
Abstract
Tumor necrosis factor-alpha (TNF-α) is one major inflammatory factor peaking at 24 h after bone fracture in response to injury; its role in bone healing is controversial. The aims of this study were to investigate whether the duration of exposure to TNF-α is crucial for the initiation of bone regeneration and to determine its underlying mechanism(s). We demonstrated that 24 h of TNF-α treatment significantly abrogated osteocalcin gene expression by human primary osteoblasts (HOBs). However, when TNF-α was withdrawn after 24 h, bone sialoprotein and osteocalcin gene expression levels in HOBs at day 7 were significantly up-regulated compared with the HOBs without TNF-α treatment. In contrast, continuous TNF-α treatment down-regulated bone sialoprotein and osteocalcin gene expression. In addition, in an indirect co-culture system, HOBs pretreated with TNF-α for 24 h induced significantly greater osteogenic differentiation of adipose tissue-derived mesenchymal stem cells (ASCs) than the HOBs without TNF-α treatment. TNF-α treatment also promoted endogenous bone morphogenetic protein 2 (BMP-2) production in HOBs, while blocking the BMP-2 signaling pathway with Noggin inhibited osteogenic differentiation of ASCs in the co-culture system. Furthermore, activation of the p38 mitogen-activated protein kinase (MAPK) signaling pathway after TNF-α treatment occurred earlier than BMP-2 protein expression. BMP-2 production by HOBs and osteogenic differentiation of ASCs in the co-culture system with HOBs was significantly decreased when HOBs were pretreated with TNF-α in combination with the p38 MAPK-specific inhibitor (SB203580). Taken together, we provide evidence that exposure duration is a critical element in determining TNF-α's effects on bone regeneration. We also demonstrate that the p38 MAPK signaling pathway regulates the expression of BMP-2 in osteoblasts, which then acts through a paracrine loop, to direct the osteoblast lineage commitment of mesenchymal stem cells.
Introduction
A
Bone remodels itself according to functional demands, adding new bone where it is needed and removing bone where it is not required. Bone remodeling begins with bone resorption by osteoclasts, involving the removal of both matrix and mineral components, followed by bone formation by osteoblasts. On bone injury, osteoblasts migrate to the injury site, contributing to bone formation by synthesizing various bone extracellular matrix molecules that are then mineralized [3 –5]. In addition, an increasing body of evidence suggests that osteoblasts are able to contribute to bone formation by producing a pool of growth factors such as bone morphogenetic protein 2 (BMP-2) and vascular endothelial growth factor [6 –11], which induce the co-migration of mesenchymal stem cells and endothelial progenitor cells into the repair site and direct their differentiation into the osteogenic and endothelial lineages, respectively. Our previous studies further demonstrated that the capacity of producing a pool of growth factors by osteoblasts is largely dependent on the interactions between osteoblasts and their residing microenvironment such as the substrates [7,12].
Inflammation is a common feature after tissue injuries and involves recruitment to the injury sites of inflammatory cells, such as macrophages, which release various inflammatory cytokines. Increasing evidence suggests that inflammation may play a key role in tissue regeneration after injuries [13]; however, this remains controversial in bone regeneration. On the one hand, various pro-inflammatory cytokines are best known for their catabolic effects of inducing bone resorption and inhibiting bone formation. These actions are demonstrated in osteoporosis, rheumatoid arthritis [14 –18], and in the impaired bone fracture healing after prolonged excessive exposure to tumor necrosis factor-alpha (TNF-α) [19]. On the other hand, there is evidence suggesting that inflammation plays a vital role in the initiation of bone fracture healing [20 –24], and this critical role in bone regeneration seems to be orchestrated by the early burst of various inflammatory factors after bone fracture. However, in contrast to the prolonged presence of inflammation in chronic diseases such as osteoporosis and rheumatoid arthritis, various inflammatory cytokines, including TNF-α, interleukin 1 (IL-1), and IL-6, are transiently expressed at the bone fracture site, peaking within 24 h of injury and dropping to basal levels within 2 to 3 days [25]. The distinct patterns of inflammation that mediate positive or negative effects on bone formation led us to hypothesize that the contradictory effects of inflammation on the bone depend on its duration: transient exposure to inflammatory cytokines stimulates bone formation, but prolonged exposure inhibits bone formation.
The aims of this study were to determine how TNF-α, one of the major inflammatory cytokines, contributes to bone formation and to investigate the mechanisms for its actions. We tested whether the short exposure of TNF-α has a role in the initiation of bone formation either by direct stimulation of osteoblast differentiation and/or by indirectly enabling osteoblasts to produce the relevant growth factors constituting an osteogenic microenvironment. We demonstrated that a short-duration exposure (24 h) of TNF-α (which mimics the natural induction course of inflammation on bone injury) not only promoted osteogenic differentiation of human primary osteoblasts (HOBs) but also stimulated HOBs to foster a microenvironment that directed the osteogenic differentiation of adipose tissue-derived mesenchymal stem cells (ASCs) in a co-culture system. We further provided evidence that these actions occurred at least in part via a BMP-2 paracrine loop regulated by the activation of the p38 mitogen-activated protein kinase (MAPK) signaling pathway in HOBs.
