
Abstract
Skin flaps are routinely used in surgery for the functional and cosmetic repair of wounds or disfiguring scars. The recent concept of therapeutic angiogenesis has emerged as an attractive approach to overcome the problem of blood supply deficiency, often resulting in the flap grafting failure. In the present study, we embedded a gelatin membrane with amniotic fluid stem cells (AFSC) derived conditioned media (ACM) to topically deliver angiogenic growth factors and cytokines into a rat model of ischemic full-thickness skin flap elevated in the epigastric region. AFSC secretome triggered the endogenous repair by the recruitment of endothelial progenitor cells. We studied the vascular perfusion rate, the vessel distribution, and the survival of ACM-treated flaps. In detail, the ischemic sectors of ACM-treated flaps showed at day 7 a perfusion level 50% higher than the preoperation baseline. The ensuing necrosis development was delayed and the histology analysis showed a normal arrangement of epidermal and dermal structures and a high density of vessels in subcutaneous tissues. Further, we found that ACM recruited CD31+/VEGFR2+ and CD31+/CD34+ cells into the ischemic subcutaneous tissues and that the isolated progenitors were capable to form clusters of von Willebrand factor-positive cells in culture. We propose ACM as a cell-free cocktail of chemokines and growth factors to be adopted for clinical applications.
Introduction
S
A lack of oxygen and nutrients especially in the distal part of the flap often results in a partial necrosis with devastating results for the patient. The complexity of events associated with ischemia, such as production of reactive oxygen species, neutrophil influx, depletion of NO, and apoptosis have directed the counteracting therapy toward pharmacological approaches, including use of antioxidants [3], vasodilators [4], and anti-inflammatory drugs [5]. The recent concept of therapeutic angiogenesis has emerged as another attractive approach to enhance blood supply and perfusion in compromised flap tissues. Therapeutic angiogenesis is defined as the administration of proangiogenic factors to convert an ischemic tissue into a viable and functional tissue through the maintenance of existing vasculature or by stimulating the formation of new vessels [6,7]. Local and systemic application of angiogenic proteins, such as PDGF [8], basic FGF [9], and VEGF [10] have been demonstrated to improve the survival of compromised flaps, while fibrin sealants have been investigated for their hemostatic and adhesive properties and for the ability to locally deliver and sustainably release growth factors [11,12], thus providing an important role as a biomatrix in pursuing the angiogenic effect.
In line with the current regenerative medicine trend, we recently demonstrated that amniotic fluid stem cells (AFSC) can mediate the tissue repair by recruiting host reparative cells [13] and that their secreted factors, contained in conditioned media, can be responsible for a cell paracrine effect on vessel growth [14]. We strongly believe that the increased blood flux achieved by the administration of the AFSC soluble factors is a keystone of the whole regenerative process, not only because of the increased nutrients perfusion, but also especially for the establishment of an “activated” regeneration niche. In the present study, we embedded AFSC conditioned media (ACM) into a 15% gelatin-membrane to topically deliver angiogenic growth factors and cytokines in a rat model of ischemic full-thick skin flap elevated in the epigastric region. We studied the vascular perfusion rate, the vessel distribution, and the survival of ACM/membrane- and only membrane-treated flaps and demonstrated an ACM-mediated recruitment of endothelial-like progenitors occurring only in the subcutaneous tissues of ACM/membrane-treated flaps.
We want to underline the benefits of a cell-free therapy with no need to differentiate uncommitted stem cells into a specialized tissue both in terms of avoiding allo-rejection and potential tumorigenic risks for the patient, and in terms of cost and operating time savings.
