
Abstract
Metabotropic glutamate receptors (mGluRs) are G-protein coupled receptors (GPCRs) activated by glutamate. The function of mGluRs is not restricted to the regulation of synaptic transmission. Although some roles of mGluR5 in mouse embryonic stem cells (ESCs) have been proposed, little is known about the significance of mGluR5 in cardiomyocyte differentiation from ESCs. We demonstrated that mGluR5 expression increased during cardiomyocyte differentiation. Activation of mGluR5 with (RS)-3, 5-dihydroxy phenylglycine (DHPG) promoted cardiomyocyte differentiation in a dose-dependent manner. DHPG significantly enhanced PI 3-kinase enhancer (PIKE) and PI3K p110α expression, but had no significant effect on Homer1b/c. The coexpression of PIKE or PI3K p110α together with Troponin T in embryoid bodies (EBs) treated with DHPG was elevated to 9.51% and 12.05%, respectively. Inhibition of mGluR5 with 2-methyl-6-(phenylethynyl)pyridine (MPEP) treating the ESCs, did hold back the cardiogenesis from the ESCs at the early differentiation stage. However, EBs applied by MPEP could not inhibit cardiomyocyte differentiation. Small interfering RNA (siRNA) of mGluR5 blocked cardiomyocyte differentiation by repressing PIKE and PI3K p110α expression, but had no notable influence on Homer1b/c. mGluR5 siRNA also decreased the DHPG-induced Ca2+ transient peak amplitude in the isolated ESC-derived cardiomyocytes. The amplitude of Ca2+ oscillation was reduced by ∼90% with si-mGluR5-3 compared with si-control. The protein expression of T-type Ca2+ channel and L-type Ca2+ channel was decreased in si-mGluR5-3–treated EBs. Taken together, these results revealed that mGluR5/PIKE/PI3K signaling pathway was involved in cardiomyocyte differentiation from ESCs. The key function of mGluR5 is probably associated with cardiogenesis and Ca2+ signal in ESC-derived cardiomyocytes.
Introduction
M
mGluR5 is present in the hearts of rats and is preferentially located in nerve terminals, ganglion cells, and elements of the nervous system. Immunohistochemistry analysis suggests that mGluR5 is more predominant in the ventricles of the heart. Moreover, mGluR5 is the only mGluR subtype located in the intercalated disks (IDs) of the cardiac muscle and in the endothelial lining of blood vessels. IDs are specialized in various ways for maintaining cell cohesion and providing low resistance bridges for the spread of excitation. Preferential localization of mGluR5 may allude to the specific role in cardiac function [3]. However, further studies are required to elucidate the role of mGluR5 during the differentiation of the embryonic heart. This may reveal the cross-talk mechanism related to mGluR5 in heart development.
Homer contains a single PDZ (the postsynaptic protein PSD-95/SAP90)-like domain and specifically binds to the carboxyl terminus of phosphoinositide-linked mGluRs. Homer1 proteins regulate synaptic cycling and Ca2+ signaling of mGluR5 [5]. PIKE (PI 3-kinase enhancer) is a recently identified brain specific GTPase, which binds PI 3-kinase (PI3K) and stimulates the lipid kinase activity. The Homer/PIKE complex couples PI3K to mGluR5 and regulates a major action of mGluR5, prevention of neuronal apoptosis [6]. Due to mGluR5 existence in adult cardiomyocytes, we hypothesized that the mGluR5/Homer/PIKE/PI3K signaling pathway may also function in cardiomyocyte differentiation.
In the present study, cardiomyocyte differentiation from embryonic stem cells (ESCs) in vitro, which faithfully recapitulates the process in vivo, was employed to evaluate the expression and function of mGluR5 during cardiomyocyte differentiation. mGluR5 is the only mGluR subtype expressed in undifferentiated ESCs. Activation of mGluR5 supported ESCs self-renewal [4]. So far, little evidence deals with mGluR5 in the ESC-derived cardiomyocyte system. The precise role of mGluR5 in cardiomyocyte differentiation remains to be elucidated, although the expression and function of mGluR5 in GABAergic neuron differentiation from ESCs has been reported [7]. Presently, we identified whether the mGluR5/Homer/PIKE/PI3K signaling pathway was involved in cardiomyocyte differentiation from ESCs. Either mGluR5 activator (RS)-3, 5-dihydroxyl- phenylglycine (DHPG), or inhibitor 2-methyl-6-(phenylethynyl) pyridine (MPEP) was used to evaluate the function of mGluR5 during cardiomyocyte differentiation, as determined by western blot and flow cytometric analysis. In addition, small interfering RNA (siRNA) of mGluR5 was applied to investigate the function of mGluR5 associated with Ca2+ oscillations and Ca2+ channel protein.
Materials and Methods
Materials
The permanent ES cell line D3 was obtained from the American Type Culture Collection (CRL-1934) [8]. Institute of Cancer Research (ICR) mice were obtained from the Experimental Animal Center, Zhejiang University, Hangzhou, China (Grade 2, Certificate No. 2007-0029).
Cell culture and treatment
ESCs were differentiated into cardiomyocytes as described previously [9]. In brief, cultures of differentiating mouse ESCs were established by forming embryoid bodies (EBs) in hanging drop culture. Thirty-milliliter drops containing approximately 600 ESCs were placed on the lids of petri dishes filled with D-Hanks solution and cultured as inverted hanging drops for 3 days. Then the EBs were transferred to float in the petri dishes for an additional 2 days. On day 5, the EBs were plated separately onto gelatin-coated (0.1%) 24-well plates. At this time (day 5), retinoic acid (RA; Sigma) or dimethyl sulfoxide (DMSO; Sigma) solvent was added to the differentiation medium to be used as positive and negative controls, respectively. EBs were treated from day 5 with mGluR5 agonist DHPG (Tocris Bioscience). mGluR5 inhibitor MPEP (Tocris Bioscience) was applied to ESCs or day 5 EBs. In this experiment, day 5+× referred to day×after the EBs were plated onto gelatin-coated plates. Rhythmically beating EBs were considered to be spontaneously beating cardiomyocytes in EB outgrowths and were defined as the phenotypic marker of successful differentiation.
