
Abstract
We have previously identified Neighbor of Punc E 11 (Nope) as a specific cell surface marker of stem/progenitor cells in the murine fetal liver that is also expressed in hepatocellular carcinoma. Here, we focus on the differential expression pattern of Nope during murine fetal and postnatal liver development as well as in a normal and regenerating adult liver including oval cell activation. In the fetal liver, Nope shows a constantly high expression level and is a useful surface marker for the identification of Dlk, E-cadherin, and CD133-positive hepatoblasts by flow cytometry. Postnatally, Nope expression declines rapidly and remains barely detectable in the adult liver as shown by quantitative real-time reverse–transcriptase polymerase chain reaction and western blot analyses. Immunohistochemically, costainings for Nope- and epithelial-specific markers (E-cadherin), markers of early hepatoblasts (alpha-fetoprotein), and biliary marker proteins (CK19) demonstrate that Nope is initially expressed on bipotent hepatoblasts and persists thereafter on commited hepatocytic as well as cholangiocytic progenitor cells during late fetal liver development. Postnatally, Nope loses its circular expression pattern and is specifically directed to the sinusoidal membrane of early hepatocytes. While Nope is only weakly expressed on cholangiocytes in the normal adult liver, activated stem/progenitor (oval) cells clearly coexpress Nope together with the common markers A6, EpCAM, and CD24 in the 3,5-diethoxycarbonyl-1,4-dihydrocollidine mouse model. In conclusion, Nope should be most useful in future research to define the differentiation stage of hepatic-specified cells of various sources and is a promising candidate to identify and isolate hepatic stem cells from the adult liver.
Introduction
T
Murine bipotent hepatoblasts have been identified and isolated by the specific cell surface markers E-cadherin [15, 16], Liv2 [17], and Dlk [7]. Several hematopoietic cell surface markers, such as CD24 [16, 18], CD44 [19], CD133 [20], and Sca-1 [16], have also been identified to be expressed by these early hepatoblasts.
Interestingly, most of these markers are also expressed by bipotent oval cells in the adult liver that can differentiate into mature hepatocytes and cholangiocytes upon activation [21 –23], suggesting a direct lineage relationship with bipotent hepatoblasts. Oval cells can be identified in the injured adult liver if hepatocyte proliferation is blocked [24], and numerous liver injury studies in rodents suggest that these cells may represent a hepatic stem/progenitor cell compartment in the adult liver [25 –28].
In previous publications, we have introduced Nope as a new and specific cell surface marker of hepatoblasts within the fetal liver at ED12.5 using cDNA microarray studies [29] and—in a more recent project—we have shown that Nope is also expressed in murine and human hepatocellular carcinoma (HCC) [30]. However, there are no reports on the expression of the fetal stem/progenitor cell surface marker Nope on committed progenitor cells during late fetal/postnatal liver development or stem/progenitor cells in the adult liver.
In the present study, we focus on the differential expression pattern of Nope during murine fetal and postnatal liver development as well as in normal and regenerating adult liver, including the 3,5-diethoxycarbonyl-1,4-dihydro-collidine (DDC) mouse model of oval cell activation.
Materials and Methods
Animals
Pregnant C57Bl/6 mice were purchased from Janvier (Le Genest St Isle, France). All animal experimental procedures were conducted under protocols approved by the Animal Care Use Committee of the University of Cologne and were in accordance with the National Health and Medical Research Committee Guidelines.
DDC mouse model of oval cell activation
Mouse liver tissue from DDC-treated mice was obtained from V. M. Factor (NIH, Bethesda, MD). Briefly, the animals were fed a DDC diet at a concentration of 0.1% in a standard rodent chow. After 4 weeks, mice were sacrificed and liver samples were taken for immunohistochemical analyses [31].
Isolation of cells from the fetal liver
Fetal livers were microdissected at different stages of embryonic development starting at ED11.5. For further analysis, livers were dissected at day 2 (2 dpn) as well as 1 to 6 weeks postnatally (wpn) and from adult animals.
Single-cell suspensions of the fetal liver were obtained as described previously [16]. Briefly, fetal liver tissues were triturated in modified Hank's buffered saline solution (HBSS) containing 1 M ethyleneglycoltetraacetic acid and subsequently dissociated with modified HBSS containing 0.2% collagenase, 0.07% DNase, and 1 mM CaCl2. Viability was estimated by trypan blue.
