
Abstract
We tested the hypothesis that simultaneous transgenic overexpression of a select quartet of growth factors activates diverse signaling pathways for mobilization and participation of various stem/progenitor cells for cardiogenesis in the infarcted heart. Human insulin growth factor-1 (IGF-1), vascular endothelial growth factor (VEGF), stromal cell–derived factor-1 (SDF-1a), and hepatocyte growth factor (HGF) plasmids were synthesized and transfected into skeletal myoblasts (SM) from young male wild-type or transgenic rats expressing green fluorescent protein (GFP). Overexpression of growth factors in transfected SM (TransSM) was confirmed by reverse transcription polymerase chain reaction, western blotting, and fluorescence immunostaining. Using our custom-made growth factor array and western blotting, multiple angiogenic and prosurvival factors were detected in TransSM, including secreted frizzled related protein-1,2,4,5, matrix metalloproteinases-3 and 9, connexin-43, netrin-1, Nos-2, Wnt-3, Akt, MAPK42/44, Stat3, nuclear factor kappa B (NFκB), hypoxia-inducible factor 1 (HIF-1α), and protein kinase C (PKC).
The conditioned medium (CM) from TransSM was cytoprotective for cardiomyocytes following H2O2 treatment [P<0.01 vs. CM from native SM (NatSM)], promoted a higher transwell migration of human umbilical cord vein endothelial cells (223.3±1.8, P<0.01) and in vitro tube formation (47.8±1.9, P<0.01). Intramyocardial transplantation of 1.5×106 TransSM (group-3) in a rat model of acute myocardial infarction induced extensive mobilization of cMet+, ckit+, ckit+/GATA4+, CXCR4+, CD44+, CD31+, and CD59+ cells into the infarcted heart on day 7 and improved integration of TransSM in the heart compared to NatSM (group 2) (P<0.05). Extensive neomyogenesis and angiogenesis in group-3 (P<0.01 vs. group-2), with resultant attenuation of infarct size (P<0.01 vs. group-2) and improvement in global heart function (P<0.01 vs. group-2) was observed at 8 weeks. In conclusion, simultaneous activation of diverse signaling pathways by overexpression of multiple growth factors caused massive mobilization and homing of stem/progenitor cells from peripheral circulation, the bone marrow, and the heart for accelerated repair of the infarcted myocardium.
Introduction
T
BMSC, SM, peripheral blood-derived endothelial progenitor cells (EPC) and resident cardiac stem/progenitor cells have all shown considerable myogenic and vasculogenic potential in both experimental and clinical settings. Besides, all these cell types have distinct mechanisms of imparting functional benefits. SM differentiate into skeletal muscle fibers, which serve as a scaffold to support the weakened infarcted myocardium, prevent left ventricular (LV) remodeling, and improve the diastolic properties of the heart [5,6], besides releasing plethora of cytokines [7]. On the contrary, besides myogenic differentiation, BMSC secrete growth factors, which act in a paracrine manner to prevent infarct size expansion via cytoprotection and angiogenesis [8,9]. Peripheral blood-derived stem/progenitor cells have a vasculogenic activity in addition to adopting a cardiac phenotype [10], and cardiac muscle-derived stem/progenitor cells may adopt phenotypes of all the cell types that are required for myocardial regeneration [11,12]. The main aim of the study was to adopt a multiprong strategy and simultaneously activate diverse signaling pathways by transplantation of SM transfected with quartet of growth factors (TransSM) having diverse functions, that is, survival, proliferation, induction of gap junctions, release of paracrine factors, and mobilization of BMSC, EPC, and cardiac stem cells for regeneration of the infarcted myocardium. To this end, we programmed SM with a select group of growth factors, including SDF-1α, VEGF, and HGF, and transplanted the genetically modified SM into the infarcted heart. Despite their controversial proarrhythmogenic nature, we opted for SM as carriers of the select quartet of growth factors for delivery to the heart due to their proven ability as excellent carriers of therapeutic gene/s [13]. Secondly, their myogenic differentiation postengraftment serves as a scaffold to attenuate LV remodeling [5]. We proposed that IGF-1 would enhance Cx-43 expression in TransSM to support their engraftment in the infarcted myocardium.
Materials and Methods
All experimental procedures were performed in accordance with the standard human care guidelines of the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication No. 85-23, revised 1996) and protocol No. 06-03-13-03 approved by the Institutional Animal Care and Use Committee, University of Cincinnati. All in vitro experiments were repeated (n=3) to ensure reproducibility of the data.
Isolation and characterization of SM
Native SM (NatSM) were isolated from skeletal muscle biopsies from the hindlimbs of young male Fisher-344 rats and characterized by flow cytometry and immunostaining as described earlier and detailed in Supplementary Materials and Methods [14] (Supplementary Data are available online at
Preparation of plasmids and cell transfection
Plasmids encoding for SDF-1, HGF, IGF-1, and VEGF were prepared as described in Supplementary Data. For transfection, cells were cultured to 80%–90% confluence and separately transfected with 1 of 4 plasmids (IGF-1, VEGF, SDF-1α, or HGF) using the cationic vector Lipofectamine-2000 (Invitrogen). Lipid–DNA transfection complexes were prepared by using a DNA (μg) to Lipofectamine-2000 (μL) ratio of 1:2.5 in the basal Dulbecco's modified Eagle medium (DMEM). After 20 min incubation at room temperature, transfection complexes were added to the cells in the presence of 10% fetal bovine serum (FBS), mixed gently, and incubated for 8 h at 37°C in a CO2 incubator. After removal of transfection complexes, cells were cultured in the DMEM containing 10% FBS 72 h before pooling different groups of cells transfected with their respective growth factor plasmid in 1 dish. TransSM were cultured at 37°C in a 5% CO2 incubator for 24 h before their use in in vitro and in vivo studies. Transfected cells for each plasmid either alone or after pooling were characterized by fluorescent immunostaining, reverse transcription polymerase chain reaction (RT-PCR), and western blotting as detailed in Supplementary Materials and Methods. The specific antibodies and primer sequences used are shown in Supplementary Tables S1 and S2 respectively.