Materials and Methods
HOBs and ASCs culturing
Permission to use discarded human tissue was granted by the Human Ethics Committee of the University of Sydney, and informed consent was obtained. Human trabecular bone (female, 10–20 years old) was chopped into 1 mm3 pieces and washed several times in phosphate-buffered saline (PBS), followed by digestion for 90 min at 37°C with trypsin/ethylenediaminetetraacetic acid (EDTA; Sigma-Aldrich) in PBS. Digested cells were cultured in complete media containing α-minimal essential medium (Gibco Laboratories), supplemented with 10% (v/v) heat-inactivated fetal calf serum (Gibco Laboratories), 2 mM
ASCs (Invitrogen) were propagated according to the manufacturer's instructions, and MesenPRO RS™ Basal Medium (Invitrogen) with the supplementations of 2 mM
HOBs-ASCs co-culturing
Cell culture inserts (Millipore), with a 0.4-μm pore-size filter, were placed in 6-well plates for the indirect co-culture of ASCs with HOBs. The ASCs were freshly digested from the culture flask and seeded on the cell culture inserts with a cell density of 8,000 cells/cm2 (∼50% confluence). The HOBs were seeded on the 6-well culture plates with a cell density of 10,000 cells/cm2. The HOBs were either cultured in normal growth medium or pretreated with 1 ng/mL TNF-α for 24 h followed by washing thrice with warm PBS to eliminate the remnant TNF-α. The relevant osteogenic genes and proteins were determined after 4 and 14 days of co-culturing, respectively.
TNF-α treatment and BMP-2/p38 MAPK signaling blocking
Three groups were set up to test the effects of TNF-α treatment on HOBs: no TNF-α treatment, 24 h of TNF-α treatment (1 ng/mL; Sigma, withdrawn after 24 h), and continuous TNF-α treatment (1 ng/mL). In the co-culture system of HOBs and ASCs, HOBs were used either without TNF-α treatment or with TNF-α treatment for 24 h, and then co-cultured with ASCs for the designated time points. Noggin (500 ng/mL; Sigma) was supplemented into the co-culture medium to block the BMP-2 signaling pathway. The p38 MAPK-specific inhibitor (SB203580, 10 μM; Cayman Chemical) was added to the HOBs culture medium to test its effect on BMP-2 production by TNF-α treated HOBs.
Cell proliferation assay
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay was used for assessing HOB cell proliferation after 1 and 7 days of culturing either with 24 h or continuous TNF-α treatment or without TNF-α treatment. One hundred micro liters of the reacted reagent from each well was transferred to a 96-well plate, and the absorbance was recorded using a microplate reader (PathTech) at a wavelength of 490 nm.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from HOBs and ASCs by adding Trizol reagent (Sigma) after the medium had been removed, according to the manufacturer's instructions. First-strand cDNA was synthesized from 0.7 μg total RNA using the Omniscript RT Kit (Qiagen) according to the manufacturer's instructions. Real-time polymerase chain reaction (PCR) was performed in Rotor-Gene 6000 (Corbett Life Science) by using Immomix (Bioline) according to the manufacturer's instructions. The Immomix (a complete ready-to-use heat-activated 2×Reaction Mix) was mixed with water, cDNA template, primers, and CYBR Green, and then preheated to 95°C for 10 min to carry out PCR assays. The relative gene expression levels for runt-related transcription factor 2 (RUNX2), osteopontin, bone sialoprotein, osteocalcin, and BMP-2 were obtained by normalizing them to the house-keeping gene (18S). Primers for the selected genes are listed in Table 1.
Alkaline phosphatase enzyme activity
For alkaline phosphatase (ALP) activity, the cell layers were washed gently, lysed in Tris buffer containing 0.2% NP-40 solution, sonicated, and centrifuged. Two micro liters of the lysate was added to 100 μL of 16.3 m
Western blotting
HOBs were washed with ice-cold PBS and lysed for 30 min in ice-cold RIPA lysis buffer [20 mM Tris-HCl, pH 7.5, 1 mM EDTA, 1 mM ethylene glycol tetraacetic acid (EGTA), 150 mM NaCl, and 1% Triton X-100, protease inhibitor cocktail (Sigma) and phosphatase inhibitor cocktail 2 (Roche)]. Protein concentration was measured using the BCA protein assay kit (Pierce). Equal aliquots of protein (20 mg) were heated at 70°C for 10 min in 4×sample buffer (WesternBreeze; Invitrogen) and 10×reducing buffer, and separated on 8%–12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels (WesternBreeze; Invitrogen). The proteins were transferred to polyvinylidene fluoride membranes, washed with 1×TBS-T (20 mM Tris-HCl pH 7.6, 137 mM NaCl, containing 0.1% Tween 20), and blocked for 1 h at room temperature in 1×TBS-T with 1% bovine serum albumin (BSA). The membranes were washed thrice followed by incubation with primary antibody anti-BMP-2 (1:1,000, Cell Signaling), anti-p38 MAPK1/2 (1:1,000, Cell Signaling), and anti-phospho-ERK1/2 (1:500), Cell Signaling] in TBS-T containing 1% BSA overnight at 4°C. After 3 washes, the membranes were incubated with secondary antibody (WesternBreeze; Invitrogen) for 60 min, followed by another 3 washes before the protein bands were visualized with chemiluminescent reagents (WesternBreeze; Invitrogen) in the Alpha Innotech Digital Imaging System (Alpha Innotech).
Statistical analysis
Data in this study were obtained from 4 independent experiments and represented with mean±standard error. For statistical analysis, first, Levene's test was performed to determine the homogeneity of variance for all the data, and then, the independent t-test was used for analyzing the data between 2 groups, while the Tukey HSD post-hoc test was used for the data among 3 groups. The SPSS 17.0 program was employed for all statistical analysis, and differences were considered significant if P<0.05.