Materials and Methods
Cell culture, conditioned media, and membrane fabrication
Human AFSC were supplied by the Cytogenetic Laboratory of Galliera Hospital as normal karyotype amniocentesis samples from 15 to 17 weeks pregnant women, after obtaining written informed consent. AFSC were isolated as previously described [14,15]. Briefly, back-up cultures from a total of 20 donors were expanded in minimal essential medium alpha (αMEM) medium (Gibco, Milan, Italy) containing 15% embryonic stem cell (ES)-fetal bovine serum (FBS), 1% glutamine, and 1% penicillin/streptomycin (Gibco), supplemented with 18% Chang B and 2% Chang C (Irvine Scientific), at 37°C within a 5% CO2 atmosphere. Once confluent, cells were pooled, labeled with CD117 MicroBeads (Milteyi Biotec), and immunoselected based on c-kit expression on a Mini-MACS apparatus (Miltenyi Biotec) following manufacturer instructions. Selected cells were plated at clonal density (10/cm2) and about 7–10 days after seeding, clones were isolated and replated and passaged few times in dishes with a larger diameter. Two AFSC clonal lines, from the same primary culture and showing a similar phenotype, were arbitrarily used at a culture passage between passages 6 and 10 in our experiments. Cultured cells were tested for the expression of lineage specific surface molecules in a Cyan ADP Cytofluorimeter (Beckman Coulter) and the absence of hematopoietic markers together with the presence of mesenchymal markers were confirmed (Supplementary Fig. S1; Supplementary Data are available online at
To obtain conditioned media, 80% confluent AFSC were plenty washed in PBS and incubated in serum-free αMEM for 16 h [14]. ACM were collected and stored in 5 mL aliquots equivalent to the medium conditioned by 4×106 AFSC. This concentration (1 mL medium conditioned by 800,000 AFSC during a 16 h culture) was used for all in vivo studies.
Lyophilized gelatin film membranes (5×5 cm and 1.5 mm thick) were prepared by casting from a solution of 15% by wt. type A gelatin (from porcine skin, Sigma Aldrich) in water as follows. Briefly, a gelatin/water solution was prepared under gentle stirring at the temperature of 50°C for 1 h, then cast into a polystyrene Petri dish kept at 23°C for 24 h. Afterward, the cast membrane was freeze-dried at −80°C for 72 h. Each membrane was regenerated before further use by its soaking with 5 mL of ACM or saline as control (CTRL). Such a membrane is easy to handle, resorbable within 24–48 h in vivo, it can entrap and slowly release proteins and finally it is a useful delivering substrate suitable for the fasciocutaneous flap experiment protocol.
Fasciocutaneous flap model and treatment
All experiments performed on animals were approved by the local legislative committee in Vienna to the Ludwig Boltzmann Institute and all experimental procedures were consistent with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (Publication No. 85-23, revised 1996). Ten (5 per group) male Sprague Dawley rats weighing 300–350 g (Harlan–Winkelmann) were used to recreate the previously established epigastric flap model [16]. The rats were anesthetized with ketamine [100 mg/kg body weight (BW)] and xylazine (10 mg/kg BW). Briefly, abdominal skin of rats was depilated and a squared fasciocutaneous flap (∼7×7 cm) was elevated in a cranial to caudal direction so that the flap was well detached from the abdominal muscular layer. Conditioned medium containing (ACM) or noncontaining (CTRL) membranes were carefully positioned on the muscular layer (Fig. 1C). To render the flap half ischemic, the left or the right (according to a randomization protocol) inferior epigastric neurovascular bundle was ligated before the split of the epigastric pedicle into a medial and a lateral branch. To avoid neighboring anastomoses, the entire flap was isolated from the surrounding tissues except for the contralateral inferior epigastric neurovascular pedicle that remained pervious and perfused the rest of the flap. The flap was then sutured back to its anatomical location. Animals were followed-up for 1 week, afterward they were sacrificed to take samples for histological and biological studies.

Experimental procedure. Most important passages of the experimental procedure.