RT-PCR studies
RT-PCR was performed for quantification of a panel of prosurvival and angiogenic genes in the 7 days heart tissue samples using our custom-made gene array (Supplementary Table S3).
Cell migration and tube formation assays
The conditioned medium (CM) from TransSM and NatSM were used for cell migration and tube formation assays as detailed in Supplementary Materials and Methods and described earlier [15].
Cytoprotective activity of CM from various SM groups
The CM from various treatment groups of SM was used as described in Supplementary Materials and Methods. Lactate dehydrogenase (LDH) leakage was measured for each sample using the LDH Assay kit as per the manufacturer's instructions and described in Supplementary Materials and Methods. Terminal dUTP nick-end labeling assay (TUNEL) was performed on cells with an In-Situ Cell Death Detection kit (Roche Diagnostics) as per the manufacturer's instructions. The degree of apoptotic cell death was determined by counting the total number of TUNEL-positive nuclei per microscopic field (400×). Caspase-3 activity assay was performed using the commercial kit (Calbiochem) as per the manufacturer's instructions.
In vitro gap junction formation between TransSM and cardiomyocytes and their functional assessment
Neonatal cardiomyocytes were isolated from newborn (1–3 days) rat pups using Neonatal Cardiomyocyte Isolation System (Worthington Biochemical) as described in Supplementary Materials and Methods. Functional assessment of gap junctions between TransSM or NatSM and cardiomyocytes was performed by the Lucifer Yellow dye transfer method (a Cx-43 dye transfer, 0.5 ng/mL; Sigma-Aldrich) as described in Supplementary Materials and Methods [16]. Parallel experiments were performed by heptanol (an inhibitor of Cx-43, 0.5 mM; Sigma-Aldrich) treatment of the cells before and during Lucifer Yellow loading. The functional status of Cx-43 between the adjacent cells was characterized by the length of the dye expansion from the scrape-loaded cells in each cell population. To elucidate electrophysiological coupling between TransSM and cardiomyocytes in cocultures, the cells were loaded with Fluo-4 acetoxymethyl ester (a green fluorescent calcium indicator, 6 μM; Molecular Probes) and calcium sparks were measured as detailed in Supplementary Materials and Methods. Briefly, 3 days after coculture, the cells were loaded with calcium indicator in the M-199 medium for 20 min at 37°C. After washing, cells were transferred to microscopic chambers in the Tyrode's solution supplemented with 20% FBS. Calcium sparks were measured using the confocal microscope (Carl Zeiss LSM-510) in a line scan mode with excitation wavelength of 488 nm and emission wavelength of ≥510 nm.
In vivo studies
Rat model of myocardial infarction and SM transplantation
An experimental model of acute myocardial infarction was developed in young female Fisher-344 rats by permanent occlusion of the left anterior descending coronary artery as described earlier [17]. The animals were grouped to receive intramyocardial injections (4–5 sites/heart) of 70 μL basal DMEM without cells (group-1) or containing 1.5×106 NatSM (group-2) or TransSM (group-3). The chest was closed and the animals were allowed to recover. Buprenex (0.1 mg/kg b.i.d) was administered for 24 h to alleviate pain. For postmortem studies, animals were sacrificed using an overdose of sodium pentobarbital at stipulated time points after their respective treatment and the heart tissue samples were excised for further studies. PCR was performed for quantification of the sry-gene for assessment of donor cell survival 7 days (n=3/group) after their respective treatment in various treatment groups of animals [18].
Histological studies
For measurement of infarction size, the heart was arrested in diastole by intravenous injection of cadmium chloride and fixed in 10% buffered formalin. Histological sections of 6-μm thickness were stained with Mason's trichrome and analyzed using computer-based planimetry with ImageJ analysis software (version-1.6065; NIH). To determine the fate of transplanted stem cells, histological tissue sections were immunostained with a primary antibody to green fluorescent protein (GFP) (Abcam) and detected with a secondary antibody conjugated to Alexa Fluor-488 (Molecular Probes). Blood vessel density was assessed by immunostaining for the vonWillebrand Factor-VIII (vWFactor-VIII) and smooth muscle actin-specific primary antibodies. Blood vessel density was expressed as the number of vessels per microscopic surface area (0.74 mm2) at 200× magnification. The other primary antibodies used in immunohistological studies were Ki-67 for cell proliferation, desmin, cardiac actinin, myosin heavy chain, and Cx-43 for myogenesis and integration of neofibers with host myocytes, and CD31, c-kit, CD44, CD45, and CXCR4 for stem/progenitor cells mobilized into the infarcted myocardium. The antibodies used are listed in Supplementary Table S1.
