
Abstract
CD41 (αIIb integrin), a specific marker for megakaryocytes and platelets, was recently shown to mark the initiation of definitive hematopoiesis in mouse embryos. However, whether embryonic CD41+ populations have a nonhematopoietic potential remains elusive. Here, we report that the CD41+ cells from the mouse E11.0 aorta-gonad-mesonephros (AGM) region and yolk sac (YS) expressed a set of mesenchymal markers (as revealed by reverse transcriptase–polymerase chain reaction), displayed myofibroblastic/fibroblastic potential in vitro under mesenchymal culture conditions, and differentiated into α-SMA+/epimorphin+/vimentin+ cells in the lungs of adult recipients after systemic transplantation. This unique cell population with fibroblastic potential expressed intermediate rather than high levels of CD41 and was negative for CD34 in the AGM region. In contrast, circulating CD41+ cells in the embryonic blood stream harbored no similar fibroblastic potential. Compared with the YS, the AGM-derived CD41+ cells had a more robust fibroblastic potential, as revealed by higher in vitro growth rates. Interestingly, the AGM-derived CD41+ cells demonstrated a stronger response to the chemotaxin of circulating blood plasma than the YS-derived CD41+ cells. We are the first group that illustrates the fibroblastic potential of an embryonic CD41+ population in vitro and in vivo, reflecting the close association between blood and mesenchyme development during mouse mid-gestation. The precise origin of these mesenchymal populations needs further clarification.
Introduction
E
A conventional marker for migrating cell populations in embryonic circulation is very important, especially for definitive hematopoietic cells. CD45 is a well-known pan-hematopoiesis marker in adult BM; however, it is not a good marker for early embryonic hematopoiesis. For instance, the high proliferative potential precursors with hemangioblastic activity reside within the CD45-negative subset of the E11.5 AGM region [3]. Runx1 is particularly useful in recognizing embryonic hematopoietic precursors in mice [4]; however, it cannot be routinely used for fractionating wild-type cells. As one of the target genes of Runx1, CD41 (also known as αIIb integrin) [2], a well-known specific marker for megakaryocytes and platelets [5], has recently been found to mark the initiation of definitive hematopoiesis in developing embryos and differentiation cultures of embryonic stem cells (ESCs) [2,6 –9]. By the fetal liver stage, the expression of CD41 is reduced, while CD45 is prevalent in all the definitive hematopoietic cells [2], suggesting a differentiation into CD45+ hematopoietic lineage-restricted cells from CD41+ hematopoietic precursors in the fetal liver.
The functional role of CD41 on the surface of hematopoietic precursor cells remains largely unknown, as CD41 knockout mice display defects only in platelet function [10]. On the other hand, CD41+ precursor cells can be detected in the YS, AGM region, and placenta, suggesting no special correlation with embryonic hematopoietic origin. Researchers usually focus on the CD41+/c-kit+ definitive hematopoietic progenitors in mice [2] or the CD41+/c-myb+ nascent HSCs in zebrafish [11]; however, little is known about the biological properties of these cells, particularly the heterogeneity in the differentiation potential of the CD41+ population in mouse embryos.
Here, we report that CD41+ cells from the mouse E11.0 AGM region and YS expressed a set of mesenchymal markers that could be detected by reverse transcriptase–polymerase chain reaction (RT-PCR), displayed fibroblastic potential in vitro, and differentiated into α-SMA+/epimorphin+/vimentin+ cells in vivo. In contrast, the circulating CD41+ cells in the embryonic blood stream displayed no similar fibroblastic potential.
Materials and Methods
Embryo dissections and preparation of single-cell suspensions
Adult C57BL/6 mice were purchased from the China Academy of Medical Sciences Animal Center, and transgenic GFP breeding pairs (C57BL/6 background) were a kind gift from Professor Ming Fan (Institute of Basic Medical Sciences, Beijing, China). C57BL/6 mice were mated overnight. Noon of the day, which was a vaginal plug, was detected and defined as E0.5. Embryos at various stages of gestation were obtained from time-mated pregnant mice. All animal care and experimental procedures were performed in accordance with the Institute of Military Medical Sciences Guide for Laboratory Animals (Beijing, China).
The embryos were carefully dissected from pregnant mice in cold α-minimal essential medium (α-MEM; GIBCO) containing 10% fetal bovine serum (FBS; HyClone). For collection of the circulating blood (CB) plasma, α-MEM without FBS was used. Embryo CB was collected within 2–3 min after separation of the YS as previously described [12]. Special care was taken to remove the umbilical and vitelline arteries from preparations of the YS and embryonic CB. The AGM region and YS were dissected and then digested with 5 mL of 0.1% collagenase NB 4 (SERVA Electrophoresis) plus 20% FBS in phosphate-buffered saline (PBS) for 60–90 min at 37°C with gentle shaking at 15 min intervals to dissociate into single-cell suspensions. The cells were washed twice with cold PBS containing 10% FBS, and cell clumps were removed by passing the cells through a 70 μm nylon mesh (BD Biosciences, Discovery Labware) before being sorted.
Fluorescence-activated cell sorting
For the separation of single-cell suspensions of total CD41+ cell populations, subfractioning of the CD41 cell populations based on the CD41 expression levels or combined with CD34 from the AGM region, YS, and CB, the cells were washed twice with PBS containing 0.1% bovine serum albumin (BSA) and then incubated with PE-conjugated anti-mouse CD41 (BD Bioscience Pharmingen) or anti-CD41 combined with FITC-conjugated anti-mouse CD34 (eBioscience) for 30 min at 4°C. PE-conjugated isotype (BD Pharmingen) or FITC-conjugated isotype (eBioscience) were used as controls. After incubation, the cells were washed twice and then sorted on a MoFlow cytometer (Cytomation) or FACSAria II Flow Cytometer (BD Biosciences). The freshly isolated cells were cultured or assayed by nested RT-PCR, Giemsa staining, transwell migration, and transplantation.