Immunohistochemistry
Immunohistochemical detection was performed on cryosections of liver tissue fixed in ice-cold acetone. Rabbit anti-human Afp (Thermo-Fisher-Scientific), rat anti-mouse CK19 (TROMA, gift from R. Kemler, Freiburg, Germany), rat anti-mouse E-Cadherin (TaKaRa), rat anti-mouse A6 (generous gift from V. M. Factor), rat anti-mouse EpCAM (BD Bioscience), and goat anti-mouse Nope (R&D) primary antibodies were detected through Cy™2-conjugated donkey anti-rabbit (Jackson Immuno Research), APC-conjugated donkey anti-rat IgG (Jackson Immuno Research), or Cy™3-conjugated donkey anti-goat IgG. Dipeptidylpeptidase (DPP) IV was detected using a fluorescein isothiocyanate (FITC)-conjugated rat anti-mouse antibody (BD Bioscience) and CD24 was stained via an APC-conjugated rat anti-mouse antibody (Biolegend). IgG controls were used to control for background staining intensity (see Supplementary Fig. S1; Supplementary Data are available online at
RNA extraction
Total RNA was isolated using the Qiagen RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. RNA quantity was estimated using a Nano-Photometer (Implen) and 5 μg of total RNA were reverse transcribed using the Superscript III First Strand Kit (Invitrogen).
Quantitative real-time reverse transcriptase–polymerase chain reaction
Differential expression of selected genes was analyzed using quantitative real-time reverse transcriptase–polymerase chain reaction (qPCR). We performed a 2-step qPCR, using the SYBR Green Master Mix and the iQ5 Sequence Detection System (Bio-Rad Laboratories). For oligonucleotide primers see Supplementary Table S1. Amplification was as follows: 50°C for 2 min, 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, and 1 min at 60°C, completed by a dissociation curve to identify false-positive amplicons. Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was used as the reference gene. All samples were tested in triplicate and the relative expression level of each gene was calculated using the ΔΔct method.
Protein extraction and western blotting
Total proteins were extracted using the sodium dodecyl sulfate (SDS) lysis buffer containing 0.5% SDS, 15 mM Tris-HCl (pH 6.8), 2.5% glycerol, 1 mM ethylenediaminetetraacetic acid (EDTA), and proteinase inhibitor (Roche). The TED lysis buffer containing 1 mM EDTA, 1 mM dithiothreitol, and proteinase inhibitor in 50 mM Tris was used for extraction of membranous proteins. After SDS-polyacrylamide gel electrophoresis with equal amounts of protein (30 μg per lane), proteins were transferred onto nitrocellulose membranes (Whatman) using a semidry blot system (Bio-Rad Laboratories). The membrane was blocked overnight at 4°C in phosphate-buffered saline (PBS), pH 7.4, containing 5% dry milk powder. Primary antibodies, including goat anti-mouse Nope (R&D), rabbit anti-human Afp (Thermo-Fisher-Scientific), and mouse anti-mouse β-actin (Sigma-Aldrich), were applied for 2 h at 25°C. After extensive washing, membranes were incubated for 1 h at 25°C with peroxidase-conjugated anti-goat, anti-rabbit (Sigma-Aldrich), or anti-mouse IgG (Bio-Rad Laboratories). Immune complexes were detected using electrochemiluminescence advance western blot substrate (Amersham) according to the manufacturer's instructions. After detection, membranes were stripped using 0.2 N NaOH for 10 min before another primary antibody was applied.
Enzymatic deglycosylation
Total (SDS) or membrane (TED) proteins of fetal livers at ED14.5 were deglycosylated using Peptide N-Glycosidase F (PNGase F) (NEB) according to the manufacturer's instructions. Briefly, livers were lysed in SDS or TED lysis buffer. Ten micrograms of either SDS or TED protein was digested with 2000U PNGase-F for 2 h and subsequently analyzed by western blotting.
Flow cytometry
Isolated ED13.5 fetal liver cells were prepared for flow cytometric analysis. Briefly, 5×105 cells were incubated with the following primary antibodies: goat anti-mouse Nope (R&D), rat anti-mouse E-Cadherin (TaKaRa), rat anti-mouse Dlk (MBL), and FITC-conjugated rat anti-mouse CD133 antibodies (eBioscience). Unconjugated primary antibodies were detected through PE-conjugated donkey anti-goat and APC-conjugated donkey-anti-rat IgG, respectively (Jackson Immuno Research). Flow cytometric analysis was performed on a BD FACS Canto (BD Bioscience) equipped with BD FACSDiva software. 7-Amino-actinomycin-D (7-AAD) was used to exclude dead cells; debris was excluded based on physical parameters (forward and side scatter characteristics, respectively). The background fluorescence level of the cell suspensions was estimated using the appropriate IgG-control.