Results
Effects of TNF-α treatment on proliferation and osteogenic gene expression of HOBs
The effects of TNF-α treatment on the proliferation of HOBs were examined. There was no significant difference found among the 3 groups where HOBs were neither treated with TNF-α, nor treated with 24 h of TNF-α, or continuous TNF-α after 1 and 7 days of culturing, and the cell numbers of HOBs in all 3 groups were significantly increased by 5-fold from day 1 to 7 (Fig. 1).
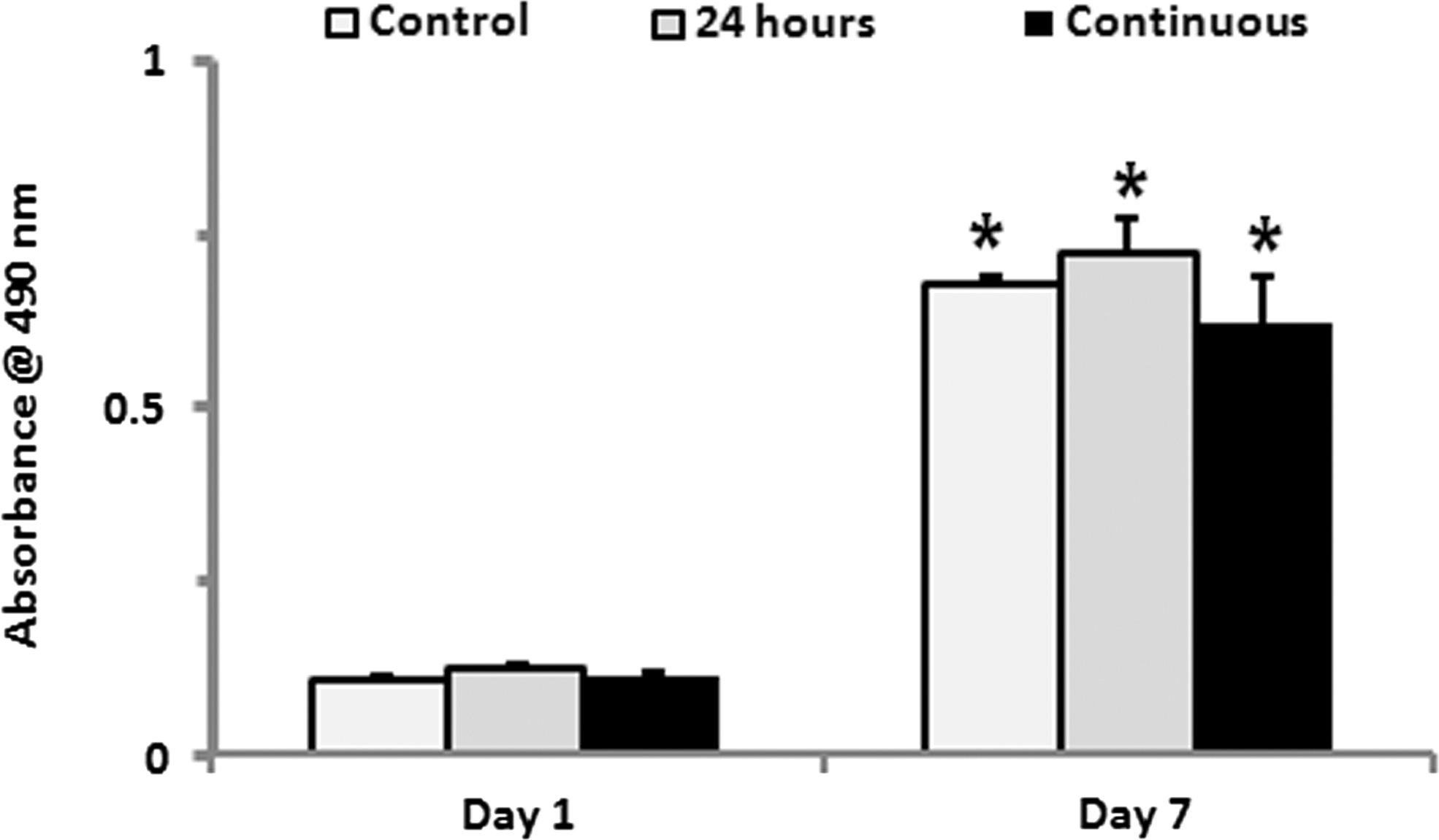
Effects of TNF-α treatment on cell proliferation of HOBs. 24 h or continuous treatment of TNF-α did not significantly affect the proliferation of HOBs, and the HOBs were significantly increased by 5-fold from day 1 to 7 in all 3 groups tested. *P<0.05. TNF-α, tumor necrosis factor-alpha; HOBs, human primary osteoblasts.
Gene expression analysis results showed that, in comparison with the control group (HOBs without TNF-α treatment), osteocalcin gene expression levels were significantly down-regulated after 24 h of TNF-α treatment, while expression levels of Runx2 and bone sialoprotein were not altered (Fig. 2A–C). Interestingly, the continuous treatment of TNF-α (up to 7 days) decreased Runx-2, bone sialoprotein, and osteocalcin gene expression levels, while the withdrawal of TNF-α after 24 h of treatment led to a significant induction of bone sialoprotein and osteocalcin gene expression at 7 days of culture, compared with those with continuous TNF-α treatment and the control group (HOBs without TNF-α treatment) (Fig. 2A–C).