Flap perfusion and survival
These 2 parameters were evaluated by 2 independent observers on 3 different vertical zones (vital, border, ischemic; 2.5×7 cm/each) of the flap (Fig. 1D) at different times: before operation, immediately after operation, and 1, 2, 3, and 7 days after surgery. To evaluate flap perfusion, we used the laser Doppler imaging (LDI; Moor Instruments Ltd.) system with a low-intensity (2 mW) laser light beam (wavelength 632.8 nm). The skin surface of supine positioned, anesthetized rats was scanned at a standardized working distance of 20 cm. Blood flow through the flap was documented as 2D color-coded image (setting of scan modus 10 ms/pixel with resolution at 256×256 pixels). Results are expressed as fold increase/decrease of perfusion units (PU) with respect to the preoperative baseline (=1), through the time and for each sector of the flap. The flap survival, as percentage (%) of necrosis, was calculated dividing the flap necrotic area to the entire flap area, multiplied by 100. Flap areas were photoimaged by Moor™ at standardized conditions of light, distance, and magnification, and analyzed with ImageJ 1.44 software (National Institutes of Health). To be considered necrotic, an area should have macroscopically visible brown- to black-colored tissue accompanied by loss of elasticity. As a quality control, areas identified as necrotic were checked for having a Laser Doppler pixel distribution curves ranging between 50 and 70 over a scale 0–255.
Histological analysis
Full-thickness skin samples, proximal to the necrotic area of flaps, were excised from each animal after euthanasia on day 7 and over night fixed in buffered neutral formalin 10% (Sigma), dehydrated in ethanol 70% (2 h), ethanol 80% (2 h), ethanol 96% (2 h), absolute ethanol (2 h), cleared in xylol and embedded in paraffin into an upright position. Fasciocutaneous cross sections were stained with Mallory's trichrome (Bioptica) to evaluate the inflammatory infiltrates (masses of red nuclei), the vascularization of subcutaneous plexus (red blood cells in gold yellow), the thickness of superficial necrotic layers (orange–red), and the organization of cutaneous structures (glands in pale pink and collagen fibers in deep blue). Density of subdermal plexus vessels with a diameter ≥40 μm was determined on 5 random fields (400× magnification), on six 7 mm thick-cross sections for each sample. Thickness of cutaneous dead layers was measured along 3 mm of each section using the Axiovision Rel 4.4 software and distribution curves were generated for each group of treatment.
Recruitment studies and progenitors isolation
Recruitment studies and progenitor isolation were performed on 10 rats. To study the recruitment of progenitor cells into the injured/repaired site, we excised the piece of flap (2×1 cm) showed in Fig. 1E. Briefly, we considered a square delimited by the medial line of the whole flap area and the outlining border of the necrotic area. This squared piece corresponds to the healing region of the flap. In fact, since after the ligation of 1 epigastric bundle just half flap is perfused, the medial line can be considered the “growth plate” of new vessels that from a pre-existing vasculature move toward an ischemic region. That full-thickness skin piece was then sliced according to depth [17]. The upper part (from the surface to a 0.15–0.20 mm depth) contained the epidermis and the papillary dermis and was discarded. The rest of the skin, corresponding to the reticular dermis, hypodermis, and fascia, being the most vascularized part of the skin, was used in our studies to isolate and identify endothelial progenitors. Samples were minced, washed in HBSS/30 mM HEPES (Euroclone) plus penicillin and streptomycin, digested in 0.25% trypsin (Gibco) and 200 U/mL type 2 Collagenase (Biochrom) for 60 min in 37°C water bath, and finally passed through a 70 μm nylon mesh to obtain a single cell suspension. Cells were centrifuged at 300 g and the lipid phase was removed from the top and discarded. Freshly isolated cells were either analyzed with a Cyan ADP cytofluorimeter (Beckman-Coulter) for the presence of different cell surface antigen markers or plated at a density of 8,000 cells/cm2 on fibronectin-coated Petri dishes (BD Biosciences). This preplating step was introduced to separate cells of the stromal-vascular fraction (SVF) from the less adherent endothelial progenitors. Forty-eight hours after plating, nonadherent suspended cells were collected and replated (25 cells/cm2) on fibronectin-coated plates to assess the presence of clonogenic progenitors [18]. We chose a fibronectin coating to provide the endothelial-lineage cells with a matrix suitable for their attachment and expansion. Medium (DMEM/F12, 10% FBS, without any supplemental growth factors) was changed every 3 days. Seven days after the replating, colonies were fixed in 10% formalin and stained for the specific endothelial marker Von Willebrand Factors [Rabbit polyclonal anti-von Willebrand factor (vWF) by DAKO and TRITC-conjugated goat anti-rabbit IgG by Jackson Immunoresearch Labs].