The heart function studies
The heart function was measured at 8 weeks after their respective treatment (group-1 n=6; groups-2 and -3 n=8/group) as detailed in Supplementary Materials and Methods [17]. The heart function was determined by transthoracic echocardiography using the Compact Linear Array probe CL10-5 on HDI/5000 SONOS CT (HP-Company). Echocardiograms were recorded through parasternal long-axis and short-axis views. Indices of contractile function, including the LV ejection fraction (LVEF) and the LV fractional shortening (LVFS) were calculated using following relations: LVFS={(LVEDD – LVESD)/LVEDD}×100 and LVEF=(LVEDD3−LVESD3)/LVEDD3×100 [left ventricle end-systolic (LVESD) and end-diastolic (LVEDD)] and results were expressed as percentage.
Statistical analysis
Statistical analysis was performed with Statview-5.0 software. All values were expressed as means±standard errors using the Student t-test, a one-way analysis of variance (ANOVA), or a 2-way factorial ANOVA for repeated measures. P<0.05 was considered significant.
Results
In vitro studies
SM purification and transfection with growth factor transgenes
SM were successfully isolated from skeletal muscle biopsies obtained from male donor Fisher-344 rats using our standard protocol. Fluorescence-activated cell sorting (FACS) analysis demonstrated that purified SM culture was more than 90% and 62% positive for desmin and CD56 expression, respectively (Supplementary Fig. S1A, B). Plasmids encoding for IGF-1, VEGF, SDF-1α, and HGF were successfully constructed (Supplementary Fig. 1C) and used for transfection of SM. Expression of growth factors was increased in SM after transfection with the respective individual growth factor plasmid as was evident from RT-PCR (Fig. 1A), western immunoblotting (Fig. 1B), and fluorescence immunostaining (Supplementary Fig. S1D).

Characterization of SM in vitro.
Gene expression profiling of TransSM
Besides a multifold increase in transgenic overexpression of the 4 growth factors (Fig. 2A), RT-PCR using our custom-made array of survival and growth factors showed upregulation of a range of prosurvival and angiogenic gene expression in TransSM (Fig. 2B). Significant among the upregulated genes in TransSM included secreted frizzled related protein (Sfrp)-1, Sfrp-2, Sfrp-4, Sfrp-5, netrin-1, inducible nitric oxide synthase (iNOS), and matrix metalloproteinases (MMP)-3 and MMP-9. We also observed elevated expression of cardiac transcription factors, GATA-4 and Mef-2c, besides significantly higher expression of Cx-43. Western blot showed activation of survival signaling molecules and transcription factors, including pAkt, p44/42MAPK, pStat3 and HIF-1α, and pPKC(pan), in TransSM as compared to NatSM (Fig. 2C). Another important feature of TransSM was a significantly higher gene (Fig. 2B) and protein level induction of Cx-43 (Fig. 2C), the key molecule for electromechanical coupling between engrafted cells and host cardiomyocytes.

Activation of multiple signaling pathways in TransSM.
Cytoprotective and angiogenic effects of CM from TransSM
The cytokine and growth factor-rich CM from TransSM (TransCM) was protective for H9C2 and prevented cell death under oxidative stress. LDH release (a marker of cellular injury) was significantly reduced in H9C2 treated with TransCM (P<0.01 vs. untreated H9C2; Fig. 3A) upon subsequent exposure to 100 μM H2O2. These results were supported by TUNEL assay, which demonstrated a reduced number of TUNEL-positive H9C2 pretreated with TransCM upon subsequent exposure to oxidative stress (P<0.01 vs. untreated H9C2 and NatCM-treated cells; Fig. 3B). Similarly, pretreatment of H9C2 with TransCM reduced caspase-3 activity in the cells upon subsequent exposure to 100 μM H2O2 (P<0.01 vs. untreated NatSMs; Fig. 3C). TransCM also promoted human umbilical cord vein endothelial cells (HUVEC) migration in the in vitro transwell migration system as compared with NatCM (P<0.01; Fig. 3D). In vitro tube formation assay on Matrigel showed higher branch points at 6 h after incubation with TransCM (P<0.01 vs. NatCM; Fig. 3E).