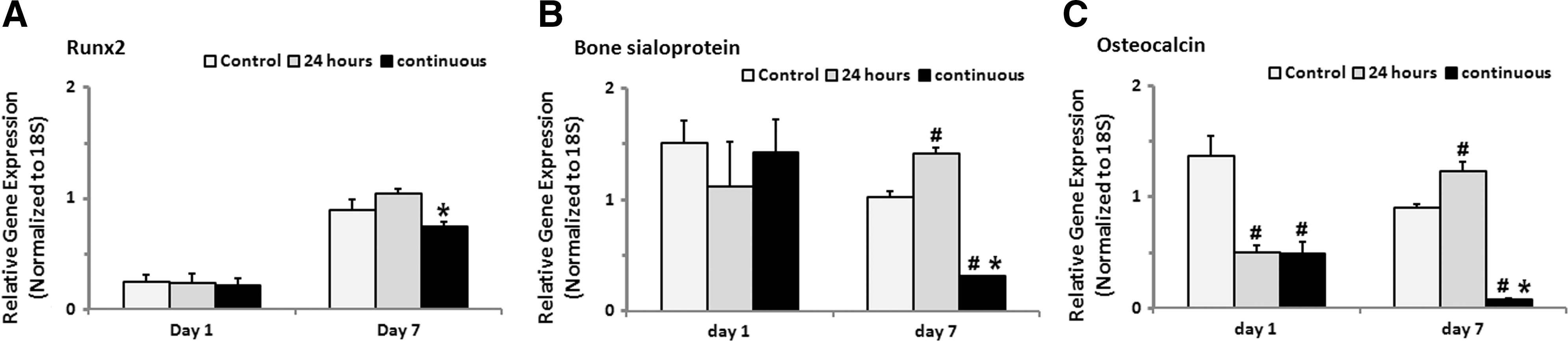
Effects of TNF-α treatment on osteogenic gene expression of HOBs. Osteocalcin gene expression of HOBs was significantly reduced after 24 h of TNF-α treatment, and Runx2, bone sialoprotein, and osteocalcin gene expression was further down-regulated at day 7 with the continuous presence of TNF-α
HOBs with 24 h of TNF-α treatment induced osteogenic differentiation of ASCs in the indirect co-culture system
To determine how TNF-α treatment affects the soluble microenvironment of HOBs, ASCs were either cultured alone (control group) or co-cultured with HOBs for 4 and 14 days. We showed that cultured HOBs without TNF-α pretreatment resulted in a significant induction in Runx-2, osteopontin, and bone sialoprotein gene expression and ALP activity of ASCs at day 4 and/or 14 in the co-culture system, and these increases in expression were further enhanced when the HOBs were pretreated with TNF-α for 24 h (Fig. 3A–D).
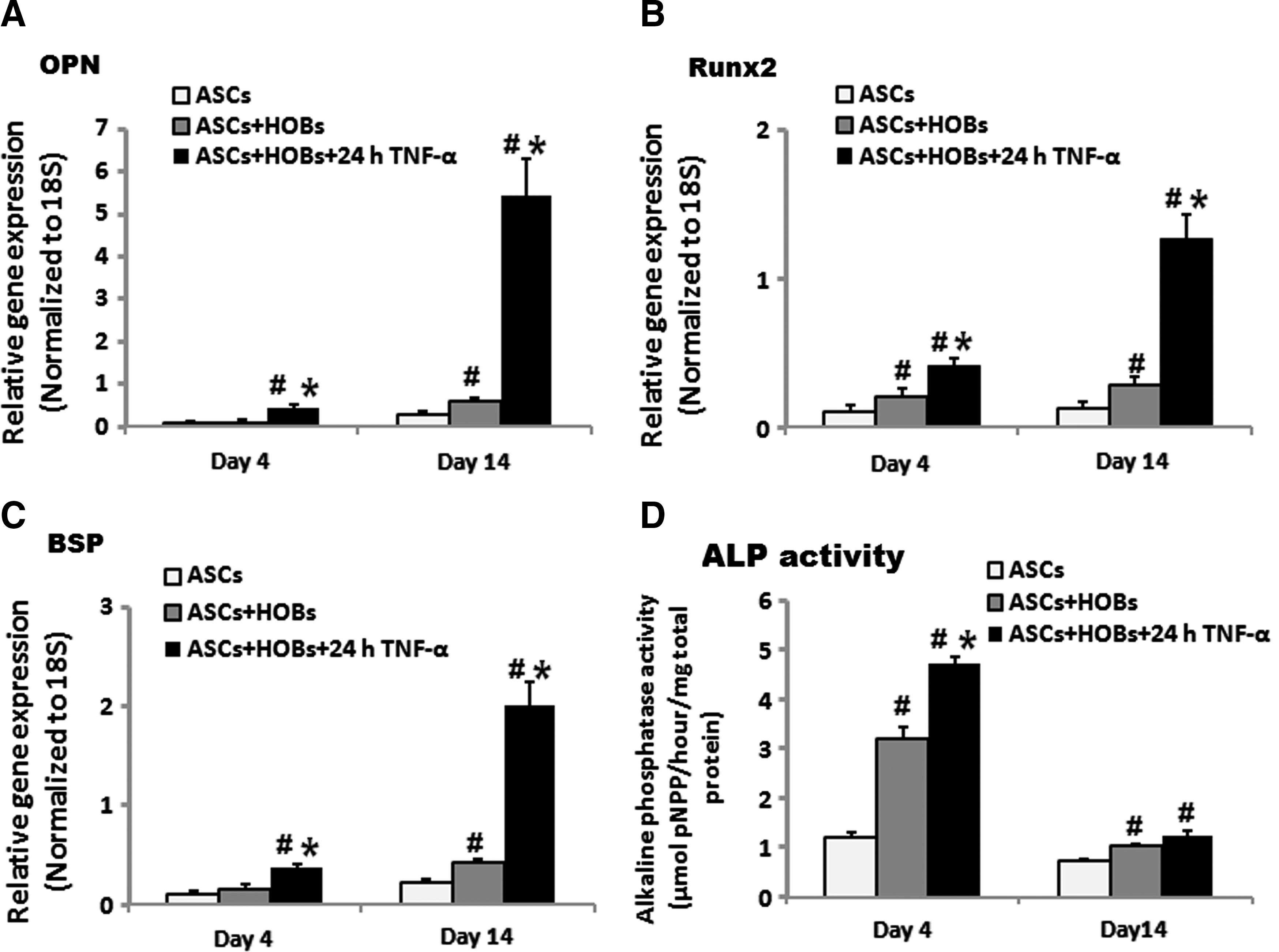
Effects of HOBs with 24 h of TNF-α treatment on osteogenic differentiation of ASCs in the indirect co-culture system. 24 h of TNF-α pretreatment further increased Runx-2
HOBs induced osteogenic differentiation of ASCs via a paracrine BMP-2 loop