Antibodies used for the cytofluorimetric analysis were: APC-conjugated anti-CD31 (clone 390), PE-conjugated anti-CD34 (clone RAM34) by E Bioscience, and PE-conjugated anti-Flk1/VEGFR2 (clone Avas 12a1) by BD PharMingen. Control samples were stained with isotype-matched irrelevant mAb. Data were analyzed using the Summit 4.3.1 computer program and are reported as Log fluorescence intensity versus number of cells.
Statistical analysis
All values are expressed as mean±standard error. Statistical differences between groups were assessed by the use of unpaired t-test and Mann–Whitney U-test. We considered a P value<0.05 to be significant.
Results
Vascularization of flap sectors
Blood perfusion through the flap was evaluated at LDI by measuring the PU average of each sector and was expressed as fold increase/decrease with respect to the preoperative baseline assumed to be equal to 1 (Fig. 2B). As a confirmation of the correctly executed ligation of the epigastric neurovascular bundle, in the immediate after the operation the vital zone of the flap was perfused as before the operation, while in the border zone and even more in the ischemic zone the perfusion was strongly decreased. In the vital zone the perfusion was doubled starting from day 1 and remained steady through the whole week without significant differences between the control and the treated animal groups. This increased perfusion in the zone of the flap vascularized by the not ligated artery (vital zone) must be considered compensative of the absence of vascularization in the flap ischemic counterpart. In the border zone a similar hyper-reaction occurred at day 1, but the increasing trend of the perfusion was maintained through the week only in the ACM animal group. At day 7 the vital and the border parts of the CTRL-treated flaps showed a reduced hyperperfusion effect, while the same parts of the ACM-treated flaps continued to show an increased perfusion effect. We conclude that the observed effect was treatment-dependent rather than a physiological response of the body consequent to the ongoing injury. In the ischemic zone, the CTRL membrane never recovered the initial perfusion (baseline level), while in the same ACM-treated zone the perfusion level at day 7 was 50% higher than the initial baseline. We concluded that, although the simple injury (ligation of the epigastric bundle) triggered an angiogenic response, this was not sufficient to re-vascularize an ischemic area. On the contrary, the ACM treatment mediated the growth and the sprouting of new vessels toward the ischemic regions.