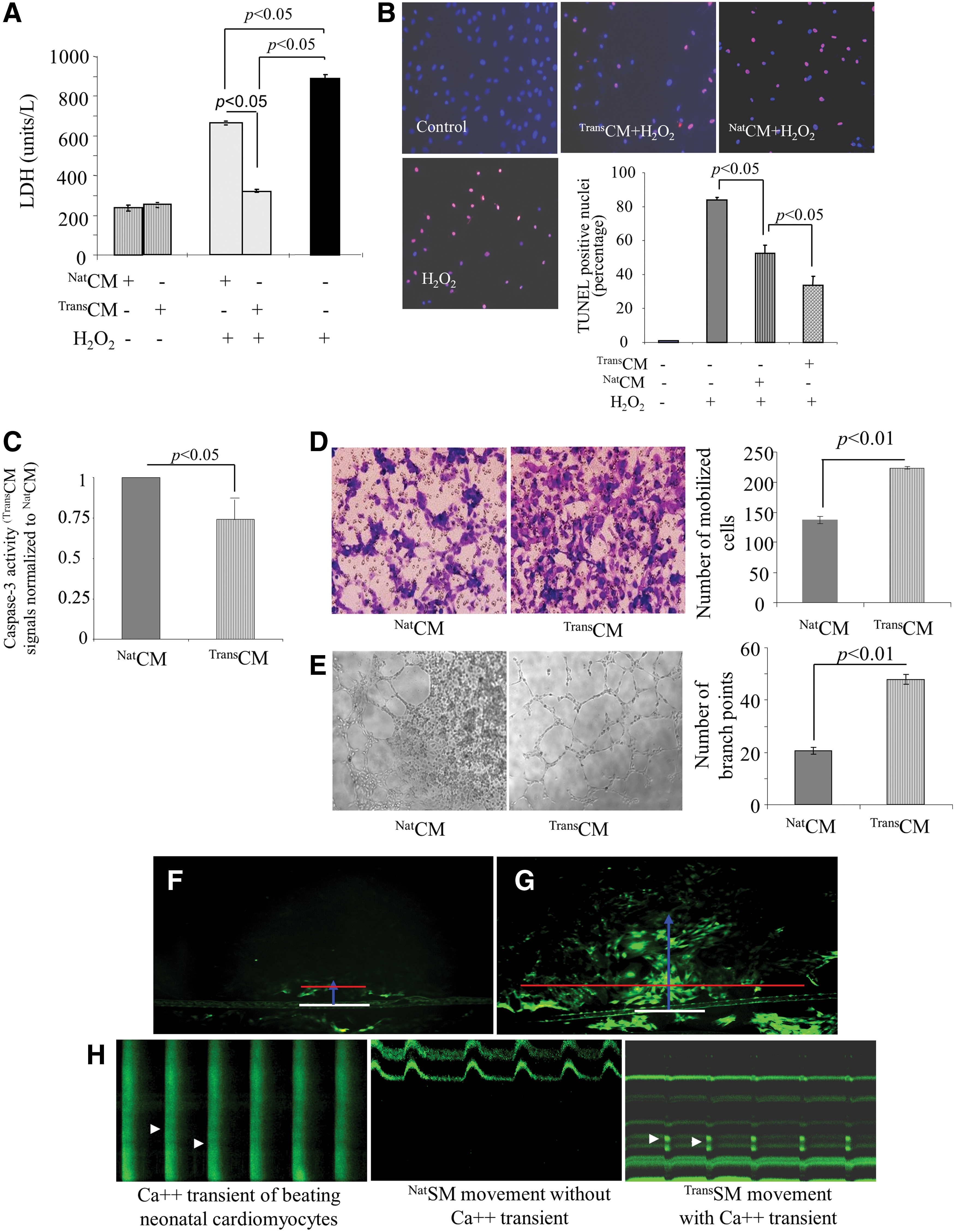
Cytoprotective and in vitro angiogenic effects of TransCM, and expression of gap junctional proteins in TransSM.
In vitro gap junction formation and their functional assessment
We cocultured neonatal cardiomyocytes with NatSM (Supplementary Fig. S2A) or TransSM (Supplementary Fig. S2B). We observed that Cx-43 events between TransSM and cardiomyocytes were significantly higher as compared to those developed between cardiomyocytes and NatSM (P<0.05; Supplementary Fig. S2C). Similar observations were made after immunostaining of cardiomyocytes cocultured with NatSM (Supplementary Fig. S2E) and TransSM (Supplementary Fig. S2D) for N-cadherin expression. We observed that N-cadherin events between TransSM and cardiomyocytes were significantly higher as compared to those developed between cardiomyocytes and NatSM (P<0.01; Supplementary Fig. S2E).
Dye transfer assay was performed using Lucifer Yellow, a dye, which specifically transfers through functional Cx-43 (Fig. 3F, G). We observed markedly higher expansion (indicated by red line) and extension (indicated by blue line) of the dye transferred from scratch point (white baseline) in TransSM (Fig. 3G) as compared to NatSM controls (Fig. 3F). Given that dye transfer (green fluorescence) was mediated via Cx-43 both in length (blue line) and width (red line) from point of scratch (white line), we inferred that Cx-43 expression was higher in TransSM as compared to NatSM. Figure 3H shows representative figures of calcium sparks to identify electrophysiologic coupling between SM (NatSM or TransSM) and neonatal cardiomyocytes on day 3 after coculture.
In vivo studies
All animals (n=16/group) survived the full length of study and there were no deaths related with the cell transplantation procedure. Each treatment group of animals consisted of 2 subgroups that were analyzed at 7 days and 8 weeks time points after their respective treatment. In addition, the subgroup harvested on day 7 consisted of 3 animals each for molecular and histological studies.