In order to investigate the mechanism(s) by which HOBs treated with 24 h of TNF-α induced osteogenic differentiation of ASCs, BMP-2 gene and protein expression levels of HOBs were determined after HOBs had been treated with TNF-α for 24 h and cultured for another 6 days. We found that short treatment (24 h) of HOBs with TNF-α led to a significant increase in mRNA and protein levels of BMP-2 at day 1, compared with the non-TNF-α treated group (Fig. 4A, B).
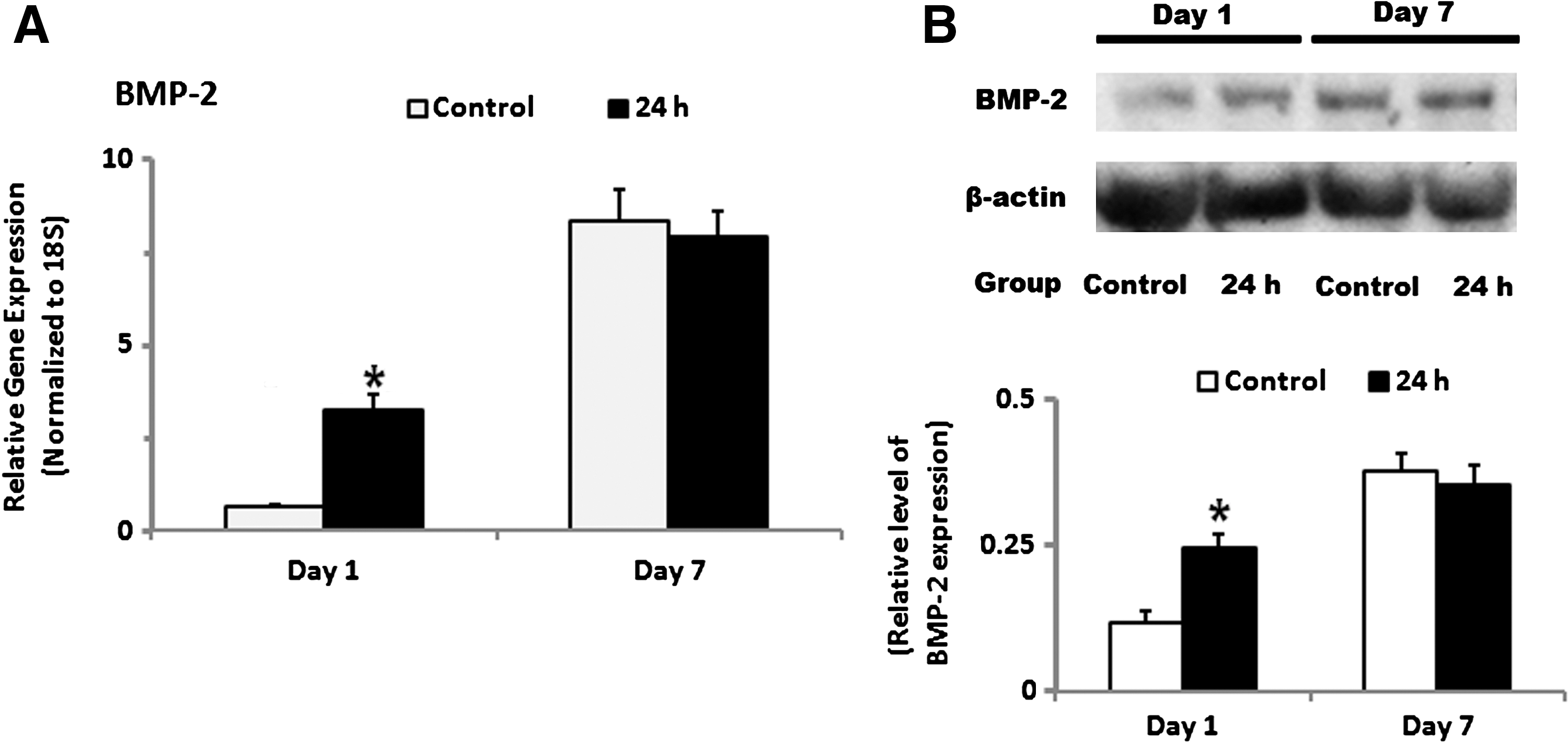
Effects of TNF-α treatment on BMP-2 production in HOBs. HOBs on 24 h of TNF-α treatment showed significantly higher BMP-2 gene as well as protein expression than those without TNF-α treatment at day 1 and/or day 7
To confirm the role of BMP-2 produced by HOBs in osteogenic induction of ASCs in the co-culture system, functional blocking of BMP-2 was performed by adding the BMP-2 inhibitor noggin into the co-culture medium. It was found that noggin treatment not only significantly reduced the levels of osteopontin, Runx2, and bone sialoprotein gene expression of ASCs after 14 days of co-culturing with HOBs (Fig. 5A), but also significantly reduced the ALP activity of ASCs in the co-culture system after 4 and 14 days (Fig. 5B).