Results
Nope can be detected at the mRNA level until 2 weeks postnatally
We investigated the relative expression levels of Nope in livers using samples of fetal (ED11.5–ED18.5), postnatal (1 wpn to 3 wpn), and a normal adult mouse liver (AL) using qPCR. We validated that Nope is highly expressed in the fetal liver starting at ED11.5, showing a 20-fold higher level compared to the adult liver. Through fetal liver development, this expression level remains unchanged. At 2 wpn, the expression level is reduced to a 10-fold higher level compared to the normal adult liver and is further reduced to a 2-fold higher level at 3 wpn and barely detectable in the adult liver (Fig. 1).
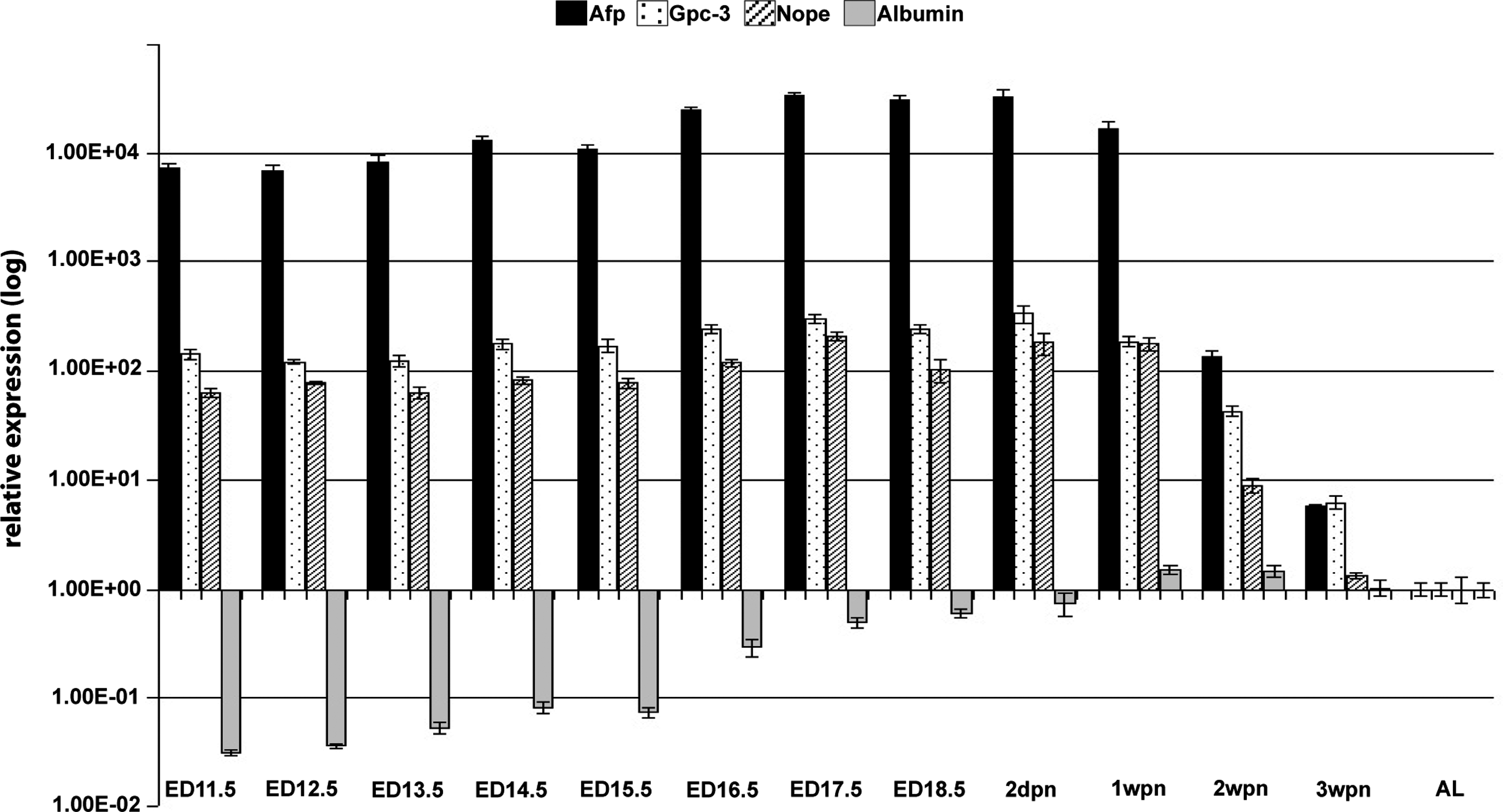
Neighbor of Punc E 11(Nope) expression during liver development. RNA was isolated from fetal (ED11.5 to ED18.5), postnatal (1 wpn to 3 wpn), and adult (AL) mouse livers, reverse transcribed into cDNA, and analyzed for the relative expression level of Nope (lines) compared to alpha-fetoprotein (Afp) (black), Gpc-3 (dotted), and albumin (gray). While Nope, Afp, and Gpc-3 show a similar expression pattern with high expression in the fetal liver and a rapid decline postnatally, the expression level of albumin as a marker of hepatocytic differentiation rises constantly during liver development until 1 wpn and remains at this level without significant changes thereafter. Expression levels were normalized to glyceraldehyde-3-phosphate dehydrogenase and calculated by the ΔΔct method and the relative mRNA expression is given as log 2-fold difference.