RT-PCR showed that the transplanted TransSM continued to express higher expression of transgenic growth factors, including SDF-1, HGF, VEGF, and IGF-1, 7 days after engraftment (Supplementary Fig. S3). The transplanted male TransSM showed better survival in the infarcted heart (Fig. 4A). PCR for the sry-gene in female rat heart on day 7 revealed extensive survival of the donor male TransSM in group-3 (P<0.05 vs. NatSM in group-2; Fig. 4A). However, we did not observe the sry-gene in DMEM-injected female animal hearts in group-1, which served as a negative control. These results were supported by immunohistological studies, which visualized the extensive presence of GFP+ TransSM at the site of cell graft in group-3 as compared to GFP+ NatSM in group-2 (Fig. 4B and Supplementary Fig. S4A). Besides, the transplanted GFP+ SM were found elongated as neofibers, which were aligned with host muscle architecture in both cell transplanted animal groups. However, double fluorescence immunostaining for GFP and Cx-43 showed that only TransSM expressed Cx-43 (red fluorescence) in the infarcted heart, whereas no Cx-43 was observed in NatSM postengraftment (Fig. 4C and Supplementary Fig. S4B). These observations were consistent with our in vitro data that TransSMs expressed high Cx-43 (Fig. 2). Fluorescence immunostaining for Ki67, a protein expressed only in nuclei of cycling cells, showed that TransSM proliferated in the infarcted myocardium more as compared to NatSM (P<0.01; Supplementary Fig. S4C). The identification of proliferating SM and their discrimination from dividing inflammatory cells in their surroundings was carried out by superimposition of GFP (green) and Ki67 (red) and 4′,6-diamidino-2-phenylindole (blue) nuclear staining (Supplementary Fig. S4C).

TransSM survived and engrafted better in the infarcted heart and promoted stem/progenitor cell mobilization.
Stem and progenitor cell homing into myocardium
The histological sections from 7-day harvested animal heart tissues from different treatment groups were immunostained for the presence of various lineages of stem and progenitor cells, which were mobilized into the infarcted heart (Fig. 4D, E and Supplementary Fig. S4D). The number of ckit+, ckit+/Gata-4+, CD31+, CXCR4+, and CD44+ cells mobilized into the infarcted heart was significantly higher in group-3 as compared to group-2. A higher number of these homed-in cells was observed in the infarct and peri-infarct areas in comparison with contralateral regions in the heart in both groups-2 and -3. Besides stem/progenitor cell mobilization, RT-PCR showed a higher expression of CD59 (also known as protectin, which is broadly expressed on hematopoietic and nonhematopoietic cells and is known for its inhibitory effects on cytolytic activity of complement) in group-3 hearts after 7 days of transplantation (P<0.01 vs. group-2; Fig. 4F, G).
Infarction size and heart function studies
Histological sections cut at the midpapillary muscle level followed by Masson's trichrome staining showed transmural infarction in all the animals at 8 weeks after coronary artery ligation (Fig. 5A). Noticeable LV wall thinning was observed at 8 weeks in the DMEM-treated group-1, whereas LV wall thickness was considerably retained in the group-3 animals. Hematoxylin/eosin staining showed an extensive myogenic activity in group-2 and group-3 animal hearts unlike the DMEM treated group-1, which had more scar tissue (Fig. 5B). Infarct size expansion was significantly more in the DMEM-treated group-1 (37.6%±0.7%) as compared to group-2 (26.6%±0.8%) and group-3 (22.2%±0.6%; P<0.01 group-3 vs. group-2; Fig. 5A). Transthoracic echocardiography of animals at 8 weeks after their respective treatment showed significant differences in the indices of LV function and chamber dimensions between the various treatment groups (Fig. 5C, D, and Supplementary Fig. S5). LVEF (49.0%±1.2%) and LVFS (20.7%±1.2%) were significantly improved in the TransSM-treated group-3 as compared to the control group-2 (42.9%±3.1% and 17.3%±0.7%, respectively), and group-1 (39.6%±2.8% and 14.3%±2.4%, respectively; Fig. 5C, D).