Assessment of cytotoxicity
To determine DHPG or MPEP cytotoxic effect on ESCs, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) tests were performed in the absence of leucocyte inhibitory factor (LIF). Five hundred cells were seeded into each well of a 96-well microtiter plate and grown in the presence of 10−5, 10−6, and 10−7 mol/L DHPG or MPEP. A negative control containing solvent diluted in medium was also included. After 10 days of culture with two changes of medium (containing the appropriate concentration of DHPG or MPEP) on days 3 and 5, the viability of the cells was determined using MTT test.
Fetal mouse heart obtainment
ICR mice (males weighing 24±2 g and females weighing 22±2 g) were housed under a 12-hour light/dark cycle and 21±1°C temperature condition. To obtain fetuses, mice (3 female and 1 male) were housed together. The day that vaginal sperm or a copulation plug was observed was defined as day 0 of gestation. Mouse hearts were obtained at embryonic days E16, 18, and 20 and from newborns (N) and adults (A).
Immunofluorescence analysis
Immunofluorescence analysis was done as described previously [9]. In brief, cultures were fixed for 10 min in ice-cold methanol. Then, the fixed cultures were permeabilized by incubation with 0.5% triton X-100 in phosphate-buffered saline (PBS) for 30 min. After treatment with cattle serum for 30 min, specimens were incubated at 4°C overnight together with the appropriate primary antibodies: anti-α-actinin (1:200; Sigma), anti-mGluR5 (Epitomics), anti-Troponin T, anti-Homer1b/c, anti-PIKE, and anti-PI3K p110α (Santa Cruz Biotechnology) (1:50). Specimens were then incubated with the respective fluorescent secondary antibodies. DAPI (4, 6-diamidino- 2-phenylindole; Sigma) was used to dye the cell nuclei. Cells were then observed under a fluorescence microscope (Leica DMIL).
Semi-quantitative reverse transcription-polymerase chain reactions
Total RNA was isolated from ESCs, EBs, and cardiomyocytes derived from EBs on day 5+3, 5+7, and 5+11, and from mouse heart tissues at various developmental stages using Trizol reagent (Gibco BRL) in accordance with the manufacturer's instructions. Reverse transcription-polymerase chain reactions (RT-PCR) were done as described previously [9]. The sense and antisense primers were as follows Table 1. cDNA synthesis was performed using 3μg RNA with MMLV RT (Invitrogen). PCR products were analyzed by 1.5% agarose gel electrophoresis, visualized with ethidium bromide staining, and then quantified using a bio-imaging analyzer (Bio-Rad). The density of the products was quantitated using Quantity One version 4.2.2 software (Bio-Rad).
Western blot
Western blot was done as described previously [9]. In brief, cells and tissues were collected in sodium chloride 150 mM, NP-40 1%, Tris, pH 8.0 50 mM, sodium deoxycholate 0.5%, SDS 0.1% (RIPA) buffer and lysed for 30 min on ice. An 80-μg aliquot of the supernatant protein from each sample was separated electrophoretically on a 7.5% sodium dodecyl sulfate–polyacrylamide gel. Subsequently, proteins were transferred onto 0.45-μm pore size PVDF membranes for 90 min and blocked for 1 h, followed by an overnight incubation at 4°C with the appropriate antibodies: anti-mGluR5 (1:5,000), anti-Troponin T, anti-Homer1b/c, anti-PIKE, anti-PI3K p110α (1:500), and anti-GAPDH (1:10,000). The membranes were then incubated with horseradish peroxidase (HRP)-conjugated antibodies (Affinity Bioreagents) (1:5,000). The proteins were visualized autoradiographically with an enhanced chemiluminescent substrate (ECL; Pierce), and scanned using a bio-imaging analyzer (Bio-Rad). The product densities were quantitated using Quantity One version 4.2.2 software (Bio-Rad).
Flow cytometry
EBs were harvested at day 5+11 and digested into single cells with 1 mg/mL collagenase 2 (Worthington Biochemical) before immunostaining. Cells were then fixed in 4% paraformaldehyde for 60 min. After treatment with 3% bovine serum albumin for 1 h, cells were incubated at 4°C overnight together with the appropriate primary antibodies: anti-mGluR5 (1:100), anti-Troponin T, anti-Homer1b/c, and anti-PIKE (1:100). Cells were then incubated with the secondary antibodies. A total of 1×104 events were routinely collected with the flow cytometer (Becton-Dickson). Differentiation was determined by comparing the fluorescence intensity of the treated cells with that of the untreated cells obtained from a DMSO solvent control plate.
Isolation of ESC-derived cardiomyocytes
EBs containing cardiomyocytes were detached from culture surfaces by incubation with 0.05% trypsin-thylenediaminetetraacetic acid for 1 min at 37°C. The cells were then dissociated with 1 mg/ml collagenase, resuspended in differentiation medium, and loaded onto a discontinuous percoll gradient. Percoll (Pharmacia) was diluted in a buffer containing 20 mmol/L 4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) and 150 mmol/L NaCl. The gradient consisted of a 40.5% percoll layer over a layer of 58.5% percoll. Cell layers were apparent after centrifugation (1,500 g) for 30 min. Cells in different layers were collected, washed, resuspended in the differentiation medium and plated for siRNA transfection.
siRNA transfection
The siRNAs targeting mouse mGluR5 mRNA and a validated negative control siRNA labeled with Alexa-488 were ordered from Qiagen as follows:
mGluR5-1: TACTGTAATCTTCATCATTTA, mGluR5-2: CTCCAGGAAGGTATGAAATAA, mGluR5-3: CAGGAGT GGATTAAGCAGTTA, and siRNA-negative control: AAT TCTCCGAA CGTGTCACGT. Briefly, the siRNA-negative control, mGluR5-1, -2, or -3 (200 pmol) were combined with 2 μL Lipofectamine-2,000 in a final volume of 200 μL in Opti-MEM medium (Invitrogen). The lipofection mixture was added to freshly passaged ESCs in suspension (15×104 cells/well of a 12-well plate). Twenty-four hours after transfection, the cells were harvested for EB formation. The lipofection mixture was also added to an isolation of ESC-derived cardiomyocytes. Twenty-four hours after transfection, the medium was changed to normal culture medium. Another 24 hours later, the cells were harvested for western blot analysis or measurement of intracellular Ca2+ concentration [Ca2+]i.