Afp as a marker of undifferentiated hepatocytes shows consistently high expression levels during all fetal stages of liver development starting at ED11.5 through 1 wpn with a 40-fold higher level compared to the normal adult mouse liver. At 2 wpn, Afp is dramatically downregulated, still showing a 20-fold higher level versus the adult liver. The expression is further reduced to a 10-fold higher level at 3 wpn, while it is barely traceable in adult liver tissue (Fig. 1).
Gpc-3, an oncofetal hepatic surface marker, is also highly expressed in the early stages of liver development, starting with a 20-fold higher level at ED11.5 compared to the normal adult liver. Parallel to the expression level of Afp, Gpc-3 swiftly declines at 2 wpn, revealing a 15-fold higher level versus the normal adult liver. Expression of Gpc-3 further declines to an 8-fold higher level at 3 wpn and is barely traceable in the adult liver (Fig. 1).
In contrast, the expression level of albumin as a marker of hepatocytic differentiation rises constantly during liver development until 1 wpn and remains at this level without significant changes thereafter (Fig. 1).
Nope expression at the protein level decreases postnatally
To determine the differential expression of Nope at protein level, we extracted total and membrane proteins of fetal, postnatal, and adult liver tissues and carried out western blot analysis. Nope protein is detectable at all stages of fetal liver development (ED14.5–ED18.5), displaying a strong band at 170 kDa and a faint band at 150 kDa in total protein extracts (Fig. 2A). However, if extracts of membrane proteins are analyzed, western blot analysis of Nope expression shows a clear band at 170 kDa as well as a strong wide band at 150 kDa (Fig. 2C). Postnatally, Nope expression is still detectable at 2 dpn and 1 wpn, but no longer upon 2 wpn (Fig. 2B, D). After deglycosylation, Nope antibody detects a single band in total protein lysate at 150 kDa, while analysis of membrane-enriched protein lysate displays 2 bands referring to 150 and 120 kDa (Fig. 2E).

Western blot analysis of Nope during liver development. Total
The expression pattern of Afp in total protein fractions was comparable to Nope, demonstrating strong bands with a molecular weight of 70 kDa through all stages of fetal liver development, but decreasing until 1 wpn (Fig. 2A, B). Two weeks after birth, expression of Afp was no longer detectable (Fig. 2B). β-Actin was used as an internal loading control (43 kDa).
Nope as a surface marker of bipotential hepatoblasts and committed progenitor cells
The expression of Nope in the developing liver was further studied by immunohistochemical costainings, analyzing the hepatocytic and cholangiocytic lineage commitment. In the early stages of liver development (ED14.5), Nope expression is restricted to hepatoblasts within the fetal liver as identified by coexpression with Afp and E-cadherin (Fig. 3A–D). At ED17.5, when hepatoblasts have differentiated into committed progenitor cells, Nope-expressing cells are still positive for Afp and/or E-cadherin (Fig. 3E–H). While Afp staining declines with ongoing liver development and is no longer traceable at 1 wpn, Nope continues to be expressed on early E-cadherin-positive hepatocytes (Fig. 3I–L). Hepatocytes in the adult liver show no significant staining for Nope or Afp, while E-cadherin is detectable only on hepatocytes located in the periportal area (Fig. 3M–P).