Effects of BMP-2 signaling blocking on osteogenic differentiation of ASCs in the indirect co-culture system with HOBs pretreated with 24 h of TNF-α. Noggin treatment significantly reduced the gene expression of osteopontin, Runx2, and bone sialoprotein of ASCs in the co-culture system after 14 days
p38 MAPK signaling pathway is involved in TNF-α-elicited endogenous BMP-2 production in HOBs
To gain an insight into the mechanism(s) by which TNF-α treatment is able to induce endogenous BMP-2 production in HOBs, the p38 MAPK signaling pathway and BMP-2 protein levels in HOBs were determined after 1, 3, 5, and 24 h of TNF-α treatment. The induction of the relative phosphorylated p38 MAPK protein in HOBs increased 5-fold after 1 h of TNF-α treatment, and then decreased but still maintained about 4, 3, and 2-fold increases at 3, 5, and 24 h respectively. BMP-2 protein level did not significantly change at 1 h, but increased 2-fold after 3, 5, and 24 h of treatment (Fig. 6A). The p38 MAPK inhibitor SB203580 was added into the culture medium to investigate whether the p38 MAPK signaling pathway is involved in the endogenous BMP-2 production induced by TNF-α treatment. The results showed that the treatment of SB203580 did not significantly affect BMP-2 expression levels at 1, 3, and 5 h, but significantly reduced expression levels at 24 h (Fig. 6B). In addition, when the HOBs were pretreated with TNF-α in combination with SB203580 for 24 h before co-culture with ASCs, the presence of SB203580 in the pretreatment regime significantly reduced the gene expression levels in the ASCs of Runx2, osteopontin, and bone sialoprotein (Fig. 6C) and the ALP activity after 14 days of co-culturing (Fig. 6D).
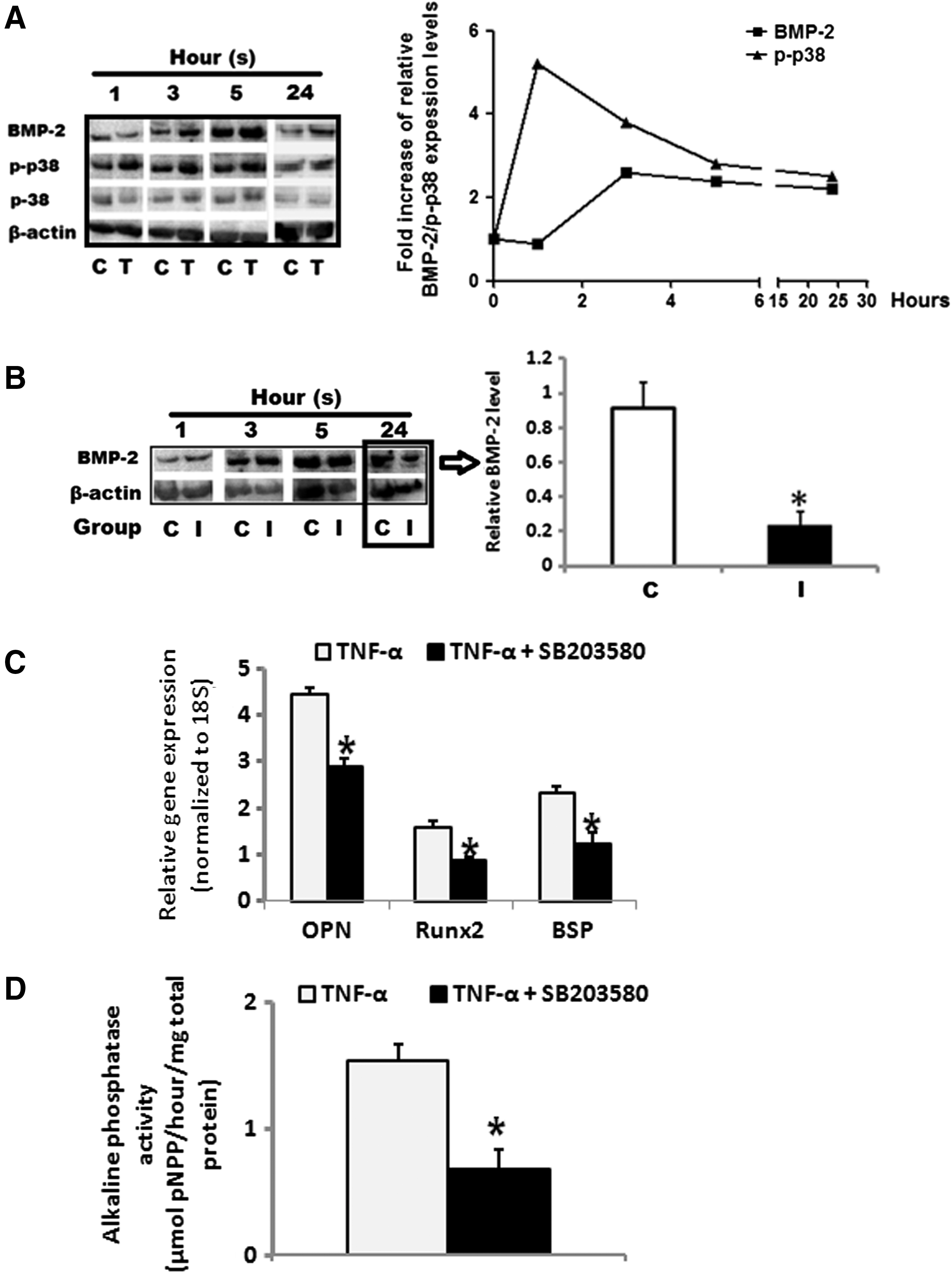
The roles of the p38 MAPK signaling pathway in TNF-α-induced BMP-2 production in HOBs. Phosphorylated p38 MAPK protein was increased up to 5 fold after 1 h of TNF-α treatment, and remained high till 24 h, while BMP-2 protein was not increased after 1 h but increased about 2-fold after 3, 5, and 24 h of treatment
Discussion
The present study demonstrated that 24 h of TNF-α treatment of HOBs significantly abrogated their osteocalcin gene expression, but the withdrawal of TNF-α after 24 h led to the recovery of Runx2, bone sialoprotein, and osteocalcin gene expression to levels that were even significantly higher than those of HOBs without TNF-α treatment. In addition, HOBs pretreated for 24 h with TNF-α, followed by indirect co-culture with ASCs, induced the osteogenic differentiation of ASCs via a BMP-2 dependent pathway. The p38 MAPK signaling pathway was found to be involved in regulating BMP-2 production in HOBs. The increased expression of BMP-2 in HOBs treated with TNF-α and the inhibition by noggin of HOB's actions on indirectly co-cultured ASCs are indicative of the presence of a BMP-2-mediated paracrine loop between HOBs and ASCs.