CD41+ cell culture and induction
In vitro mesenchymal differentiation
For mesenchymal differentiation, the sorted CD41+ cells or the subfractioning cells (1×105 cells per cm2) were cultured in α-MEM medium (10% FBS, 10 ng/mL basic fibroblast growth factor [bFGF], and 0.1% penicillin/streptomycin). The medium was replaced 3 days after plating and was then replaced every other day. The cells were passaged by 0.25% trypsin when they reached 80%–90% confluence.
Hematopoietic CFU-C assay
A hematopoietic colony-forming unit-culture (CFU-C) assay was performed as previously described [3]. Briefly, the hematopoietic semisolid medium was composed of 0.9% methylcellulose (Sigma), 2 mM penicillin/streptomycin (GIBCO), 2 mM glutamine (GIBCO), 1% BSA (GIBCO), 5% protein-free hybridoma medium II (GIBCO), 200 μg/mL iron-saturated transferrin (Sigma-Aldrich Corp.), 0.45 mM monothioglycerol (Sigma), 15% FBS (GIBCO), and a cytokine cocktail, including rmSCF (50 ng/mL; PeproTech), rmIL-3 (10 ng/mL; PeproTech), rhIL-11 (10 ng/mL; R&D), rmGM-CSF (10 ng/mL; PeproTech), rhTpo (20 ng/mL; R&D), and rhEpo (3 U/mL; Kirin). After 7 days of inoculation, colony-forming unit-erythroid (CFU-E), burst-forming unit-erythroid (BFU-E), colony-forming unit-Granulocyte/Megakaryocyte/Granulocyte and Monocyte (CFU-G/M/GM), and CFU-Mix were counted.
Nested RT-PCR
Nested RT-PCR for freshly isolated CD41+ cells was performed as previously described [3]. The specific primers used for the nested RT-PCR are listed in Table 1. The RT-PCR products were separated in a 1% agarose gel and visualized by staining with GelGreen™ dye (Biotium).
α-SMA, alpha-smooth muscle actin; MEF-2C, myocyte enhancer factor 2C; SPP-1, secreted phosphoprotein 1 (also named osteopontin); Flk-1, vascular endothelial growth factor receptor (Flk-1/KDR).
Immunofluorescence staining
Immunofluorescence staining was performed as previously described [13]. Briefly, for double immunofluorescence staining of cultured CD41+ cells or the sub-fractioning cells on 96-well plates, permeabilized cells were incubated with mouse antialpha-smooth muscle actin (anti-α-SMA, 1:800; Sigma) and rabbit anti-vimentin (1:100; Cell Signaling Technology, Inc.). FITC-conjugated goat anti-mouse IgG and TRITC-conjugated goat anti-rabbit IgG were used as secondary antibodies (1:100; Jackson ImmunoResearch Laboratories).
For frozen sections of mouse AGM and YS tissues, biotin-conjugated rat anti-mouse CD41 (1:200; BD Bioscience Pharmingen) and anti-α-SMA (1:800; Sigma) were co-stained, followed by a combination of PE-Streptavidin (eBioscience) and FITC-goat anti-mouse IgG (Jackson ImmunoResearch Laboratories). For frozen sections of mouse lungs in transplantation, rabbit anti-GFP (Proteintech Group) and rat anti-mouse CD31 (BD Bioscience Pharmingen), mouse anti-α-SMA (Sigma-Aldrich Corp.), mouse anti-vimentin (Sigma), and mouse anti-epimorphin (a gift from Dr. Yohei Hirai) were used. Confocal images were collected by an LSM 510 META confocal system (Zeiss; Carl Zeiss MicroImaging). All the nuclei were visualized by 2 μM 4′,6-diamidino-2-phenylindole (DAPI; Sigma).
Transplantation assay
Six-week-old adult C57BL/6 mice were given a single 6-Gy dose of whole-body irradiation from a 60Co source as previously described [14]. Freshly sorted GFP+CD41+ cells from E11.0 AGM, YS, and CB tissues of GFP-transgenic C57BL/6 mice were intravenously injected into the tail vein of irradiated wild-type C57BL/6 mice. Each group of recipient mice (AGM, YS and CB) received 6×104 GFP+CD41+ cells along with 2×105 GFP−CD41− partner cells sorted from non-GFP offspring derived from pregnant GFP-transgenic C57BL/6 mice. At week 4, the harvested tissues, including the heart, liver, spleen, lung, and kidney, were cut into 6 μm frozen sections using a HM 505E cryostat (Microm, part of Thermo Fisher), fixed with cold acetone for 10 min, and screened for GFP+ cells under the fluorescence microscope.
Transwell migration assay
Cell migration was assessed using Falcon HTS FluoroBlok 24-well inserts (BD Biosciences) with 5-μm pores. After sorting, AGM-derived CD41+ and YS-derived CD41+ cells (1–2×105) were immediately placed on each insert in an upper chamber in 250 μL of α-MEM. A 500 μL volume of α-MEM or the same amount of collected CB plasma was placed in the bottom chamber. Twelve hours later, the migrated cells in the bottom chamber were fixed with 4% paraformaldehyde, and carefully stained by 2 μM DAPI. The number of migrated CD41+ cells was determined by averaging the numbers in ten random fields for each group.
Phase-contrast and fluorescence imaging
Brightfield images were collected using an ECLIPSE TE2000-U microscope (Nikon Corporation Precision Equipment Company). Images were acquired using a digital camera (Cool SNAP 5.0; Roper Scientific). Fluorescence images were analyzed by inverted light microscopy (IX70; Olympus Optical Co.). Confocal images were collected on an LSM 510 META confocal system (Zeiss; Carl Zeiss MicroImaging).