Measurement of [Ca2+]i
The isolated ESC-derived cardiomyocytes were cultured in glass-bottom culture dishes. Following siRNA transfection, the cells were washed twice with PBS and loaded with 5 μmol/L fluo4-acetoxymethyl ester (fluo4-AM) in KrEB/Henseleit buffer for 30 min at 37°C in the dark. Cells were then washed twice with KrEB/Henseleit buffer and maintained at 37°C in a temperature controlled perfusion chamber (PerkinElmer, and placed on the stage of an inverted microscope (Nikon). To monitor intracellular Ca2+ oscillations, 100 μmol/L DHPG was added to the cells through the perfusion chamber.
Statistical analysis
Data are expressed as mean values with standard deviation. At least 3 independent experiments were performed. Statistical analysis was performed with one-way analysis of variance. P values of P<0.05 and P<0.01 were considered to be significant.
Results
mGluR5 expression in cardiomyocytes differentiated from ESCs in vitro
Apparent morphological cell changes have been described previously [9] regarding cardiomyocyte differentiated from ESCs (Fig. 1A a–c). The differentiated cells positively stained for sarcomeric protein α-actinin, indicating that the ESCs differentiated into cardiomyocytes (Fig. 1A d). Cells were double-stained with antibodies against α-actinin, Troponin T, and mGluR5 at day 5+7. As a result, there was a correlation between α-actinin and mGluR5 levels during differentiation (ie, cardiomyocytes expressing α-actinin also expressed a high level of mGluR5, while negative areas were less positive for mGluR5). Troponin T and mGluR5 protein were also coexpressed in ESC-derived cardiomyocytes (Fig. 1B).
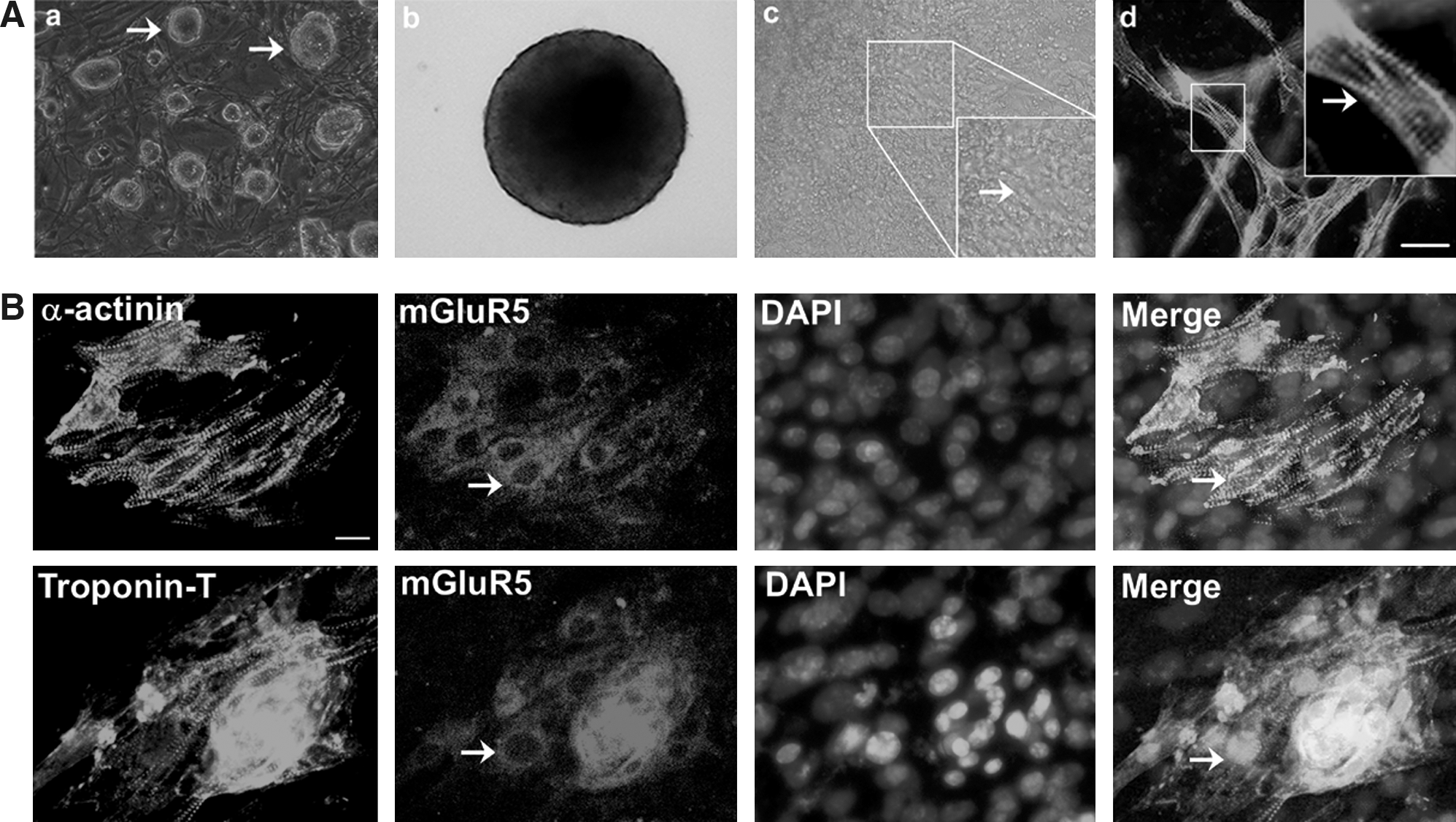
Cardiomyocyte differentiation from mouse embryonic stem cells (ESCs) and immunofluorescent analysis of metabotropic glutamate receptors (mGluR5) expression. (
As shown in Fig. 2A, there was a significant increase in the percentage of EBs containing beating areas from day 5+3 to day 5+11. As determined by semi-quantitative RT-PCR, mRNA expression of mGluR5 was detected in ESCs, which was in accordance with the reported literature [4]. From day 5+3 to 5+11, the level of mGluR5 mRNA expression increased in parallel with contracting cardiomyocyte maturation during differentiation. RA significantly enhanced mGluR5 mRNA expression compared with that of the control. The protein expression level of mGluR5 paralleled the trend of mRNA expression (Fig. 2C). Flow cytometry analysis showed that only 5.61±1.20% of cells double-stained for α-actinin and mGluR5 in the control at day 5+7, while 11.72±2.00% of cells treated with RA were positive staining for α-actinin and mGluR5. Only 3.80±1.06% of cells double-stained for Troponin T and mGluR5 in the control, while the proportion of positive double-staining cells were significantly increased to 8.74±1.53% in the RA treatment group (Fig. 2D). In conclusion, these results suggest that mGluR5 has a developmentally dependent expression pattern during cardiomyocyte differentiation from mouse ESCs.