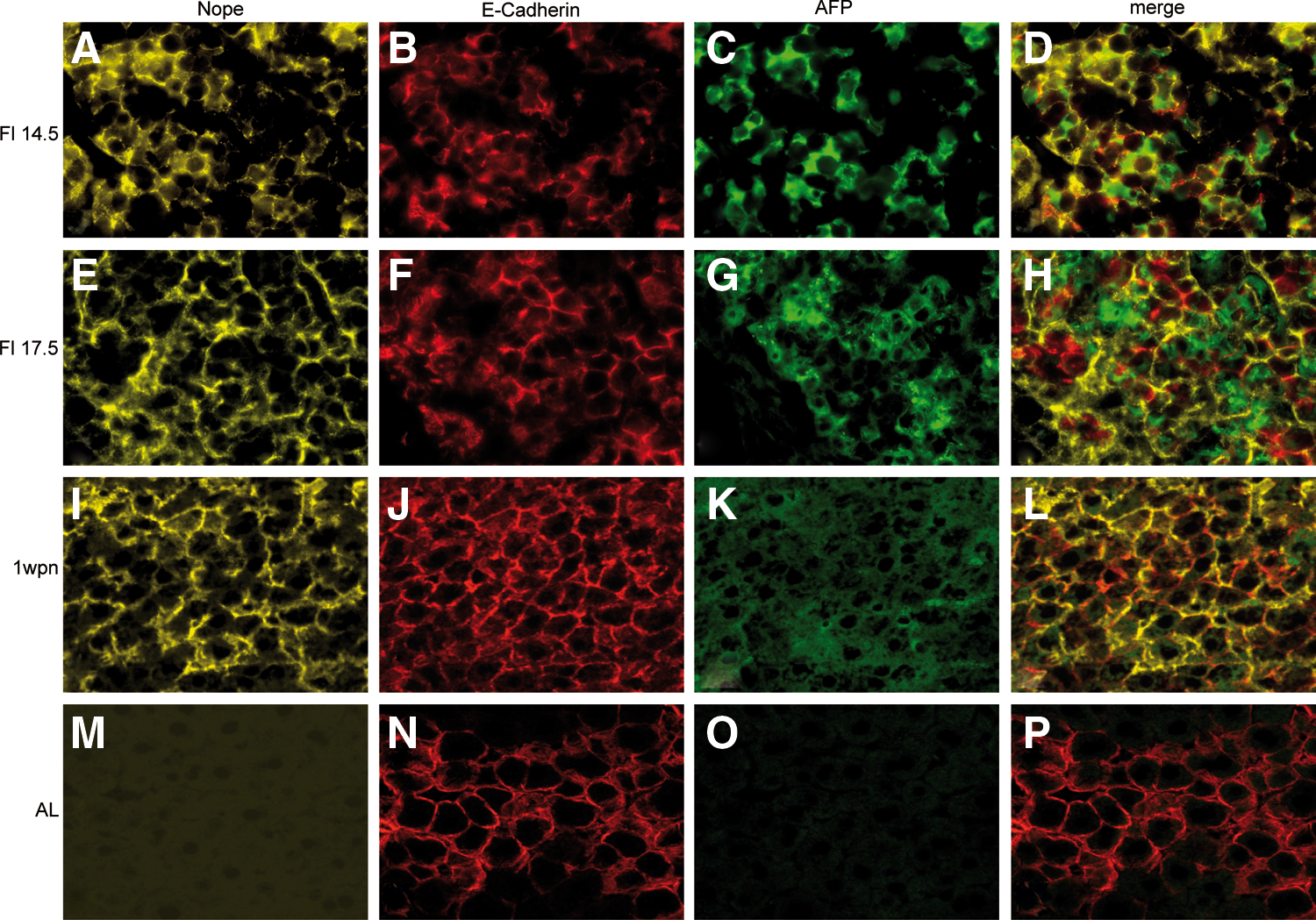
Immunohistochemistry of Nope, Afp, and E-cadherin during liver development. Cryosections of early fetal
Combined stainings with Nope, Afp, and the cholangiocyte marker CK19 demonstrated that CK19 is not detectable in bipotential hepatoblasts at ED14.5 (Fig. 4A–D). After differentiation into committed progenitor cells at ED17.5, Nope expression is not restricted to Afp-positive hepatocytic progenitor cells, but is also clearly detectable on early bile ducts, that is, committed cholangiocytic progenitor cells (Fig. 4E–H). Postnatally and in the adult liver, Nope expression continues to be present on early and differentiated cholangiocytes (Fig. 4I–P). IgG controls were used to control for background staining intensity (see Supplementary Fig. S1).

Immunohistochemistry of Nope, Afp, and CK19 during liver development. Cryosections of early fetal
Membranous localization of Nope on early hepatocytes
The localization of Nope on the plasma membrane was further studied by costainings of different liver developmental stages with DPPIV. DPPIV is not detectable before ED17.5 and shows a circular staining pattern at ED18.5 (Fig. 5B). At 2 dpn, DPPIV staining becomes more distinct on the canalicular membrane of early hepatocytes (Fig. 5F) and is a specific marker for the canalicular membrane thereafter (Fig. 5J).

Membranous localization of Nope during liver development. Cryosections of late fetal
Interestingly, the membranous expression pattern of Nope traverses similar modifications during liver development. While Nope shows a circular staining pattern of the whole plasma membrane of hepatoblasts in the fetal liver until ED18.5 (Fig. 5A), localization of Nope is specifically directed to the sinusoidal membrane of early hepatocytes postnatally (Fig. 5E, I).
The merged pictures show the overlapping expression of Nope and DPPIV in the fetal liver (Fig. 5D) that becomes more separate at 2 dpn (Fig. 5H) and is nicely complementing each other with DPPIV on the canalicular and Nope on the sinusoidal membrane of early hepatocytes at 1 wpn (Fig. 5L).