Statistical analysis
The results were expressed as the means±standard deviation of at least 33 experiments for CD41 fluorescence-activated cell sorting (FACS) and at least 4 experiments for the migration assay. The significance of the migration assay was determined using Student's t- test: *P<0.01, compared with the control data.
Results
Since CD41 has been reported to be a definitive hematopoietic cell marker in mouse embryos, the hematopoietic potential of CD41+ cells has been extensively investigated [2,6 –8]. In addition, some reports have suggested an absence of endothelial potential in CD41+ cells by in vitro tube-forming assays on OP9 stromal layers [2,15]. Here, we have focused on the mesenchymal potential and migration ability of CD41+ cells during embryogenesis.
CD41 expression analysis of the AGM, YS, and CB at E11.0
To date, two methods (ie, fluorescence-activated cell sorting [FACS] and magnetic cell separation [MACS]) have been typically utilized to isolate hematopoietic cells with specific markers. To eliminate the possibility of cross-contamination between the CD41+ and CD41− populations [6] throughout the study, all the CD41+ cells were sorted by FACS, a relatively more stringent method than MACS. Fig. 1A illustrated 1 typical FACS experiment of at least 33 independent experiments. The percentages of CD41+ cells were 4.94±1.46 (AGM), 8.52±1.27 (YS), and 2.37±0.60 (CB) (Fig. 1B).
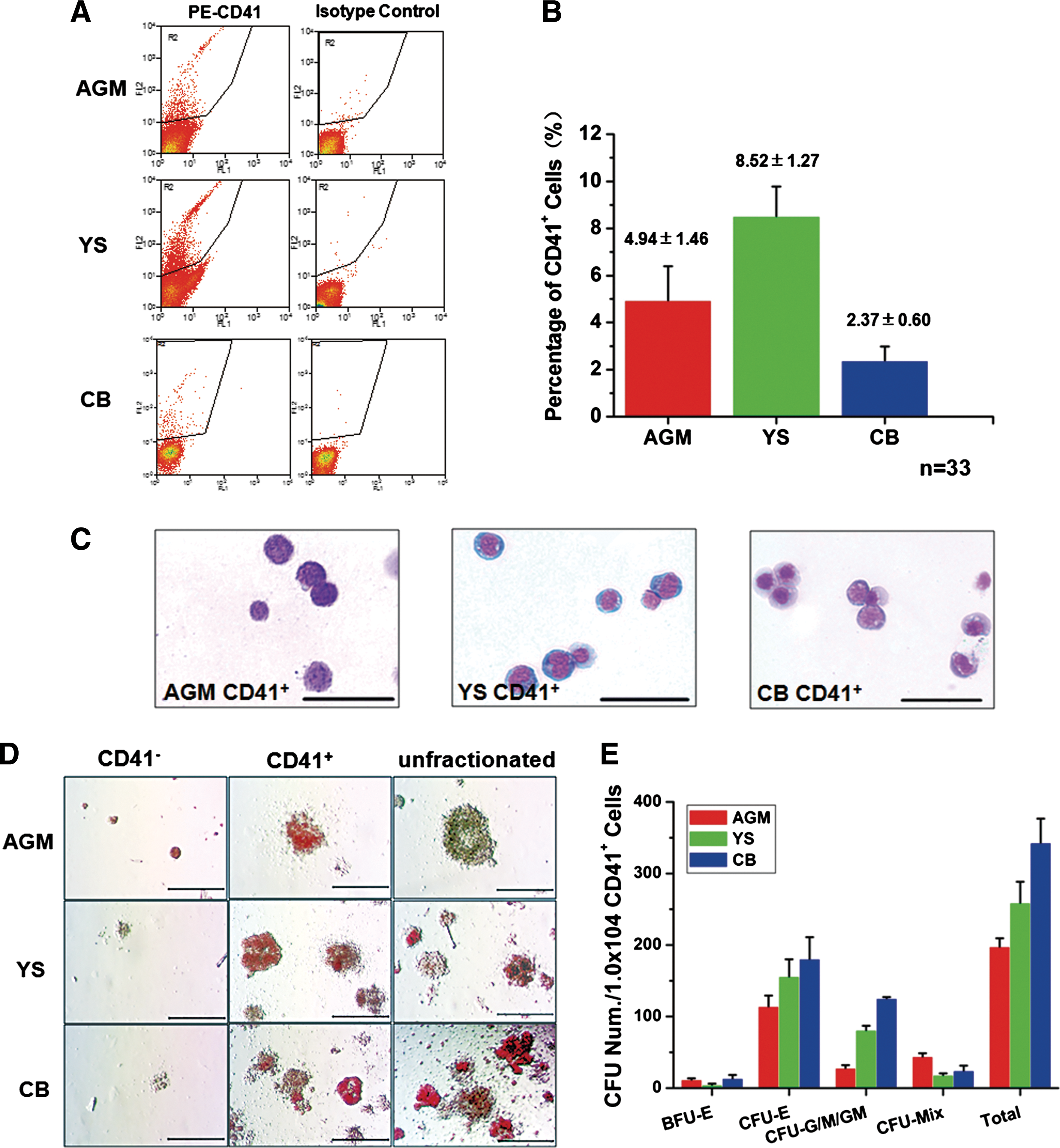
Fluorescence-activated cell sorting (FACS) and hematopoietic colony-forming unit-culture (CFU-C) assay of aorta-gonad-mesonephros (AGM), yolk sac (YS), and circulating blood (CB)-derived CD41+ cells. Collagenase-digested single cells of mouse AGM, YS, and CB-derived CD41+ cells at E11.0 were sorted by FACS.