Expression pattern of mGluR5 during cardiomyocyte differentiation.
To further confirm the expression pattern of mGluR5 in ESC-derived cardiomyocytes, the in vivo presence of mGluR5 in the embryonic heart was identified with RT-PCR and western blot (Fig. 2B) at embryonic days E16, 18, and 20, and in newborn (N) and adult (A) mouse hearts. Both gene and protein expressions of mGluR5 increased to a maximum level in the newborn stage, and levels were maintained in adult hearts, which is concomitant with mGluR5 expression in the ESC-derived cardiomyocytes.
mGluR5/Homer/PIKE/PI3K signaling pathway proteins expressed in cardiomyocytes derived from ESCs
A time-course of western blot analysis revealed that during cardiomyocyte differentiation, expression of PIKE and PI3K p110α was consistently up-regulated in EBs in the control from day 5+3 to 5+11, while Homer1b/c sustained a relatively steady level during differentiation (Fig. 3D). Furthermore, the results of flow cytometry analysis showed the coexpression ratios of Homer1b/c, PIKE, or PI3K p110α together with Troponin T on day 5+7 were 5.03±1.06%, 3.52±0.80%, and 4.53±1.02%, respectively, in the control (Fig. 3E). In the Troponin T-positive cells derived from ESCs, we observed coexpression of Homer1b/c, PIKE, and PI3K p110α with Troponin T using immunofluorescence on day 5+7. The fluorescence-merged area of PIKE with Troponin T was more correlated than that of Homer1b/c or PI3K p110α with Troponin T (Fig. 2E). These results suggested that mGluR5/Homer/PIKE/PI3K signaling pathway proteins were all expressed in cardiomyocytes derived from ESCs. PIKE may be important to link mGluR5 and PI3K through Homer protein to play a pivotal role in cardiomyocyte differentiation from ESCs.
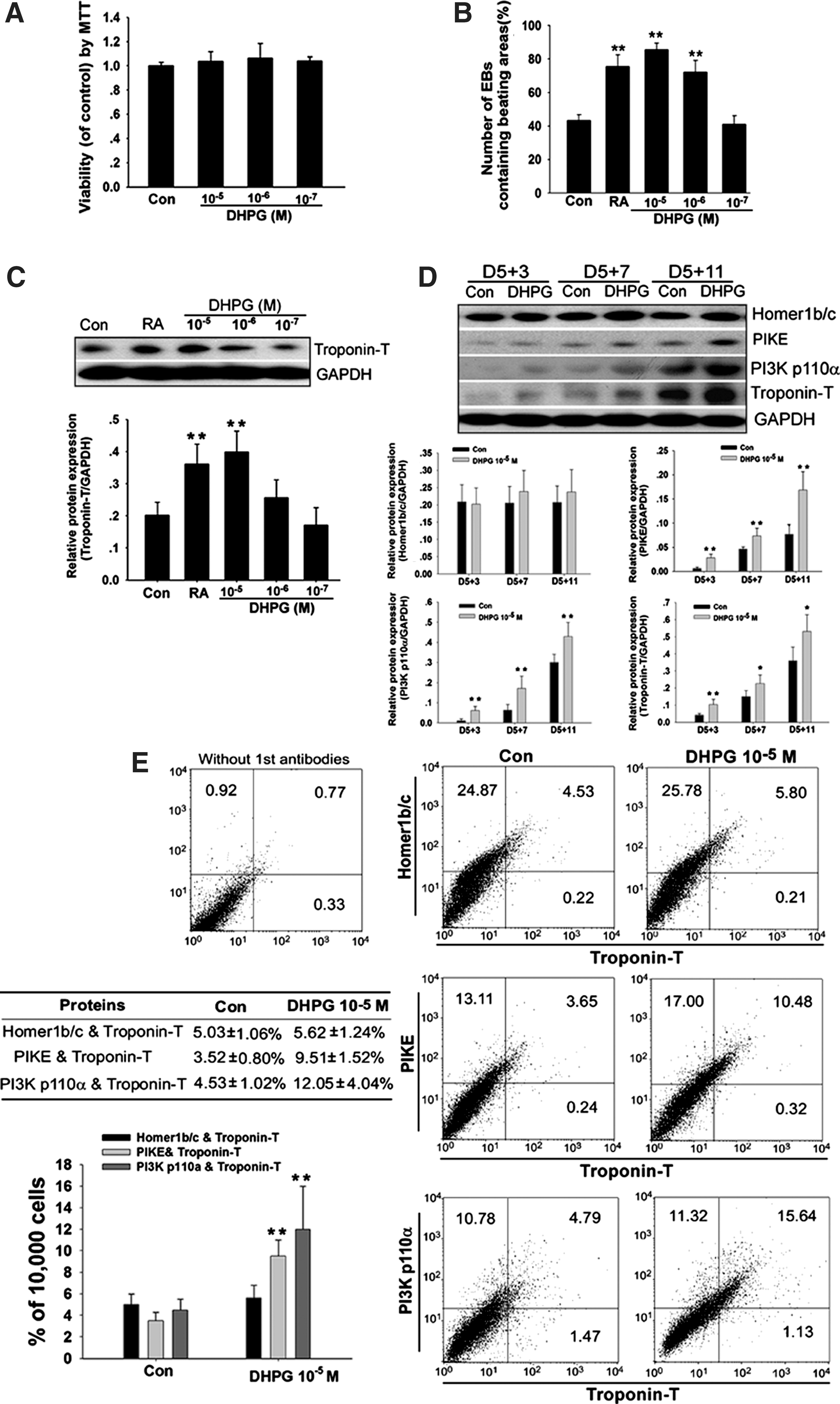
Effect of mGluR5 activator (RS)-3, 5-dihydroxyl- phenylglycine (DHPG) on cardiomyocyte differentiation in vitro.