Fluorescence-activated cell-sorting analysis of bipotent hepatoblasts for Nope and other surface markers
Fluorescence-activated cell sorting (FACS) analysis was performed to show coexpression of Nope with other surface markers of bipotent hepatoblasts. At ED13.5, 2% of the total cell suspension stain positive for the epithelial-specific marker E-cadherin (Fig. 6A). Costaining with Nope reveals a total overlap with the fraction of E-cadherin-positive cells (Fig. 6B). Likewise, the combined staining with Dlk demonstrates that all Dlk-positive cells (2%) are also positive for Nope (Fig. 6C). However, the surface marker CD133, positive in 2.9% of the dissociated liver cells, displays an overlap of double-positive cells for Nope and CD133 in 1.6% of the total cell suspension, while an additional single-positive subpopulation of CD133 remains (Fig. 6D).
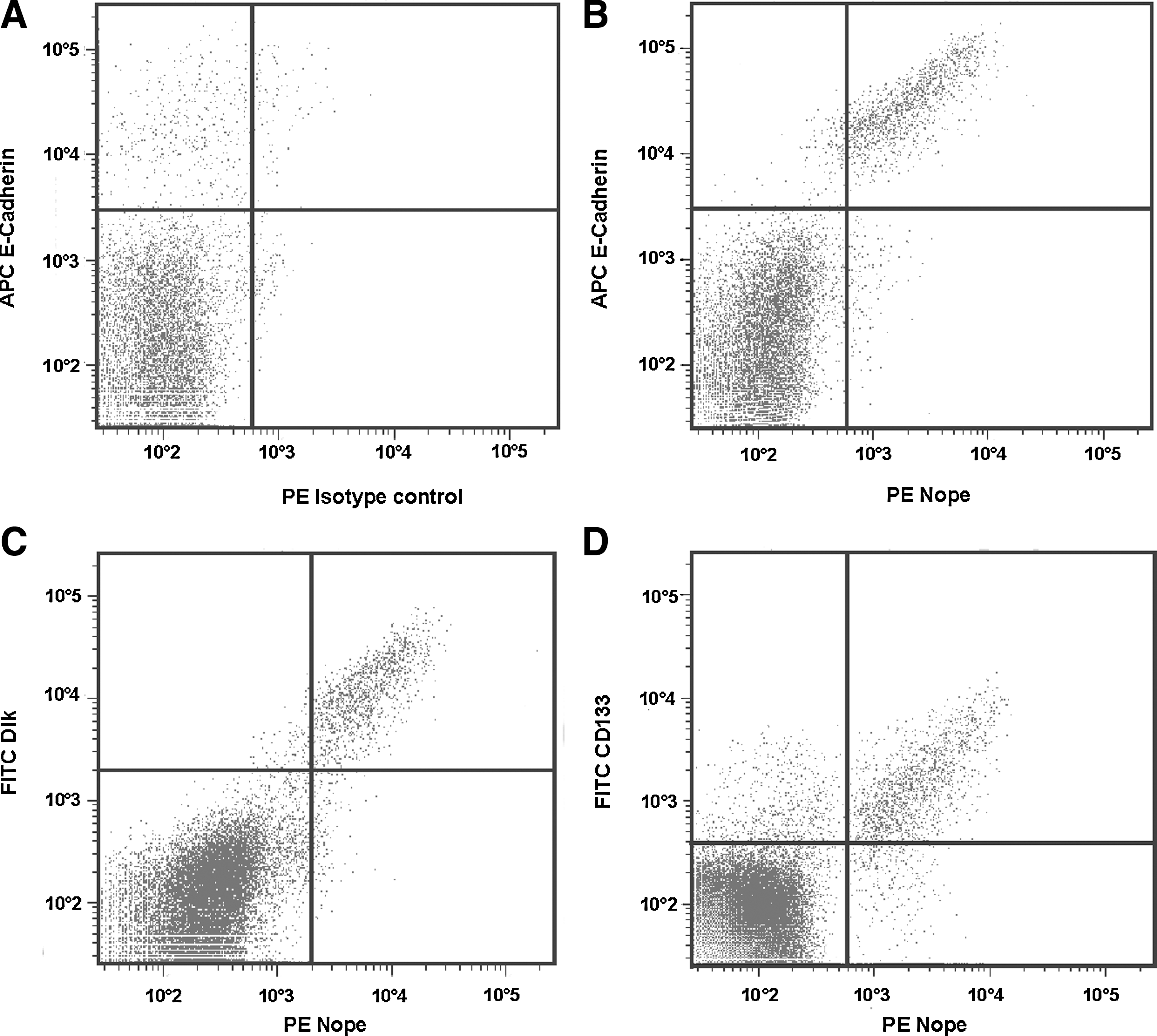
Fluorescence-activated cell-sorting analysis of Nope+ hepatoblasts for other cell surface markers. Dissociated fetal liver cells at ED13.5 were stained with an isotype control
Nope expression on activated oval cells in the DDC mouse model
In the DDC mouse model of oval cell activation, Nope clearly identifies not only bile ducts, but also activated oval cells radiating from the periportal region and forming primitive ductular structures that are characterized by their typical morphological appearance (small in size, ovoid nucleus, and scant basophilic cytoplasm) (Fig. 7A, E, I). Staining with the common markers of murine oval cells A6 (Fig. 7B), CD24 (Fig. 7F), and EpCAM (Fig. 7J) confirms coexpression with Nope on ductular and oval cells (Fig. 7D, H, L). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (Fig. 7C, G, K).