AGM-, YS-, and CB-derived CD41+ cells contained different numbers of CFU-Cs
After sorting, the CD41+ cells were analyzed by Giemsa staining. The AGM-derived CD41+ cells demonstrated a relatively higher nuclear-cytoplasmic ratio than the YS- and CB-derived CD41+ cells (Fig. 1C). The hematopoietic differentiation potential of these CD41+ cells was assayed as previously described [3,12]. One week later, various types of erythromyeloid colonies were counted under the microscope. There were rare and small hematopoietic colonies in the CD41− subpopulation, while most of the hematopoietic colonies were derived from the CD41+ subpopulation (Fig. 1D). Then, the number of BFU-E, CFU-E, CFU-G/M/GM, and CFU-Mix colonies was statistically analyzed. A significant difference in CD41+ cell colony yield was observed in BFU-E (AGM vs. YS, P<0.05; YS vs. CB, P<0.05), CFU-E (AGM vs. CB, P<0.05), CFU-G/GM/M (AGM vs. YS, P<0.01; AGM vs. CB, P<0.01; YS vs. CB, P<0.01), CFU-Mix (AGM vs. YS, P<0.01; AGM vs. CB, P<0.05), and total colonies (AGM vs. YS, P<0.05; YS vs. CB, P<0.05; AGM vs. CB, P<0.01) (Fig. 1E).
Expression of mesenchymal markers in the AGM- and YS-derived CD41+ cells
Several pieces of evidence have indicated that the mesenchyme is associated with embryonic hematopoiesis [16,17], in addition to its supportive function. First, we used nested RT-PCR to analyze the expression of mesenchymal markers, including a-SMA, vimentin, myocyte enhancer factor 2C (MEF-2C), and secreted phosphoprotein 1 (SPP-1), as well as the hematopoietic/endothelial markers CD45 and vascular endothelial growth factor receptor (Flk-1). While both AGM- and YS-derived CD41+ cells were positive for the three types of markers, CB-derived CD41+ cells were negative for MEF-2C, SPP-1, and Flk-1. Of note, for a-SMA, the CB- and YS-derived CD41+ cells had a lower expression level than the AGM-derived CD41+ cells. The CB-derived CD41+ cells had a lower expression of vimentin than those derived from the AGM and the YS. The SPP-1 expression level in the AGM CD41+ cells was lower than that of the YS-derived cells (Fig. 2). Moreover, we examined the expression of the mesenchymal marker α-SMA in AGM and YS tissues by immunostaining (Supplementary Fig. S1; Supplementary Data are available online at
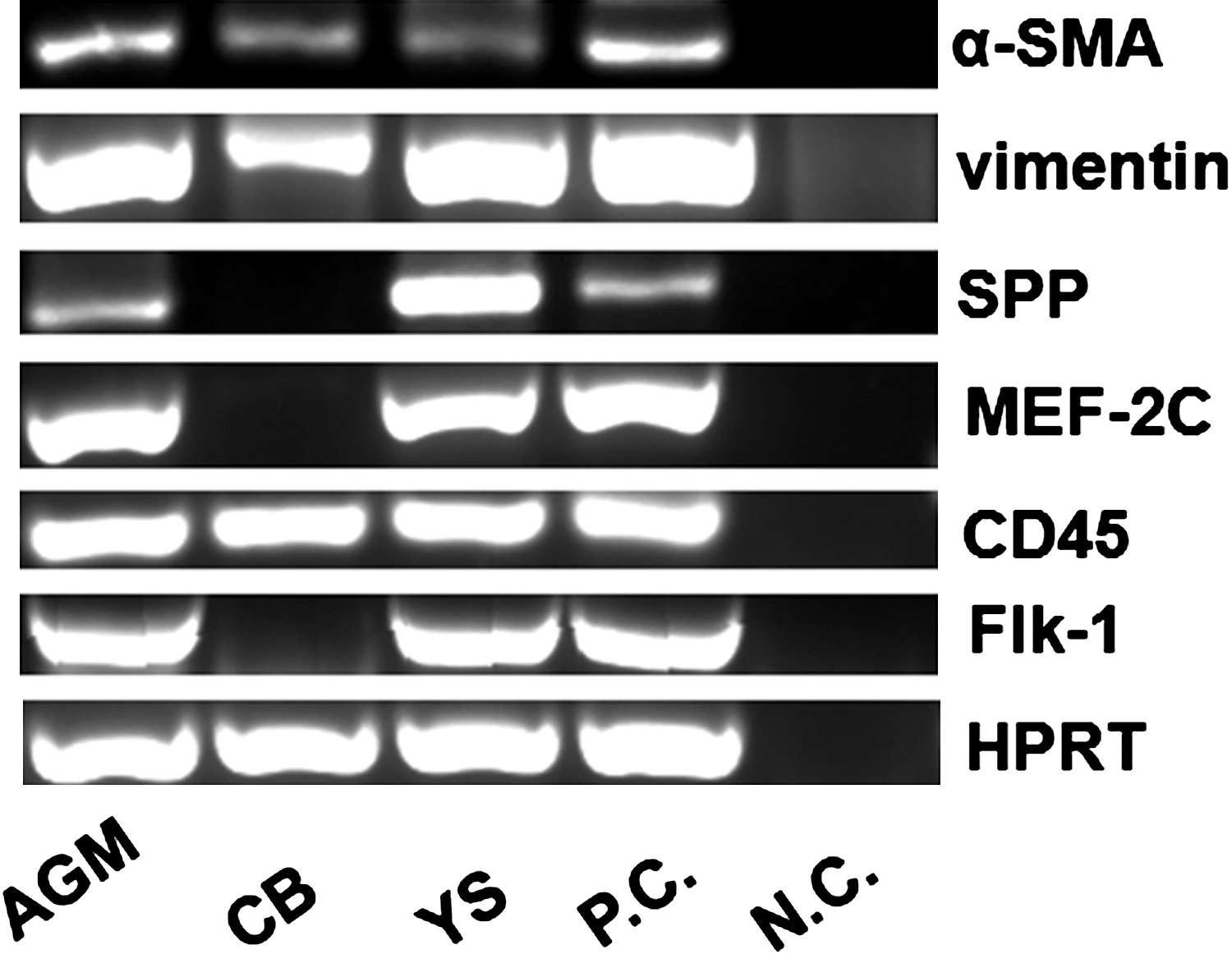
Analysis of mesenchymal marker expression in AGM and YS-derived CD41+ cells. Nested reverse transcriptase–polymerase chain reaction analysis of the freshly isolated CD41+ cells from mouse AGM, YS, and CB at E11.0. “P.C.” (positive control) indicates mouse fetal liver cDNA. “N.C.” (negative control) indicates water as the template.