Specific activation of mGluR5 enhanced cardiomyocyte differentiation from ESCs
The specific activator of mGluR5, DHPG, was employed to evaluate the possible function of mGluR5 in cardiomyocyte differentiation. As shown in Fig. 3A, DHPG at 10−5, 10−6, and 10−7 mol/L concentration had no toxic effects on ESCs. DHPG was applied to EBs from day 5 and the percentage of EBs containing beating areas was evaluated at day 5+7. DHPG significantly increased the percentage of beating EBs in a dose-dependent manner (Fig. 3B). In parallel, expression of Troponin T protein was increased in the DHPG treatment group (Fig. 3C), suggesting that formation of sarcomeric proteins was promoted by DHPG.
Next, we explored whether mGluR5/Homer/PIKE/PI3K signaling was involved in DHPG-induced activation of cardiomyocyte differentiation. As shown in Fig. 3D, DHPG (10−5 mol/L) significantly enhanced PIKE and PI3K p110α protein expression; however, it had no significant effect on Homer1b/c during differentiation, especially at the late stage. The coexpression ratios of Homer1b/c, PIKE, and PI3K p110α with Troponin T in the DHPG treatment group were changed to 5.62±1.24%, 9.51±1.52% (p<0.05), and 12.05±4.04% (p<0.05), respectively, compared with that in the control (Fig. 3E). From these results, we identify that pharmacological activation of mGluR5 enhances cardiomyocyte differentiation through activation of mGluR5/Homer/PIKE/PI3K signaling.
Specific inhibition of mGluR5 blocked cardiomyocyte differentiation from ESCs
As shown in Fig. 4A, MPEP at 10−5, 10−6, and 10−7 mol/L concentration had no toxic effects on ESCs. At first, MPEP was applied to EBs from day 5 and the percentage of EBs containing beating areas was evaluated during differentiation. To our surprise, MPEP could not decrease the percentage of beating EBs containing cardiomyocytes (Fig. 4B). In parallel, expression of Troponin T protein was not decreased in the MPEP treatment group, suggesting that MPEP applied to EBs from day 5 could not inhibit cardiomyocyte differentiation (Fig. 4C). However, when MPEP (10−5 mol/L) was applied to ESCs from hanging drop cultures, it did decrease the percentage of beating EBs containing cardiomyocytes (Fig. 4D). Western blot analysis also detected that MPEP significantly decreased PIKE, PI3K p110α, and Troponin-T protein expression; however, it had no significant effect on Homer1b/c on day 5+7. From these results, we identify that function of mGluR5 is probably indispensable to the early stage of cardiomyocyte differentiation, and inhibition of mGluR5 in ESCs blocked cardiomyocyte differentiation through inhibition of mGluR5/Homer/PIKE/PI3K signaling.
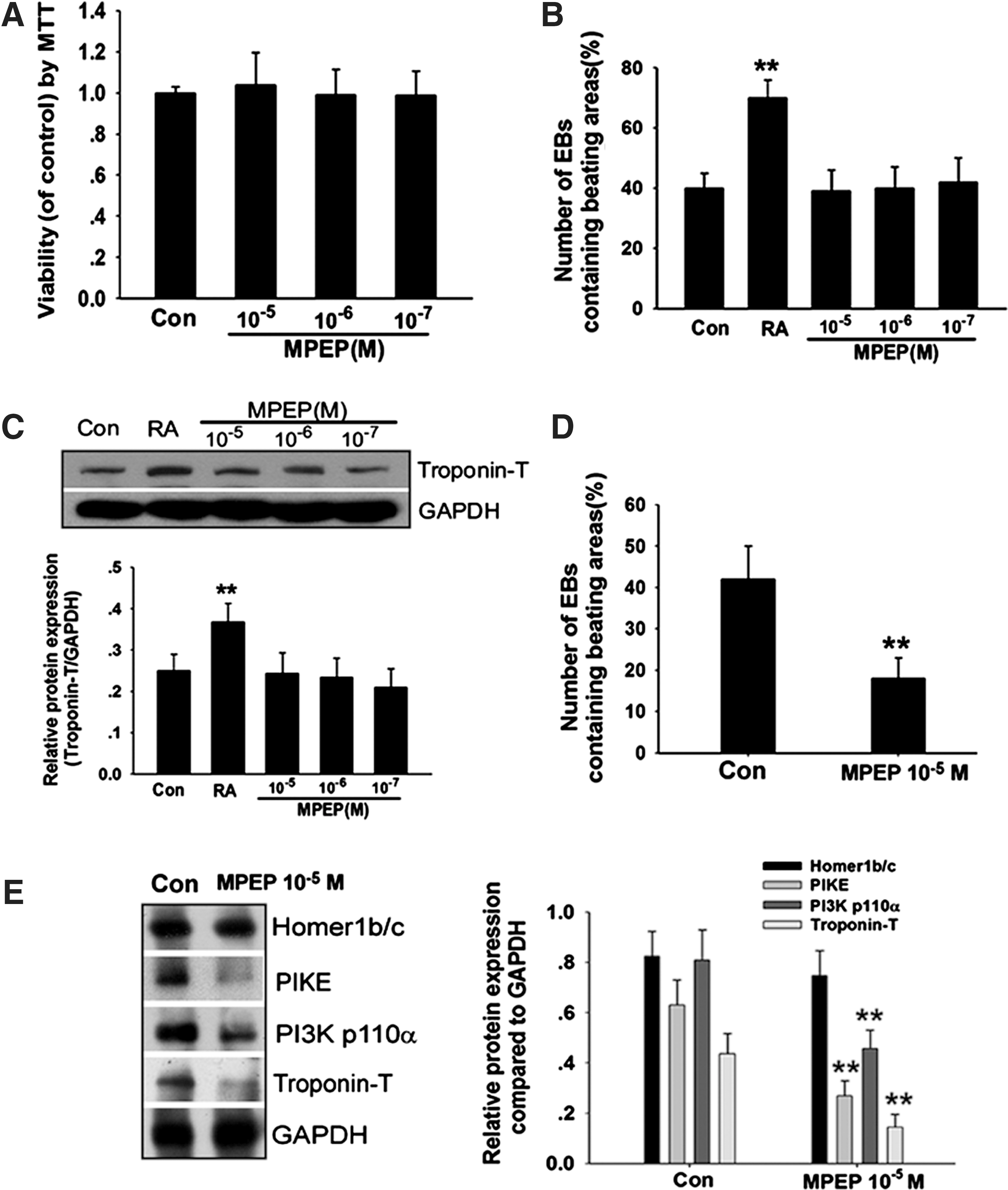
Effect of mGluR5 inhibitor 2-methyl-6-(phenylethynyl pyridine (MPEP) on cardiomyocyte differentiation in vitro.