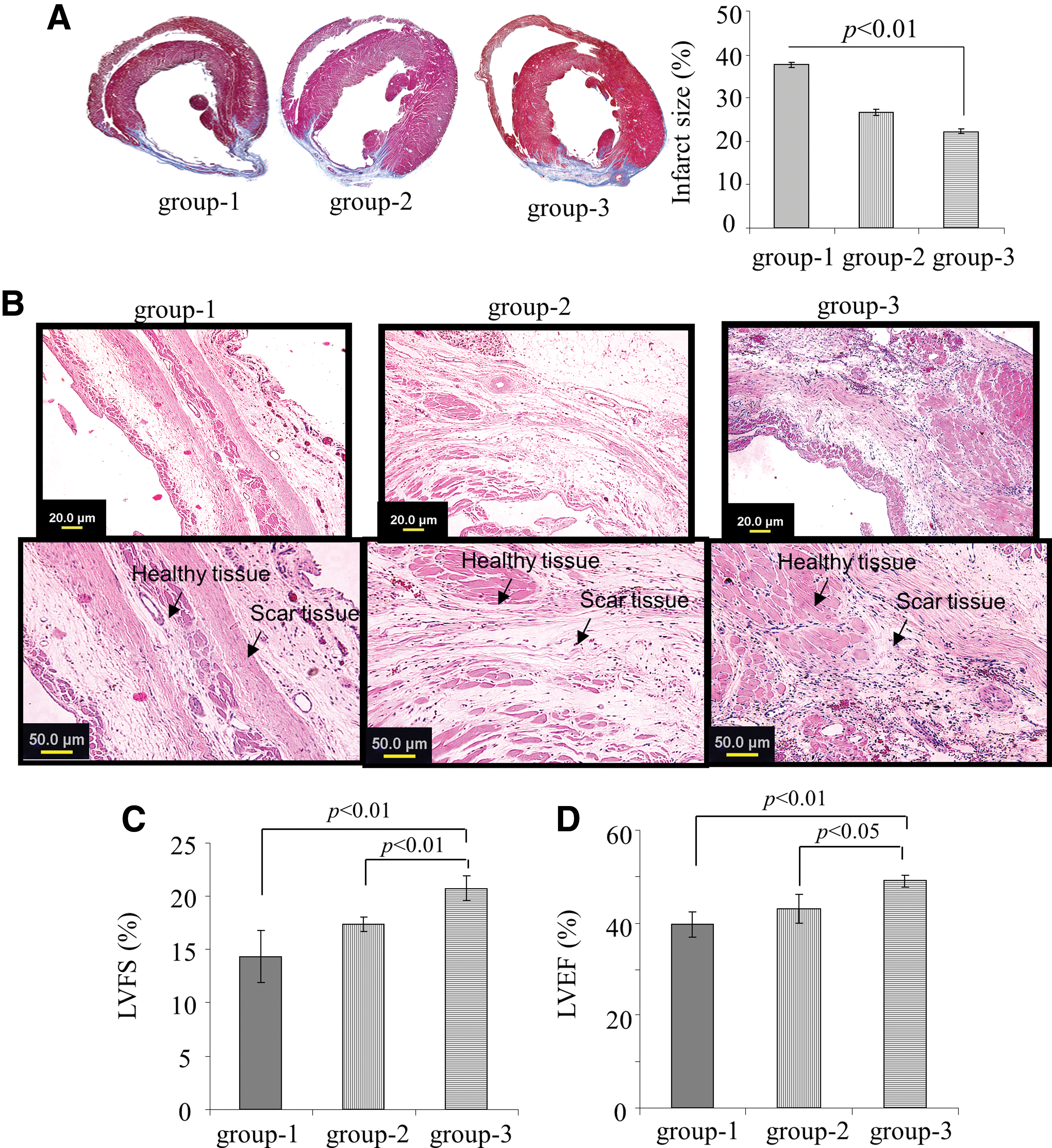
TransSM engraftment attenuated infarct size and improved global heart function.
TransSM transplantation and neomyoangiogenesis in the heart
Double fluorescence immunostaining of the histological tissue sections for GFP and desmin expression revealed increased myogenic differentiation of the transplanted SM in the infarct and peri-infarct regions in both cell transplanted groups-2 and -3 in the infarct and peri-infarct regions (Fig. 6A, B).
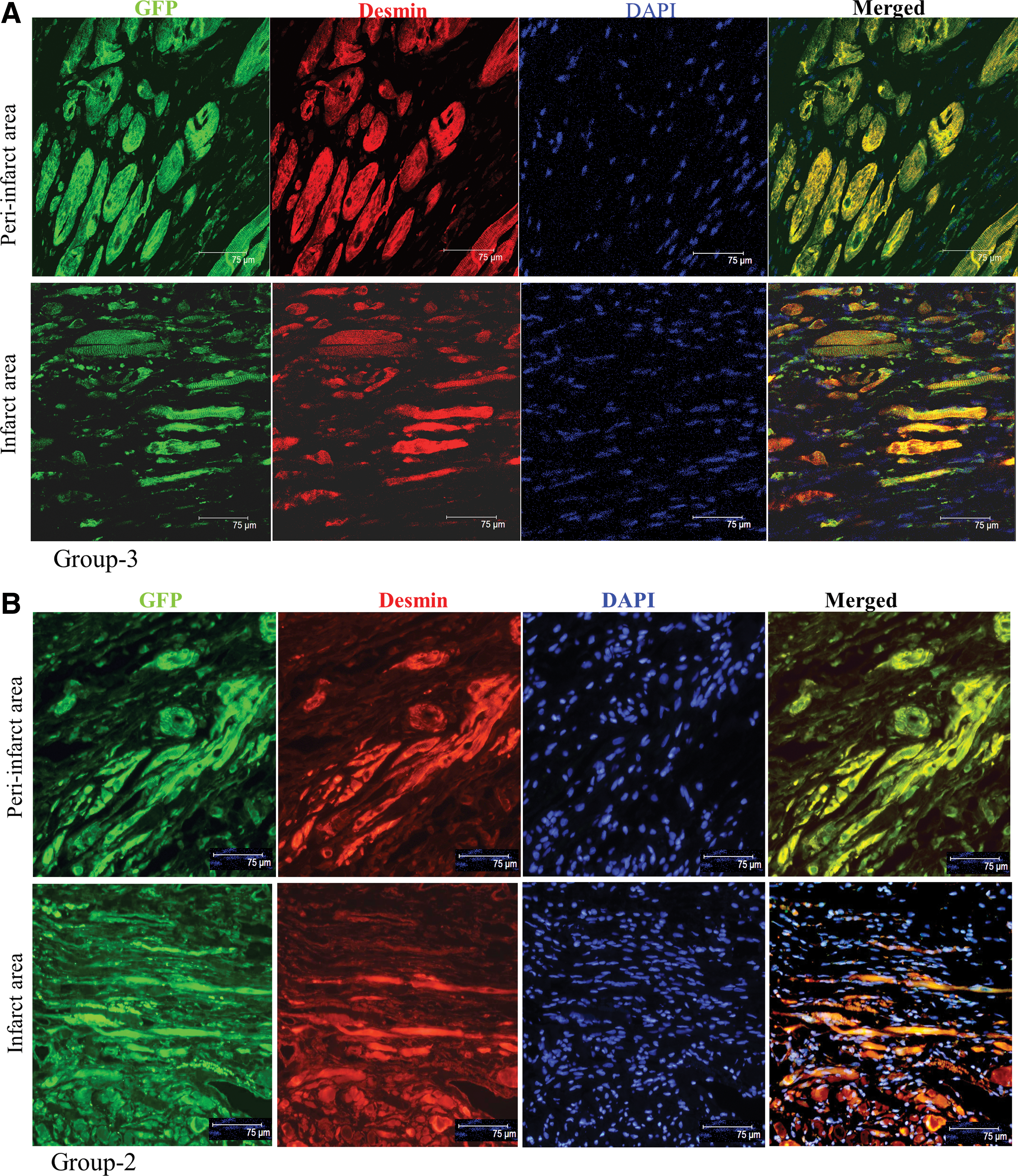
TransSM enhanced myogenesis in the infarcted myocardium. Double fluorescence immunostaining of histological tissue sections from:
Blood vessel density was measured in the center and peri-infarct regions after immunostaining of histological sections with the anti-vWFactor-VIII and antismooth muscle actin-specific antibodies (Supplementary Fig. S6). A large number of mature blood vessels were found in the areas surrounding the cell graft in group-2 and group-3. The blood vessel count per surface area (0.74 mm2) was highest in both peri-infarct (55.2±2.8) and infarct (31.5±1.9) regions in the TransSM transplanted group-3. This was more (P<0.01) than the peri-infarct (30.8±2.2) and infarct (21.3±1.4) regions in the DMEM group and (41.0±1.4) and (26.0±1.5) in the NatSM group. Although the number of blood vessels observed in the group-2 was higher as compared to the DMEM group-1, no significant difference (P>0.05) was observed between these groups either in the infarct or peri-infarct regions.
Discussion
The complexity of events occurring in the infarcted heart subsequent to ischemic injury necessitates a therapeutic strategy that could address various aspects of the disease process and minimize the severity of ischemic damage. Despite extensive research in stem cell therapy for myocardial repair during the past decade, search for finding a right donor cell type with ideal characteristics is still on. It is believed that reprogramming of stem cells to adapt to the harsh microenvironment of the ischemic myocardium for better survival and myogenic differentiation besides improved regional blood flow in the ischemic heart is an effective strategy [18 –20]. Our multipronged strategy explores the potential of stem/progenitor cell mobilization, survival, and angiomyogenic differentiation in response to the concentration gradient of the select group of cytokines having specific functions. Besides simultaneously recruiting the intrinsically available stem/progenitor cells in this study, release of growth factors from genetically reprogrammed SM altered the microenvironment of the ischemic heart via paracrine activity. These molecular events in response to SM-based overexpression of multiple growth factors led to additional therapeutic benefits in terms of attenuated infarct size and improved global heart function more than the transplantation of NatSM.
Given the low efficiency of nonviral method of transfection in general and multiple transfection protocol in particular, we opted to transfect the SM with individual transgenes. This was followed by pooling of the transfected cells that ensured their priming with each other's paracrine secretions before transplantation. Our multimodal nonviral gene therapy approach successfully accomplished activation and participation of the intrinsic ligand/receptor systems: VEGF/VEGFR, HGF/cMet, IGF-1/IGF-1R, and SDF/CXCR4, in the myocardial repair processes by creating a gradient