Detection of adult hepatic stem/progenitor (oval) cells by Nope. Cryosections of DDC mice livers were costained for Nope
Discussion
The first report on a gene named Neighbor of Punc E11 (Nope) was by Salbaum and Kappen in 2000 [32]. By Northern blot analysis, they reported the expression of Nope in the brain, skeletal muscle, and heart of adult mice. In addition, using in vitro hybridization, they demonstrated Nope to be expressed in the developing mouse embryo, preferentially in developing muscle tissues and the nervous system.
We show that Nope is a glycosylated protein with 2 isoforms of different sizes, representing a soluble, cytoplasmic isoform of 170 kDa and a membrane-bound isoform of 150 kDa. It is current knowledge that Nope is a member of the immunoglobulin superfamily and is thought to be essential in axon guidance by similarity with its family members neogenin and deleted in colorectal cancer (Dcc) [32]. Dcc and neogenin are netrin receptors that are expressed during transition of undifferentiated into differentiated cell types, preferentially expressed by differentiating neurons in the central nervous system [33]. Neogenin is thought to act in netrin receptor-adhesive clustering and it may be involved in maintenance of multipotent progenitor cells [34]. However, we could not confirm any binding of netrins to Nope (M. Koch, unpublished data).
In our present study, we provide evidence on mRNA and protein level that Nope is constantly expressed in the fetal liver starting at ED11.5, but is rapidly downregulated postnatally and remains barely detectable in the adult liver. Functional implications of Nope in the developing liver remain speculative, but could be in homotypic cell–cell contact, guiding the formation of hepatic cords or being involved in signal transduction as a protein in the plasma membrane by coupling an extracellular ligand to second-messenger cascades inside the cell.
Our immunohistochemical studies demonstrate that Nope is initially expressed on hepatoblasts and thereafter persists on committed hepatocytic as well as cholangiocytic progenitor cells during late fetal liver development. Postnatally, Nope loses its circular expression pattern and is specifically directed to the sinusoidal membrane of early hepatocytes. When hepatocytes have reached terminal differentiation in the adult liver, Nope is no longer detectable while it remains only weakly expressed on cholangiocytes. However, in the DDC mouse model of oval cell activation, Nope clearly identifies oval cells radiating from the periportal region and forming primitive ductular structures. Oval cells can be identified in the injured adult liver if hepatocyte proliferation is blocked [24] and numerous liver injury studies in rodents suggest that these cells may represent a hepatic stem/progenitor cell compartment in the adult liver [25 –28].
In addition, oval cells are also considered to be a cellular precursor for HCC [35]. In a previous study, we have already shown that Nope is also a surface marker of murine and human HCC comparable to the established oncofetal marker proteins Afp, a cytoplasmic glycoprotein, and Gpc-3, a membrane protein that belongs to the family of heparan sulfate proteoglycans [30, 36 –38]. The similar expression profile of Nope and these markers during liver development with a strong expression in the fetal liver, rapid decrease after birth, and minimal or no expression in the normal adult liver suggests that regulation of these markers underlies similar pathways during normal liver development. Comparable to the expression of Nope on stem/progenitor (oval) cells in the DDC mouse model, Afp becomes expressed on activated oval cells after an experimental liver injury using the 2-AAF/PH rat model [21]. Grozdanov et al. established Gpc-3 to be differentially expressed in proliferating oval cells in different models of rat liver injury [39].