mGluR5 down-regulation impairment of cardiomyocyte differentiation from ESCs
To further confirm the role of mGluR5 in ESC-derived cardiomyocytes, we enzymatically dissociated day 5+7 EBs and isolated cardiomyocytes with percoll gradient separation. Three siRNA oligos targeting mGluR5 (si-mGluR5-1, si-mGluR5-2, and si-mGluR5-3) were transiently transfected into the isolated cardiomyocytes. By transfecting an irrelevant siRNA-oligo labeled with Alexa-488, we estimated the transfection efficiency to be >90% (Fig. 5A). The efficiency of si-mGluR5-1, si-mGluR5-2, and si-mGluR5-3 was first evaluated by transiently transfecting ESC-derived cardiomyocytes. si-mGluR5-1 and si-mGluR5-3 were determined to substantially decrease the levels of mGluR5, while si-mGluR5-2 was less effective in knocking down mGluR5 (Fig. 5B).

Effect of mGluR5 down-regulation on cardiomyocyte differentiation from ESCs.
Next, we further investigated whether a transient down-regulation of mGluR5 in ESCs could blockade ESC differentiation toward a cardiomyocyte fate. Two effective siRNA oligos targeting mGluR5 (si-mGluR5-1 and si-mGluR5-3) were transiently transfected into ESCs. At day 5+7 of EB differentiation, the percentage of beating EBs was reduced by ∼55% and 85% with si-mGluR5-1 and si-mGluR5-3 (each at 50 nM) compared with si-control, respectively (Fig. 5C). To further confirm the role of mGluR5 at the early stage of cardiomyocyte differentiation (early cardiogenesis), we detected the cardiac transcription factors GATA4 and MEF2C in EBs on day 3, day 5, and day 5+3. As shown in Fig. 5D, si-mGluR5-3 significantly decreased the protein expression of GATA4 and MEF2C on day3, leading to a low expression of the cardiac specific protein α-actinin on day 5+3.
In addition, immunofluorescent analysis detected the coexpression of PIKE and α-actinin in si-mGluR5-3–treated ESC-derived cardiomyocytes on day 5+7. The sarcomeric protein, α-actinin, was disorganized inside si-mGluR5-3–treated ESC-derived EBs, which suggested there was an abnormal cardiac myofibrillogenesis in these cells. Although the PIKE protein was also disorganized in si-mGluR5-3–treated ESC-derived EBs, there was still a good correlation between PIKE and α-actinin, which further confirmed that PIKE play a pivotal role in the mGluR5/Homer/PIKE/PI3K signaling pathway (Fig. 5E).
Concurrently, treatment with mGluR5 siRNAs remarkably decreased the expression of PIKE and PI3K p110α proteins in EBs (Fig. 5F), leading to a dramatic reduction of the beating activity in EBs observed at day 5+7. However, this had no remarkable influence on Homer1b/c. These results indicated that mGluR5 promotes cardiomyocyte differentiation by activation of mGluR5/Homer/PIKE/PI3K signaling pathway.
mGluR5 down-regulation impairment of Ca2+ oscillations and Ca2+ channel protein
To investigate the relationship between the function of mGluR5 and Ca2+ oscillations, we first detected the protein expression of T-type Ca2+ channel and L-type Ca2+ channel in cardiomyocyte differentiation from ESCs. As shown in Fig. 6A, T-type and L-type Ca2+ channels increased between day 5+3 and day 5+11, which was in parallel with the increasing level of mGluR5.