Osteoblastic development is divided into 2 stages: cell proliferation and differentiation, with progress from extracellular matrix synthesis to maturation and mineralization accompanied by dynamic changes in osteogenic gene expression [26]. In this study, TNF-α treatment for up to 7 days did not significantly change the HOBs' proliferation, suggesting that a local TNF-α boost at the bone injury site would not impair osteoblast proliferation. Contradictory effects of TNF-α on osteogenic differentiation have been reported in numerous studies [15 –17,23,27 –29]; on the one hand, TNF-α is a negative regulator of osteoblastic differentiation both in vitro and in vivo; on the other hand, the absence of TNF-α signaling is associated with impaired fracture healing. However, the studies just mentioned neglected the fact that the induction of inflammatory factors after bone injury is dynamic and transient and TNF-α is continuously applied in these studies. In this study, we mimicked the short boost in TNF-α levels occurring on injury and in differentially treated HOBs with TNF-α (24 h or continuous) and evaluated the corresponding changes in the osteogenic gene expression of HOBs. In line with the reported studies just mentioned, the current study showed both the effects of TNF-α signaling on osteogenic gene expression of HOBs, but we revealed that the duration of TNF-α signaling might be the main factor deciding either anabolic or metabolic effects on bone regeneration. In comparison with the control (HOBs without TNF-α treatment), continuous treatment with TNF-α resulted in the reduction of osteogenic gene expression in HOBs, but short treatment (24 h) with TNF-α led to significant increases in osteocalcin and bone sialoprotein gene expression levels after 7 days of culture. These findings provide evidence explaining the contradictory effects of inflammation on bone formation and suggest the important beneficial role of inflammation peaks within 24 h of bone injury.
We further investigated the effects of TNF-α treatment on the soluble microenvironment generated by osteoblasts, and demonstrated that the HOBs under 24 h of TNF-α treatment induced osteogenic differentiation of ASCs in an indirect co-culture system. These results revealed another critical role of the TNF-α boost after bone injury in promoting bone formation. The TNF-α boost triggers osteoblasts to foster an osteogenic microenvironment, subsequently directing the differentiation fate of the MSCs migrating from the surrounding environment into an osteogenic lineage. In agreement with our findings, a recent study also found that short-term (48 h) TNF-α treatment was able to contribute to bone formation by directly inducing osteogenic differentiation of MSCs, while long-term exposure (4 weeks) inhibited their osteogenic differentiation [30]. However, contrary to our current study, Huang et al. [30] also found that the effects of TNF-α on osteogenic differentiation were dose dependent: 0.1 ng/mL or lower leads to positive effects, while higher than 1 ng/mL TNF-α showed an inhibitory effect. In our study, we showed that all 3 doses (0.1, 1, and 10 ng/mL) of TNF-α had a similar pattern in inducing osteogenic gene expression of HOBs (data not shown). We believe that this discrepancy may result from different regimes of TNF-α stimulation and examination time and the cell types used.