AGM- and YS-derived CD41+ cells differentiated into fibroblast-like cells in vitro
Considering the existence of α-SMA+/CD41+ cells in mouse embryos, we presumed that the CD41+ population may have mesenchymal differentiation potential. To confirm this hypothesis, we cultured the isolated CD41+ cells from the AGM and the YS in a minimal mesenchymal differentiation system (10 ng/mL bFGF+10% FBS). After 7–10 days of culture, the primary CD41+ cells from both tissues gave rise to adherent cells, in addition to typical nonadherent hematopoietic cells (Fig. 3A
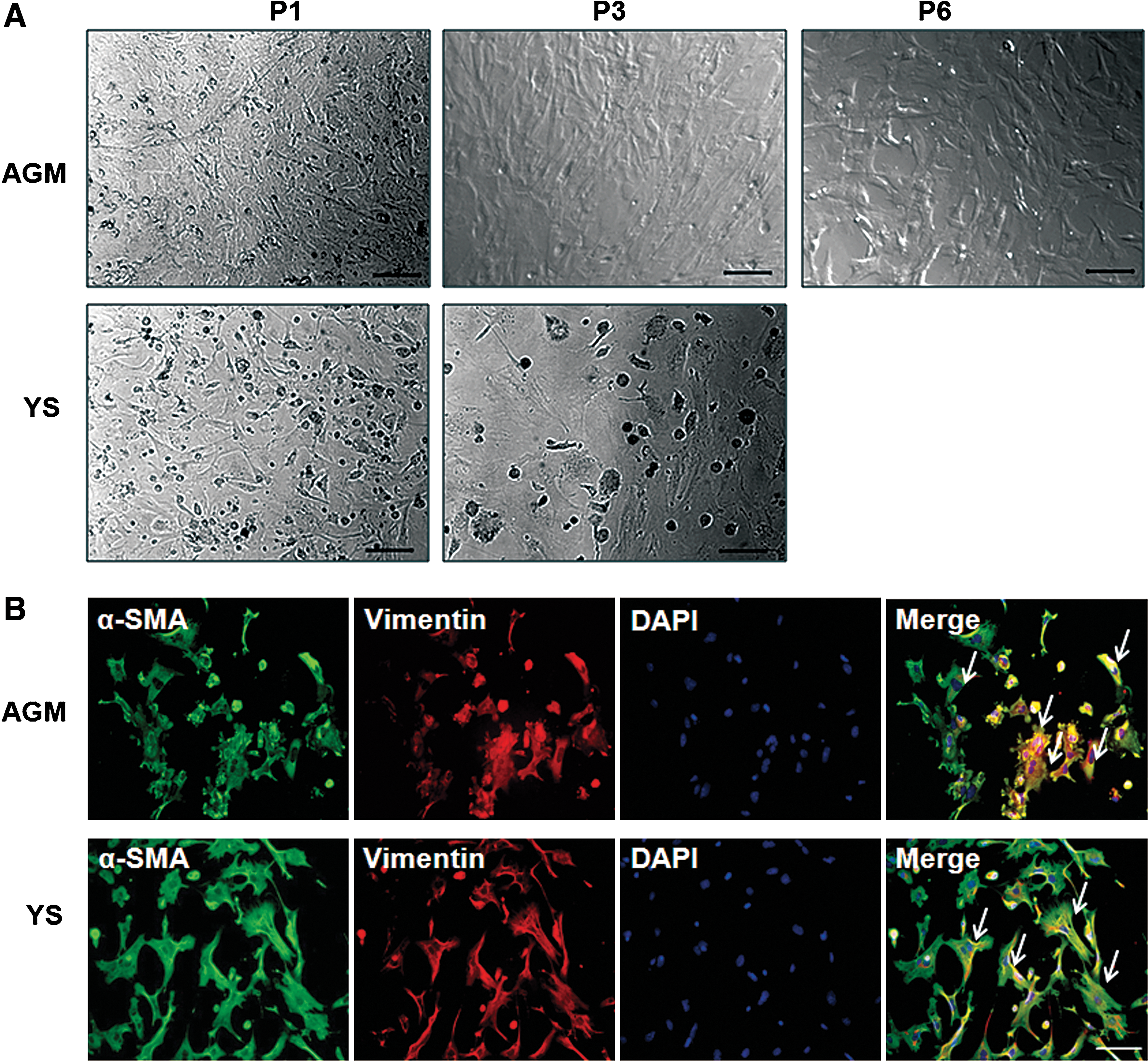
In vitro mesenchymal differentiation of CD41+ cells.
The AGM- and YS-derived CD41+ populations could differentiate into fibroblast-like cells in vivo
To further confirm the mesenchymal differentiation potential of CD41+ cells, we extended the in vitro study to an in vivo transplantation assay of mice treated with a single 6-Gy dose of whole-body irradiation with 60Co. Notably, this condition was not optimal for hematopoietic repopulation, but was appropriate for mesenchymal engraftment. The freshly fractionated GFP+/CD41+ cells and GFP−CD41− (partner cells) from the E11.0 AGM, YS, or CB of GFP-transgenic mice (Supplementary Fig. S2) were injected into the tail veins of irradiated adult mice.