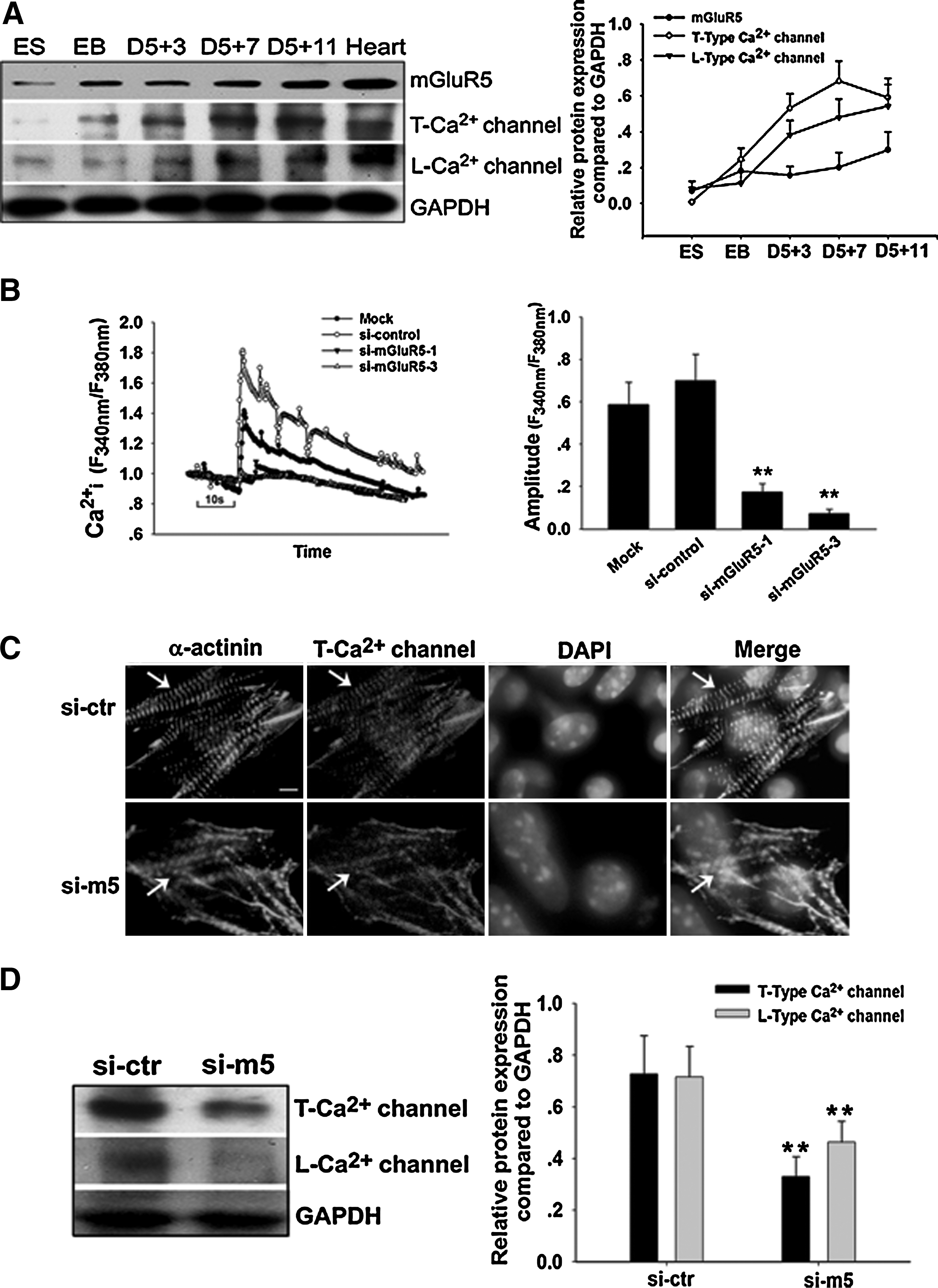
Effect of mGluR5 down-regulation on [Ca2+]i and Ca2+ channel protein.
[Ca2+]i was measured to investigate the function of mGluR5 in cardiomyocytes using Fluo-4-AM as a Ca2+ indicator. Two mGluR5 siRNAs were transfected into the isolated ESC-derived cardiomyocytes. DHPG induced a substantial increase in [Ca2+]i oscillation amplitude in the si-control group. Si-mGluR5-3 substantially decreased the level of DHPG induced [Ca2+]i oscillation amplitude, while si-mGluR5-1 was relatively less effective. This was analogous with the efficiency of the 3 siRNA oligos to repress the expression of mGluR5. The amplitude of [Ca2+]i oscillation was reduced by ∼77% and 90% with si-mGluR5-1 and si-mGluR5-3, respectively, compared with the si-control (Fig. 6B). These data convincingly depict a key role for mGluR5 in the Ca2+ oscillation of ESC-derived cardiomyocytes. On the other hand, the formation of T-type Ca2+ channel protein was disorganized in si-mGluR5-3–treated ESC-derived EBs (Fig. 6C). The expressing level of T-type and L-type Ca2+ channels were also decreased in si-mGluR5-3–treated ESC-derived cardiomyocytes (Fig. 6D). These results suggested that mGluR5 plays a significant role in the Ca2+ channel protein expression and that mGluR5 is essential for cardiomyocyte differentiation from ESCs.
Discussion
It is becoming increasingly evident that mGluR5 is functional in stem/progenitor cells, regulating proliferation, differentiation, and survival of these cells. Cappuccio et al. [4] found that undifferentiated mouse ESCs express mGluR5. This is an intriguing finding, because mGluR5 is conventionally considered as a “synaptic” receptor activated by glutamate released from nerve endings. However, there are few studies to elucidate the role of mGluR5 in non-neuronal cells. Meanwhile, mGluR5 belongs to GPCRs. GPCRs mediate functions of the nervous and immune systems; however, their roles in cardiomyogenesis remain largely unexplored. Scott et al. [10] have explored that the GPCR angiotensin receptor-like 1b regulates early development of myocardial progenitors in zebrafish. Toman et al. [11] have identified that the GPCR endothelial differentiation gene-1 regulates heart development. Knockdown of the GPCR C-X-C chemokine receptor 4 disrupts cardiac differentiation from pluripotent stem cells [12]. Therefore, it is worthwhile to explore the function of mGluR5 and the relationship of GPCRs in the cardiac differentiation.
mGluR5 is coupled with polyphosphoinositide hydrolysis and has an interesting and unique feature among all mGluR subtypes to generate oscillatory increases in intracellular Ca2+ in response to agonist activation [13]. Due to this property, mGluR5 may regulate early events in cell growth and development [4,13]. Previous studies have shown that mGluR4 plays a pivotal role in neuron differentiation and maturation. Until now, very little research has focused on mGluRs in non-neuronal cells. In particular, there have been no reports regarding mGluR5 function in ESC-derived cardiomyocytes. Our results demonstrated that mGluR5 expression increased in a developmental-dependent pattern during cardiomyocyte differentiation from ESCs. The pharmacological activation of mGluR5 could drive differentiation of ESCs into cardiomyocytes. Moreover, we found mGluR4 expression showed robust down-regulation in late differentiation, and mGluR4 was not expressed in the adult mouse heart (unpublished data). These results revealed that mGluR5, but not mGluR4, may be indispensable for cardiomyocyte differentiation and maturation. Our findings revealed that which subtype of mGluRs in ESCs did drive the commitment to differentiate preferentially into cardiomyocytes. This supported the emerging idea that the function of mGluRs extended beyond the regulation of synaptic transmission and involved basic cellular processes, such as proliferation and differentiation of stem/progenitor cells [14].