Our proposed explanation for the dual effects of TNF-α on osteogenic gene expression of osteoblasts/MSCs is that TNF-α, on one hand, has direct negative effects on osteogenic differentiation; on the other hand, it owes the positive effects to activating some rescuing pathways that can compensate its negative effects. When TNF-α exposure continues for a short time, the rescuing loop might overcome its negative effects; whereas with prolonged exposure, the negative effects dominate. BMP-2 is a key molecule in the early stages of bone healing [31,32], and it has been widely used for bone tissue regeneration [33 –35]. Interestingly, TNF-α was found to be able to induce BMP-2 production in different cell types, including endothelial cells and chondrocytes [36 –38]. From this study, we propose that short TNF-α treatment promotes BMP-2 production in osteoblasts, which subsequently encourages bone formation in a paracrine/autocrine manner. Our results demonstrated that 24 h of TNF-α treatment stimulated BMP-2 production in HOBs, and the supplementation of BMP-2 inhibitor (Noggin) in the co-culture medium inhibited osteogenic gene expression and ALP enzyme activity of ASCs induced by HOBs after 24 h of TNF-α treatment, suggesting that stimulating a paracrine BMP-2 loop in HOBs is one of the important mechanisms of the positive effects on bone formation mediated by a short period (24 h) of TNF-α treatment. However, this direct negative effect of TNF-α might overwhelm its positive effects of the BMP-2 loop if TNF-α is continuously present as in the case of rheumatoid arthritis. In addition, continuous exposure of TNF-α has been shown to be detrimental to BMP-2-induced osteogenic differentiation via the inhibiting Smad signaling and Ras/Rho-MAPK pathway [39].
The noggin used in this study would also be able to modulate the activities of BMP-4,-7,-13, and -14, all known to play an important role in bone development and regeneration. In future studies, it would be of great interest to look into the role(s) of each BMP member in TNF-α-mediated catabolic and anabolic effects on bone regeneration. Another interesting finding of this study is that a short boost in BMP-2 levels in the first 24 h appears to be sufficient and critical for inducing osteogenic differentiation of ASCs, as no difference in BMP-2 gene and protein expression was found between the control and 24 h of the TNF-α treatment group at day 7, and inhibition of BMP-2 production in the HOBs on treatment with the p38 signaling inhibitor abrogated the osteogenic induction of ASCs in the co-culture system. This short boost (24 h) of BMP-2 might model the up-regulated BMP production found during the initial phase in osteotomy, which is essential for accelerating the osteogenic differentiation of recruited MSCs, providing an initial stimulus for bone growth [40]. The finding of this study would be of great importance in optimizing the current BMP-2 therapy regime for promoting bone formation.
We also investigated the mechanism for TNF-α induction of BMP-2 production in osteoblasts. Various mechanisms and signaling pathways have been shown to be involved in BMP-2 gene regulation during limb morphogenesis and osteogenic differentiation [41,42], and a number of transcription factors were identified that bind to the promoter region of BMP-2 gene and regulate its expression [43,44]. As one of the down-stream signaling pathways of both BMP-2 and TNF-α, the p38 MAPK signaling pathway plays a positive role in regulating bone formation [45 –49]. However, several studies demonstrated that the p38 MAPK signaling pathway is also able to induce endogenous BMP-2 production [50 –52]. We propose that in this study, p38 signaling served as the mediator between TNF-α treatment and endogenous BMP-2 production in HOBs, and is responsible for the production of endogenous BMP-2 in HOBs. This hypothesis was further supported by our data where TNF-α treatment activated the p38 MAPK signaling pathway earlier than the BMP-2 protein induction, while the MAPK p38 inhibitor treatment abrogated BMP-2 protein expression in HOBs and subsequently reduced the osteogenic differentiation of ASCs co-cultured with HOBs. It is important to note that the beneficial effects of the TNF-α-p38 MAPK-BMP-2 loop on osteogenic differentiation may only be evident on short-term treatment (24 h). This conclusion is supported by the in vivo results reported by Zhou and his colleagues [53], who in agreement with our study demonstrated activation of the p38-MAPK signaling pathway by TNF-α; however, in contrast to our findings, they demonstrated that the continuous inhibition of the TNF-α-mediated P38-MAPK signaling pathway up-regulated osteogenic gene expression in vivo.
In summary, this study demonstrates that a short exposure to TNF-α, but not continuous exposure, not only promotes osteogenic differentiation of HOBs, but also stimulates HOBs to secrete soluble factors that foster a microenvironment favoring osteogenic differentiation of ASCs. This action is at least, in part, dependent on a BMP-2 paracrine loop that is regulated via the p38 MAPK signaling pathway. The outcome of the study highlights the importance of the timing of TNF-α appearance during bone healing and explains the possible reason for the contradictory effects of inflammation on bone regeneration, and may substantially contribute to developing strategies for bone regeneration by controlling the timing of inflammatory responses after bone injury and/or replacing bone defects with biomaterials.
Footnotes
Acknowledgments
The authors acknowledge the Australia National Health and Medical Research Council (NHMRC) for funding this research. The authors acknowledge Rebecca Cooper Foundation. The authors are also grateful to Mrs. Barbara James for her assistance in isolating HOBs.
Author Disclosure Statement
No competing financial interests exist.