We analyzed the overall hematopoietic engraftment at different time points. The peripheral blood (PB) samples were obtained via the tail vein at week 2, 3, and 4 after transplantation. The BM samples were collected at week 4 after transplantation. In Supplementary Fig. S3A, the chimerism of the AGM-derived GFP+ cells was 0.670%±0.431% (week 2), 1.115%±0.233% (week 3), and 4.535%±2.638% (week 4) in PB. The chimerism of YS-derived GFP+ cells was 0.370%±0.202% (week 2), 0.083%±0.055% (week 3), and 0.050%±0.162% (week 4) in PB. The chimerism of CB-derived GFP+ cells was 0.090%±0.070% (week 2), 0.087%±0.046% (week 3), and 0.053%±0.032% (week 4) in PB. In BM, the chimerism of GFP+ cells was 4.895%±0.190% (AGM), 0.22%±0.127% (YS), and 0.060%±0.062% (CB) 4 weeks after transplantation, as shown in Supplementary Fig. S3B. The overall engraftment of the AGM CD41+ population was higher than that of the YS and CB in both PB and BM. The hematopoietic repopulation gradually decreased in the recipients transplanted with YS CD41+ cells (Supplementary Fig. S3A). Although the overall engraftment of YS is lower than that of AGM, several GFP+ fibroblast-like cells derived from YS GFP+/CD41+ cells around blood vessels are clearly observed in the lungs of recipient mice.
Four weeks after transplantation, we analyzed the distribution of GFP+/CD41+ cells in the heart, liver, spleen, lung, kidney, and BM. The GFP+/CD41+ cells were mainly observed in the lung, BM (GFP+/CD45+ in most, data not shown), and spleen (data not shown). Importantly, we observed the engraftment of transplanted GFP+ cells from the AGM (Fig. 4) and YS (Fig. 5) in the lungs of recipient mice, as confirmed by anti-GFP antibody (Figs. 4A and 5A). These GFP+ cells were mostly located around the CD31+ blood vessels (Figs. 4B and 5B). As a membrane protein, epimorphin was observed to be well expressed on the engrafted GFP+/CD41+ cells (GFP protein was mostly located in the cytoplasm and the nuclei) (Figs. 4C and 5C). However, the other mesenchymal cell markers, including vimentin (Figs. 4D and 5D) and α-SMA (Figs. 4E and 5E), were observed to be partially expressed on the engrafted GFP+/CD41+ cells, with some GFP+ cells co-expressing CD31 (Figs. 4B and 5B). These results suggested that both the AGM- and YS-derived CD41+ cells could differentiate into fibroblast-like cells in vivo.

In vivo mesenchymal differentiation of AGM-derived CD41+ cells in injured mice. AGM-derived GFP+/CD41+ cells and GFP−/CD41− partner cells were co-transplanted into 60Co-irradiated mice via the tail vein. The mice were sacrificed 4 weeks after transplantation. AGM-derived GFP+/CD41+ cells in mouse lung frozen tissue sections were confirmed with anti-GFP antibodies

In vivo mesenchymal differentiation of YS-derived CD41+ cells in injured mice. YS-derived GFP+/CD41+ cells and GFP−/CD41− partner cells were transplanted into 60Co-irradiated mice via the tail vein. The mice were sacrificed 4 weeks after transplantation. YS-derived GFP+/CD41+ cells in mouse lung frozen tissue sections were confirmed with anti-GFP antibodies
Analysis of subsets in CD41+ cells with fibroblastic potential
After validating the fibroblastic potential of the CD41+ populations from the AGM and the YS, we tried to distinguish more detailed subsets of CD41+ cells with the fibroblastic potential.
First, we sorted the AGM and the YS cells based on the CD41 expression level (ie, the CD41high and CD41intermediate (int) subpopulations reported by Robin et al. [9]) (Fig. 6A). We found that the fibroblastic potential (α-SMA+/vimentin+ cells) was exclusively enriched in the CD41int rather than the CD41high subpopulation (Fig. 6B, C). The CD41high subpopulation failed to generate adherent fibroblast cells, and went on to die in the mesenchymal differentiation medium (Fig. 6A). Based on the report by Robin et al. that the HSC potential is enriched in CD41int but not in CD41high (with a high proportion of megakaryocytic lineage cells [9]), we proposed that both immature hematopoietic and fibroblastic potential was enriched in the CD41int subpopulation, while the CD41high subpopulation contained more differentiated hematopoietic cells.

Fibroblastic differentiation of the CD41int and CD41high subsets derived from AGM and YS tissues.
Next, we categorized the CD41+ cells by CD34 expression. It is interesting that CD34 is reported to be expressed in endothelial cells, hematopoietic progenitors, and a subset of mesenchymal cells in fetal mouse tissues [18,19]. Therefore, we further sorted the CD41int cells into CD34+ and CD34− subpopulations (Fig. 7A). Evident fibroblastic potential was detected in the CD41int/CD34− AGM cells and in both YS cell subsets (Fig. 7).

Fibroblastic differentiation of the CD41int/CD34− and CD41int/CD34+ subsets derived from AGM and YS tissues.
We also tested the purity of the sorted CD41int population. The purity of AGM-derived CD41int and YS-derived CD41int is 94.3% and 97.7%, respectively (Supplementary Fig. S4). These results provide evidence for the reliability of our FACS data.
The CD41+ fibroblastic potential was significantly reduced in embryonic circulation
Since the fibroblastic potential of the AGM- and YS-derived CD41+ cells was detected, we asked whether these CD41+ cells with fibroblastic potential would migrate into the embryonic circulation. Under the same culture conditions, the CB CD41+ cell-derived adherent cells could not proliferate even 2 weeks after plating. In morphology, these adherent cells resembled macrophage cells (Fig. 8A). Immunofluorescence staining demonstrated rare α-SMA-positive cells or no vimentin-positive cells in the culture (Fig. 8B). More importantly, almost no engrafted GFP+ cells were detectable in the lungs of recipient mice injected with the CB GFP+/CD41+ cells (Fig. 8C). These results indicated that CB CD41+ cells have almost no potential to be differentiated into fibroblast-like cells in vitro and in vivo.