of the respective ligands (growth factors) in the infarcted heart via transplantation of SM overexpressing these growth factors. We have previously shown the effectiveness of each of the 4 selected growth factors individually for treatment of experimental myocardial infarction (Supplementary refs. [6–9]). Although it is difficult to compare data from these studies with our present study results due to the difference in experimental design as well as difference in growth factor delivery strategies and the cell type used in transgene delivery, these data formed the basis of our multipronged strategy. Transgenic delivery of the individual growth factors showed effects specifically related to the particular growth factor and lacked the novelty of diverse signaling mechanisms achieved during the present study as a result of simultaneous delivery of multiple growth factors. Thus, activation of the multiple signaling mechanisms resulted in reduced cell apoptosis, increased cell proliferation, massive stem/progenitor cell mobilization, and homing to the site of ischemic injury for participation in the repair process through angiomyogenesis.
The rationale of selection of various growth factors was their functionally distinct role to mediate the process of myocardial repair. The selection was based on following criteria: (1) IGF-1 is a pleiotropic factor, which interacts with its widely distributed IGF-1R receptors to impart its cytoprotective, proproliferative, and prodifferentiation effects. We have recently shown that IGF-1 also promotes Cx-43 induction and translocation into mitochondria for its dual role of stem cell integration as well as protection postengraftment [18]. IGF-1 stimulation also results in HIF-1α induction via the Erk pathway, while hypoxia driven HIF-1α expression is independent of Erk [21]. However, high levels of HIF-1α protein expression is known to occur by stem cell factor stimulation under normoxic conditions in Akt- and ERK-dependent pathways [22]. (2) The interaction of VEGF ligand with tyrosine kinase receptors, VEGFR1 and VEGFR2, is proangiogenic and effectively promotes endothelial cell survival, proliferation, and growth with the activation of Akt signaling [23,24]. Although, hemangioma formation after VEGF gene delivery to the heart has been reported, the study used uncontrolled retroviral-based continuous VEGF expression [25] unlike our nonviral transient VEGF expression. (3) HGF is a known chemoattractant that induces stem cell migration in response to the concentration gradient developed in the infarct. HGF acts by providing a cue for the mesenchymal-epithelial transition factor (Met) expressing population of cardiac stem and progenitor cells [26]. HGF/Met ligand/receptor interaction regulates cell growth, cell motility, and morphogenesis by activating a tyrosine kinase signaling cascade [27]. Administration of HGF enhanced the efficacy of cell transplantation [28] and promoted cardiomyocyte proliferation in experimental animal models [29]. Furthermore, intramyocardial transplantation SM nonvirally modified for HGF expression has the capacity to limit infarct size expansion and cardiac dysfunction and these effects were accentuated by scaffold-based delivery of the cells [30]. (4) SDF-1α is a known ligand for CXCR4 receptors expressed on BMSC. The transient upregulation of SDF-1α in the infarcted heart during the acute phase after infarction provides a cue for mobilization and homing of the BMSC into the infarcted heart [31] besides acting as a retention factor to ensure their participation in the repair process [32]. Given their distinct functional role, we proposed the aggregated effect of the 4 growth factors to recruit and involve the intrinsically available stem/progenitor cell types in the repair process.