In the present study, mGluR5/Homer/PIKE/PI3K signaling pathway was confirmed to be involved in cardiomyocyte differentiation from ESCs. Homer proteins have recently been identified as novel high-affinity ligands that modulate the ryanodine receptor Ca2+ release channels in the heart, through an enabled/vasodilator-stimulated phosphoprotein homology 1 domain that binds to proline-rich regions in target proteins. Homer1b/c acted as a scaffold protein, linking mGluR5 to extracellular signal-regulated protein kinase cascades in neurons [15]. Taken together, Homer proteins serve as adaptors that functionally link mGluR5 to intracellular Ca2+ stores. Our results demonstrated that Homer1b/c was highly expressed in ESC-derived EBs. In addition, DHPG could significantly enhance PIKE and PI3K p110α expression at late differentiation, while there was no notable effect on Homer1b/c expression, and similarly in si-mGluR5-3–treated EBs. It is probable that Homer1b/c is also a scaffold protein in cardiomyocytes that remains stable, and it may not be modulated in cardiomyocyte differentiation via regulating mGluR5. Rong et al. [6] have recently identified a new PIKE that links mGluR5 to PI3K through Homer proteins as adaptors that bind mGluR5. mGluR5 agonists enhance mGluR5-Homer-PIKE complex formation, leading to activation of PI3K and inhibition of staurosporine-induced neuronal apoptosis. Our results showed that mGluR5 agonist, DHPG, could activate the mGluR5/Homer/PIKE/PI3K signaling pathway, leading to cardiomyocyte differentiation from ESCs. Small interfering RNA of mGluR5 blocked cardiac differentiation through repressing the mGluR5/Homer/PIKE/PI3K signaling pathway. Among these related signaling proteins, PIKE and PI3K could be greatly elevated with the mGluR5-agonist DHPG or decreased with si-mGluR5-3 treatment during cardiomyocyte differentiation and maturation. We presumed that PIKE played a key role in transmitting signals from mGluR5 to PI3K. Rong et al. [6] showed that the interaction between PIKE and PI3K is guanosine triphosphate (GTP)-dependent, a feature similar to the Ras family of GTPases, suggesting that PIKE serves as a GTPase. Moreover, mGluR5 also belongs to the G-protein coupled receptors. Our results suggested that mGluR5 may be directly regulating the PIKE/PI3K signaling pathway and is involved in cardiomyocyte differentiation.
The robust oscillatory pattern of Ca2+ signaling initiated by activation of mGluR5 has been proposed to be a result of a “dynamic uncoupling” mechanism. This involves rapid cycles of receptor phosphorylation and dephosphorylation with Ser 839 being implicated most recently as the site of reversible covalent modification. Following agonist stimulation, mGluR5 is rapidly phosphorylated by PKC, disabling productive receptor-G protein coupling [16]. This mechanism is probably similar to that reported for Ca2+ oscillations initiated by Ca2+-sensing receptor activation [17]. However, this is clearly different from the Ca2+-induced mechanism that releases Ca2+ proposed to explain the majority of Ca2+ oscillatory behaviors elicited by (submaximal) agonist stimulation of various G protein-coupled receptors [18]. mGluR5 density is the predominant controlling factor of the oscillation frequency in mGluR5a expressed in Chinese hamster ovary cells [19]. Our results demonstrated that the mGluR5 agonist DHPG could induce a substantial increase in [Ca2+]i oscillation amplitude. In addition, siRNA of mGluR5 could substantially decrease the levels of the DHPG-induced [Ca2+]i oscillation amplitude. These results implied that altering mGluR5 expression has notable effects on the amplitude of Ca2+ oscillation stimulated by the mGluR5 agonist DHPG. Cytoplasmic Ca2+ is important already early in embryogenesis in the specification and commitment steps of the cardiac precursor cell population leading to differentiation, before it becomes necessary for excitation–contraction coupling [20]. Our results suggested that only early suppression of mGluR5 in ESCs could inhibit cardiomyocyte differentiation, and decrease in intracellular Ca2+ in response to agonist activation. The function of mGluR5 is probably related to Ca2+ at the early stage of cardiomyocyte differentiation, and robust mGluR5-elicited Ca2+ oscillation is indispensable in cardiomyocyte differentiation from ESCs.
Exposure to the L-type channel blockade with diltiazem resulted in an inhibition of cardiomyogenesis in the embryonic cardiac progenitor cells [20]. T-type Ca2+ channels play an important role in the morphogenesis of myocardium and cardiac conduction system during mouse embryonic heart development [21]. mGluR5-dependent Ca2+ elevations is largely due to the influx of extracellular Ca2+[22]. Our results suggested that the protein expression of T-type and L-type Ca2+ channels were decreased in si-mGluR5-3–treated ESC-derived cardiomyocytes. As we know, L-type Ca2+ channel and T-type Ca2+ channel are pathways for Ca2+ entry, so we may conclude that suppression of mGluR5 inhibits cardiomyocyte differentiation by blocking the formation of T-type and L-type Ca2+ channels.
In conclusion, mGluR5 shows a developmentally dependent expression pattern during cardiomyocyte differentiation from mouse ESCs. The mGluR5 agonist DHPG enhances the appearance of spontaneously beating cardiac cells and the expression of PIKE and PI3K proteins. The mGluR5 antagonist MPEP, treated on ESCs, inhibits the appearance of spontaneously beating cardiac cells and the expression of PIKE and PI3K proteins. In addition, siRNA down-regulation of mGluR5 decreases PIKE and PI3K proteins, resulting in cardiogenesis suppression from ESCs. It also inhibits T-type Ca2+ channel and L-type Ca2+ channel proteins. Moreover, mGluR5 siRNA reduces the DHPG-induced Ca2+ transient peak amplitude in the isolated ESC-derived cardiomyocytes. We conclude that the mGluR5/PIKE/PI3K signaling pathway plays an important role in cardiomyocyte differentiation from mouse ESCs, and that the function of mGluR5 is probably related to the cardiogenesis and Ca2+ signal in cardiomyocytes differentiated from ESCs.
Footnotes
Acknowledgments
This work was supported by the National Natural Sciences Foundation of China (No. 30873068, 30600762), the Scientific Foundation of the Bureau of Traditional Chinese Medicine of Zhejiang Province (No.2009CA052), the Education of Zhejiang Province (No.Y201018522), and the Key Creative Team of Zhejiang Province (No. 2010R50047).
Author Disclosure Statement
The authors indicate no potential conflicts of interest.