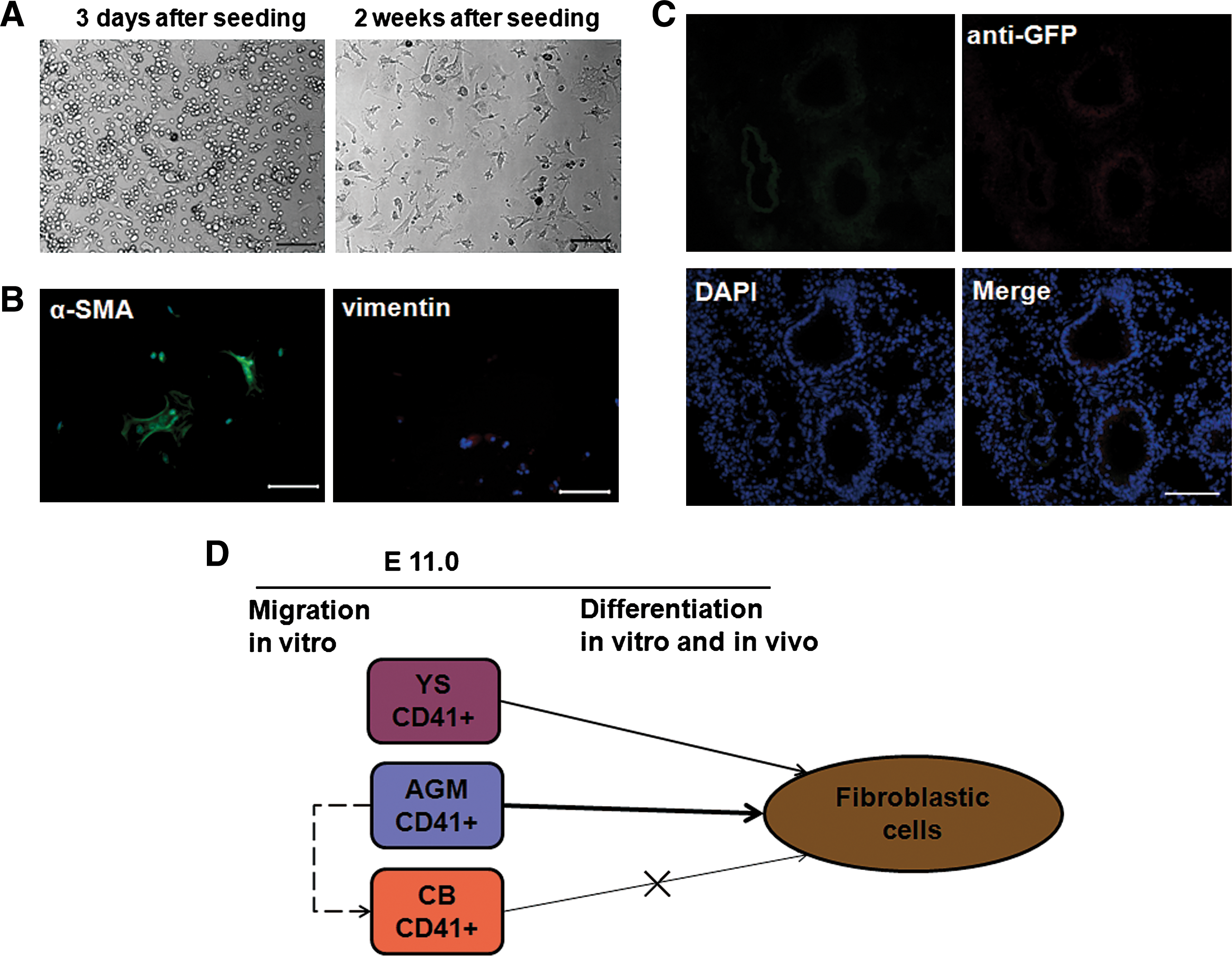
Fibroblastic potential of CB-derived CD41+ cells.
The AGM- but not the YS-derived CD41+ population could respond to CB plasma
As previously reported [11], migration is another important characteristic of hematopoietic and mesenchymal stem cells (MSCs), particularly in embryos. At E11.0, definitive hematopoietic cells migrate into the fetal liver, a site where hematopoietic cells can expand and mature. To compare the migration ability of the AGM and YS-derived CD41+ cells, we established a transwell migration assay. The same number of CD41+ cells (1–2×105 cells) was placed on the upper insert, while the CB plasma was placed on the outer well. After a 12 h incubation, the cells on the bottom plate were stained with DAPI and counted as migrated cells (Supplementary Fig. S5A). We determined the number of migrated CD41+ cells in the AGM, YS, and control experiments by averaging the amounts from ten random fields observed under the fluorescence microscope. The results showed that the AGM-derived CD41+ cells respond to the chemotaxin of CB plasma more strongly (compared with the control, P<0.01) than those of the YS (compared with the control, no significance) (Supplementary Fig. S5B).
Discussion
The hematopoietic potential of CD41+ cells has been extensively investigated [2,6 –8]. Hematopoiesis is believed to be closely associated with the development of the endothelium and mesenchyme before the fetal liver stage. Our data provided the first evidence that the AGM- and YS-derived CD41+ cells harbored fibroblastic potential in vitro and in vivo, in addition to their canonical hematopoietic capacity.
