems to stimulate liver regeneration. The effects of MSC-CM pretreatment on gene expression in the resected liver tissue indeed showed a significant increase in TNF-α, IL-6 and IL-1Ra expression at time of PH. Although further evaluation is required, these data support the possibility that pretreatment with MSC-CM is able to prime the liver prior to surgery and accelerate regeneration after liver resection. This is also supported by the finding that 2 days after PH only gene expression of TGF-β, known to be important in the termination phase of liver regeneration, was significantly upregulated in animals pretreated with MSC-CM. This in contrast to significantly upregulated gene expression levels of TNF-α, HGF and PCNA, relevant in earlier stages of hepatocyte proliferation, found in animals first treated at time of PH.
The abovementioned hypothesis that pretreatment with MSC-CM primes the liver prior to surgery and thereby accelerates the regenerative response, seems the most likely explanation for the different results in liver weight gain and hepatocyte proliferation between animals pretreated and treated at time of PH with MSC-CM. However, a possible other underlying mechanism could be that pretreatment stimulates the early recruitment or activation of stem/progenitor cells, which in turn aid to the regeneration process.
Aside from hepatocyte proliferation and liver weight gain, the functional and metabolic recovery of the liver is an essential component of liver regeneration after injury. At any given time, liver function reflects a complex balance between cellular proliferation and metabolic homeostasis [37 –39]. During the early phase of liver regeneration, a large part of the liver's energy is needed for rapid proliferation of parenchymal and nonparenchymal cells, thereby decreasing the amount of energy available for metabolic functions. Because of the increase in the percentage of proliferating cells, one could therefore expect a decrease in metabolic activity in MSC-CM-treated animals compared to control animals. However, in our experiments, no decrease in serum albumin or increase in bilirubin was found. We can therefore conclude that MSC-CM increases liver regeneration without negatively affecting metabolic homeostasis.
In this study we deliberately chose to use stem cell-conditioned media, rather than a cell transplantation strategy. Compared with MSC-CM, cell transplantation has the potential advantage of regeneration via (trans)differentiation or cell fusion and possible prolonged effects on the micro-environment by long-term engrafted MSC. However, although differentiation of MSC into hepatocyte-like cells has been described, differentiation into fully functional hepatocytes is rare. Moreover, engraftment levels of MSC are often insignificant and cell transplantation may lead to sensitization [40]. Although our findings show the effectiveness of MSC-CM in stimulating liver regeneration, concerns regarding the safe application in humans remain. Especially the use of (bovine) serum in the culture media of MSC raises concerns about transmission of prions, zoonosis, and xenogenic immune responses. However, our experiments with MSC-CM from serum-free cultured L-MSC showed similar effects on liver regeneration as MSC-CM from serum-containing cultures. This illustrates that MSC-CM can be produced under conditions suitable for safe application in humans and furthermore that stimulation of liver regeneration by MSC-CM is not caused by factors present in serum.
In summary, MSC-secreted factors are effective in stimulating liver regeneration after surgical resection by influencing expression levels of cytokines and growth factors relevant for cell proliferation, angiogenesis, and anti-inflammatory responses. Therefore, MSC-secreted factors represent a promising therapeutic strategy to improve liver regeneration in patients with a small-for-size liver graft or after extensive liver resections.
Footnotes
Acknowledgments
The authors would like to thank Dr. Wendy van Veelen, Jasper Deuring, and Werner Helvensteijn for technical support; Dr. Yuedan Li and Dr. Martijn Brugman for help with gene array data analyses and Dr. Jeroen Demmers for helping with mass spectrometry (Erasmus MC Proteomics Center). This study has been financially supported by the Astellas Trans(p)la(n)t(at)ional Research Prize and the Liver Research Foundation (SLO) Rotterdam.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.