SM have been extensively studied for their ability to deliver therapeutic transgenes to the heart [33 –35]. Mostly, these studies involved conventional strategy of genetically modifying SM with 1 or 2 transgenes with proangiogenic and/or prosurvival effects and showed improved angiogenesis in the infarcted heart besides attenuation of infarct size. On the contrary, our novel strategy of multiple growth factor transgene delivery is aimed to harness their combined beneficial effects for treatment of infarcted heart. Unlike previous reports [33 –35], we provide a clear evidence that the select quartet of growth factors created a growth factor gradient for mobilization of recipients own pool of BMSC, EPC, and resident cardiac stem cells for participation in the repair process. Besides growth factor gradient, stem/progenitor cell mobilization and recruitment was also facilitated by paracrine release of MMP from TransSM [36]. MMP3 and MMP9 are implied in biogenesis of extracellular matrix and participation in tissue remodeling during various physiological and pathological processes [37]. Both VEGF and SDF-1α upregulate MMP-9 expression and cause shedding of KitL and recruitment of c-Kit+ BMSC [36]. Release of sKitL by MMP-9 enables the bone marrow repopulating cells to translocate to a permissive vascular niche favoring differentiation and reconstitution of stem/progenitor cell pool. We also observed a significantly higher expression of the Sfrp family Sfrp1, Sfrp2, Sfrp4, and Sfrp5. Sfrps belong to a family of diverse glycoproteins, which structurally resemble surface frizzled receptors and participate in activation of Wnt signaling, cellular differentiation, and cell death [38]. Upregulation of Wnts protein (such as Wnt3a) is observed in hypoxia and has been shown to induce apoptosis in cardiomyocytes through activation with its receptor frizzled [39]. It is currently thought that Sfrps, such as Sfrp2, compete with the frizzled receptor for Wnt ligands based on their structural resemblance, thereby preventing the activation of Wnt signaling, and hence exert antiapoptotic effects on cardiac myocytes and reduce fibrosis in the hearts [40,41]. Recently, Laeremans et al. [42] reported that a peptide (namely, UM206) blocked frizzled receptors resulting in reduced infarct size and improved functional recovery of the hearts. These results show that UM206 had the effect similar to Sfrps in terms of cardiac repair, which further support the rationale that the blocking of frizzled receptor (as is shown by the upregulation of Sfrps in our study) is effective in inhibiting Wnt3a and its detrimental effects on the myocardium.
More importantly, we observed higher Cx-43 in TransSM, which helped the cells in engraftment. In vitro studies showed that NatSM cocultured with neonatal cardiomyocytes expressed N-cadherin and Cx-43 at the cell–cell junctions [43]. However, most studies have reported lack of N-cadherin and Cx-43 expression by NatSM postengraftment and their failure to electromechanically couple with host cardiomyocytes despite a higher expression of both the proteins by nondifferentiated SM [44]. In our observation, TransSM expressed significant amounts of Cx-43 until day 7 after engraftment. However, we did not observe Cx-43 expression in 8 weeks histological sections.
Despite promising results, the study was not without limitations. Although there was significant increase in blood vessel density, the role and fate of the mobilized cells in angiomyogenesis in the infarcted heart at 8 weeks time point could not be determined due to lack of specific marker on the cells. The problem was further accentuated by difficulty in identifying GFP label on transplanted SM at 8 weeks, thus making it hard to distinguish neofibers originating from transplanted SM from those of mobilized cells. The inability of SM to undergo cardiogenic differentiation and electromechanically couple with host myocytes may warrant their replacement as a carrier with other cells having the potential to undergo cardiomyogenic differentiation. Furthermore, in vivo studies to assess the effectiveness of CM from transfected SM are also warranted. In conclusion, use of cytokines with SM for accelerated mobilization of stem/progenitor cells from peripheral circulation, bone marrow and the heart, and enhanced cell survival through secretion of paracrine factors is a novel approach for regeneration of the infarcted myocardium.
Footnotes
Acknowledgments
This work was supported by funds provided by the Department of Pathology and NIH grants No. R37-HL074272; HL080686; HL087246 (M.A.) and HL-087288; HL-089535 (Kh.H.H.).
Author Disclosure Statement
None declared.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.