These fibroblast-like cells were shown to express some fibroblastic markers. Among these markers, vimentin is the major cytoskeleton component of mesenchymal cells and is often used as a marker of mesenchymally derived cells. Epimorphin is a mesenchymal cell-associated membrane protein found by Hirai et al. [20] that is highly expressed around blood vessels [21], the bile duct [22,23], and other duct structures in the mammary glands and pancreas [24]. α-SMA is commonly used as a marker of myofibroblasts/fibroblasts [25]. Considering the morphologic appearance and the myofibroblast/fibroblast marker expression in culture and the in vivo transplantation experiments, we can assume that these CD41+ cells differentiate into myofibroblast/fibroblast cells. Meanwhile, we have noticed that not all the engrafted GFP+/CD41+ cells were co-stained fibroblastic markers (vimentin or α-SMA), as shown in Figs. 4 and 5. There are two possibilities: (i) Only a small portion of the CD41+ population have the fibroblastic potential, which is supported by the analysis of subsets in CD41+ cells with a fibroblastic potential. These unique cells are enriched in CD41int subpopulation, particularly in the CD41int/CD34− subset in AGM. (ii) Fujita, et al. [26] has also reported that the HSC-derived myofibroblasts observed in the acute phase gradually became α-SMA-negative fibroblasts during the chronic phase in a mouse model of myocardial infarction. This may be an alternative explanation that not all the engrafted GFP+/CD41+ cells were co-stained fibroblastic markers. Taken together, these CD41+ cells-differentiated myofibroblast/fibroblast cells identified in the study may be not the same as the wild-type fibroblasts, which need to be further elucidated by gene expression profile analysis.
To our knowledge, these CD41+ cell-derived myofibroblast/fibroblast cells may be of three origins.
(i) The CD41+ cell-derived myofibroblast/fibroblast cells may arise from a putative common precursor for hematopoietic and mesenchymal cells, which is much debated [27 –29]. It is difficult to provide direct evidence for this putative origin in vivo; an in vitro alternative single-colony model is further needed, as previously described for a hemangioblast model [3,30].
(ii) The CD41+ cell-derived myofibroblast/fibroblast cells may arise from HSCs as Ogawa et al. and Fujita et al. previously reported. Their findings established the HSC origin of myofibroblast/fibroblasts in both in vitro and in vivo models [27,31 –34]. Our findings here demonstrated for the first time that CD41+ cells can be differentiated into myofibroblasts/fibroblasts, providing additional supporting evidence for HSC-derived fibroblasts in embryonic blood-forming sites.
(iii) The CD41+ cell population may be heterogeneous with regard to mesenchymal and hematopoietic potential. The simultaneous generation of hematopoietic cells and MSCs was observed in both a human ESC differentiation system [35] and the mouse AGM region [36]. However, we cannot rule out the possibility that a rare number of MSCs exist in AGM-derived CD41+ cells. In fact, Kaiser et al. has reported that BM cells can give rise to MSCs in cultures with a heterogeneous CD34 and CD45 phenotype [37]. As a counterpart of CD45 in embryonic development, CD41 may be also expressed in a rare population of MSCs or in mesenchymal precursor cells.
Little is known about the functional role CD41 plays on the surface of hematopoietic precursors, with the exception of a report that CD41 regulates cell numbers at sites of hematopoiesis [38]. In addition, Berlanga et al. showed that CD41 influences mast cell adhesion properties in BM [39]. Here, we want to know whether CD41 is involved in the migration of definitive hematopoietic cells during embryonic development. To our surprise, YS-derived CD41+ cells have almost no responsiveness to CB plasma, whereas AGM-derived CD41+ cells respond significantly. Moreover, it can be concluded that these AGM-derived CD41+/α-SMA+ cells may never or only rarely migrate into the CB, based on our results that CB-derived CD41+ cells have no capacity to differentiate into fibroblastic cells in vitro and in vivo. These results are consistent with the two waves of colonization into the embryonic liver by HSCs from the AGM region (from E11.0 to E12.0) and the YS (from E12.0 to E13.0) [40]. However, these differences in migration ability and fibroblastic potential cannot be completely explained as the regulation of cell adhesion or cell number by the CD41 protein expressed in the CD41+ population [38,39]; additional regulation of other molecules from specific cell-located microenvironments may be required.
Our results shed light on the existence of definitive hematopoietic cells with a fibroblastic potential. However, further elucidation of the origin and the functional role of CD41+ cells with fibroblastic potential during embryonic development is needed. The optimal systems for such a study are CD41-Cre transgenic mice [38] or zebrafish [41], which may provide direct evidence by in vivo cell lineage tracing. These studies of the hematopoietic cell (or HSC) origin of mesenchymal cells may also facilitate the development of novel therapeutic approaches for diseases and injuries of the connective tissues [42].
In summary, our data revealed the following to be true at E11.0 (i) In addition to hematopoietic potential, the AGM and YS-derived CD41+ cells had the potential to differentiate into epimorphin+/α-SMA+/vimentin+ cells, assumed to be fibroblast cells. (ii) Furthermore, this unique population with fibroblastic potential expressed intermediate rather than high levels of CD41, being negative for CD34 in the AGM region. (iii) The AGM-derived CD41+ cells may have a higher capacity than YS-derived CD41+ cells to migrate into circulation. The CD41+ cells with a fibroblastic potential may rarely migrate into the blood stream (Fig. 8D).
Footnotes
Acknowledgments
The authors would like to thank Dr. Yohei Hirai (Department of BioScience School/Graduate School of Science and Technology, Kwansei Gakuin University, Japan) for kindly providing the monoclonal anti-epimorphin antibody MC-1. This work was supported by the National High Technology Research and Development Program of China (No: 2006AA02A107, 2011AA020109), the Major State Basic Research Program of China (No: 2010CB945504, 2011CB964804), and the National Nature Science Foundation of China (No: 81101812, 30871410 and 30010180).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.