Similar expression patterns have been described for other hepatic stem/progenitor cell markers like Dlk, EpCAM, and CD133: Dlk is expressed in the murine fetal liver from ED10.5 to ED18.5 and is not detectable after birth or in the adult liver [7]. Dlk-positive fetal liver cells are large hepatic cells expressing albumin and are able to differentiate into hepatocytes as well as biliary epithelial cells [7]. We have previously demonstrated by FACS analysis that expression of Dlk is specific for Afp-positive hepatoblasts within the fetal liver [16]. In the present study, we show by FACS analysis that bipotent hepatoblasts identified by Dlk are likewise positive for Nope. While Dlk is not expressed in liver regeneration after partial hepatectomy [40], Dlk has been described as a marker for a subset of oval cells in hepatocytic differentiation in 2-AAF/PH and retrorsine/PH rat models [22, 40]. Also, Yanai et al. reported that Dlk is expressed on 20.5% of human HCC, where it was more frequently found in HCC of younger patients [41].
EpCAM is a surface marker of hepatoblasts that is expressed from the onset of liver organogenesis to approximately ED14.5 [42]. After lineage commitment, EpCAM expression is restricted to committed cholangiocytes as well as mature cholangiocytes [43]. Moreover, previous studies identified EpCAM as a surface marker that is expressed on mouse and rat oval cells [23, 44], and we could demonstrate in our study that murine oval cells coexpressed EpCAM and Nope. Recently, EpCAM has been proposed to be a potential marker of liver cancer stem cells [45].
Prominin (CD133) is a membrane glycoprotein that is found in neuroepithelium, but also in various other epithelia of the mouse embryo [46]. In the adult mouse, prominin has been detected in the brain ependymal layer and in the kidney tubules. In a very recent study, Kakinuma et al. reported CD133 as a positive selection marker for bipotent hepatoblasts in the mouse [20]. In our study, we confirmed the expression of Nope and CD133 on bipotent hepatoblasts using FACS analysis, but describe an additional subpopulation that expresses CD133 alone and probably represents hematopoietic stem cells. In 2007, Yovchev et al. had already described the expression of CD133 on the surface of activated oval cells in the 2-AAF/PH rat model [23]. In our study, however, murine oval cells showed no positive staining for CD133. Finally, CD133 has been frequently used as a marker of liver cancer stem cells in cell lines [47] and HCC [48, 49].
Overlapping expression on hepatic stem/progenitor cells and hematopoietic cell populations has also been described for A6 and CD24. Nevertheless, both markers have been useful in our study to show coexpression with Nope on activated murine oval cells: The A6 antibody, previously described by V. Factor and colleagues, detects biliary epithelial cells of all ducts, including canals of Hering, in the normal adult liver. In the fetal liver, however, A6 expression is not restricted to hepatoblasts, showing weak signals between ED12 and 15, but is also found on hematopoietic cell fractions [50, 51]. In mouse models of oval cell activation, A6 is known to recognize not only bile ducts and ductules, but also proliferating oval cells [52].
CD24 is a surface sialoglycoprotein that is mainly expressed on cells of hematopoietic origin, including lymphocytes, granulocytes, thymocytes, monocytes, erythrocytes, and dendritic cells. While we and others detected expression of CD24 on bipotent hepatoblasts [18, 23, 29], it is also described on biliary epithelial cells [53], on oval cells activated in the 2-AAF/PH rat model [23] and in DDC-treated mouse livers [53]. Interestingly, the later one was the first study to show the isolation of a progenitor cell population from the untreated adult mouse liver [18]. CD24 was also described to be a marker of functional liver tumor-initiating cells [54] and might be useful to predict poor prognosis [55].
The high number of common markers of fetal and adult hepatic stem/progenitor cells led to the speculation that oval cells may be a subset of undifferentiated fetal liver cells that remain in adulthood. However, a precise lineage relationship between epithelial cells in the fetal liver and oval cells in the adult liver has not yet been established. Among all the presented markers, only EpCAM, A6, and CD24 had already been demonstrated to identify oval cells in the mouse model. Among these, A6 and CD24 are not specific for hepatoblasts within the fetal liver, but also stain positive multiple hematopoietic cell fractions. Therefore, only EpCAM and Nope as a newly identified surface marker of murine adult hepatic stem/progenitor (oval) cells might be especially valuable markers to follow on this way.
In conclusion, Nope is a new surface marker of fetal and adult hepatic stem/progenitor cells. Due to its differential expression and staining pattern during liver development, Nope will be most useful in future research to define the differentiation stage of hepatic-specified cells of various sources in vitro and in vivo. Its coexpression with common stem/progenitor cell markers clearly suggests that Nope is a promising candidate to identify, characterize, and isolate hepatic stem cells from the adult liver.
Footnotes
Acknowledgments
We are grateful to G. Holz and E. Konze for their excellent technical assistance. We want to thank V.M. Factor, M. Kitade and J. Marquardt for their invaluable support to get liver tissue from DDC mice and V.M. Factor for the generous gift of A6 antibody.
This project was supported by a grant from the DFG (Deutsche Forschungsgemeinschaft) to D.N.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.