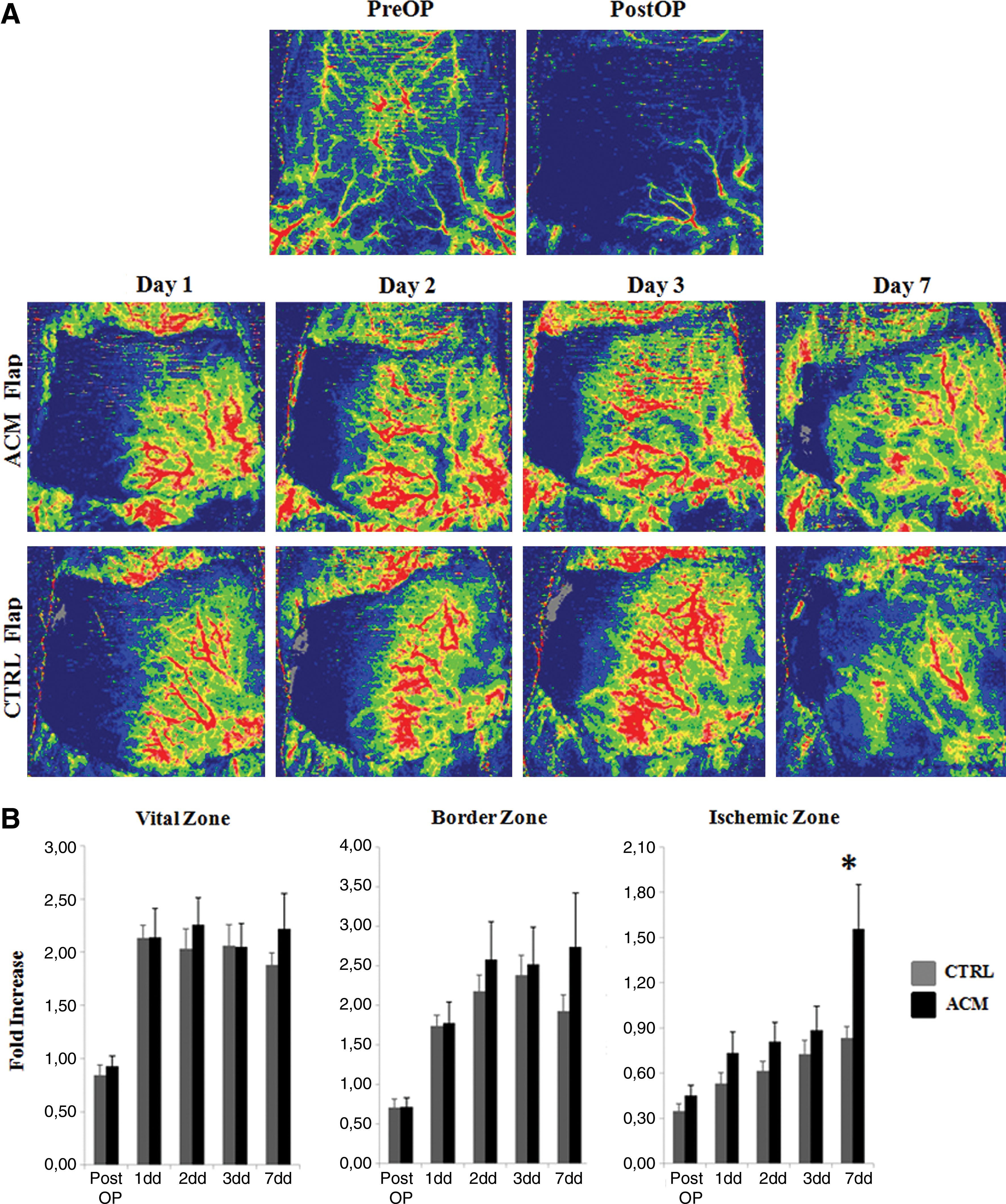
Flap perfusion.
Necrosis development through the flap
We measured the necrotic areas developed through the flap and calculated the percentage (%) of necrosis to the entire flap area. Necrosis started to be evident in the ischemic (ligated) part at day 1 for the flaps treated with the CTRL membrane and at day 2 for the ACM-treated flaps (Fig. 3). Once started, necrosis spread through the flap toward the midline and the CTRL membrane failed to stop its development. The survival of the flap is linked to the blood supply and, looking at the perfusion data (Fig. 2) of the ischemic zone, one can notice that in the CTRL group the perfusion increased very slowly during the week, never getting to the baseline. On the contrary, in the ACM group the ischemic zone suffered a less severe blood supply deficiency during the first 3 days postsurgery and recovered a complete perfusion at day 7 (Fig. 2). Indeed, in the CTRL group, where the ischemic zone was never recovered, the necrosis % was progressively increasing, while in the ACM group, where vessels finally crossed the ischemic part, at day 7 the necrosis% was significantly lower (Fig. 3).
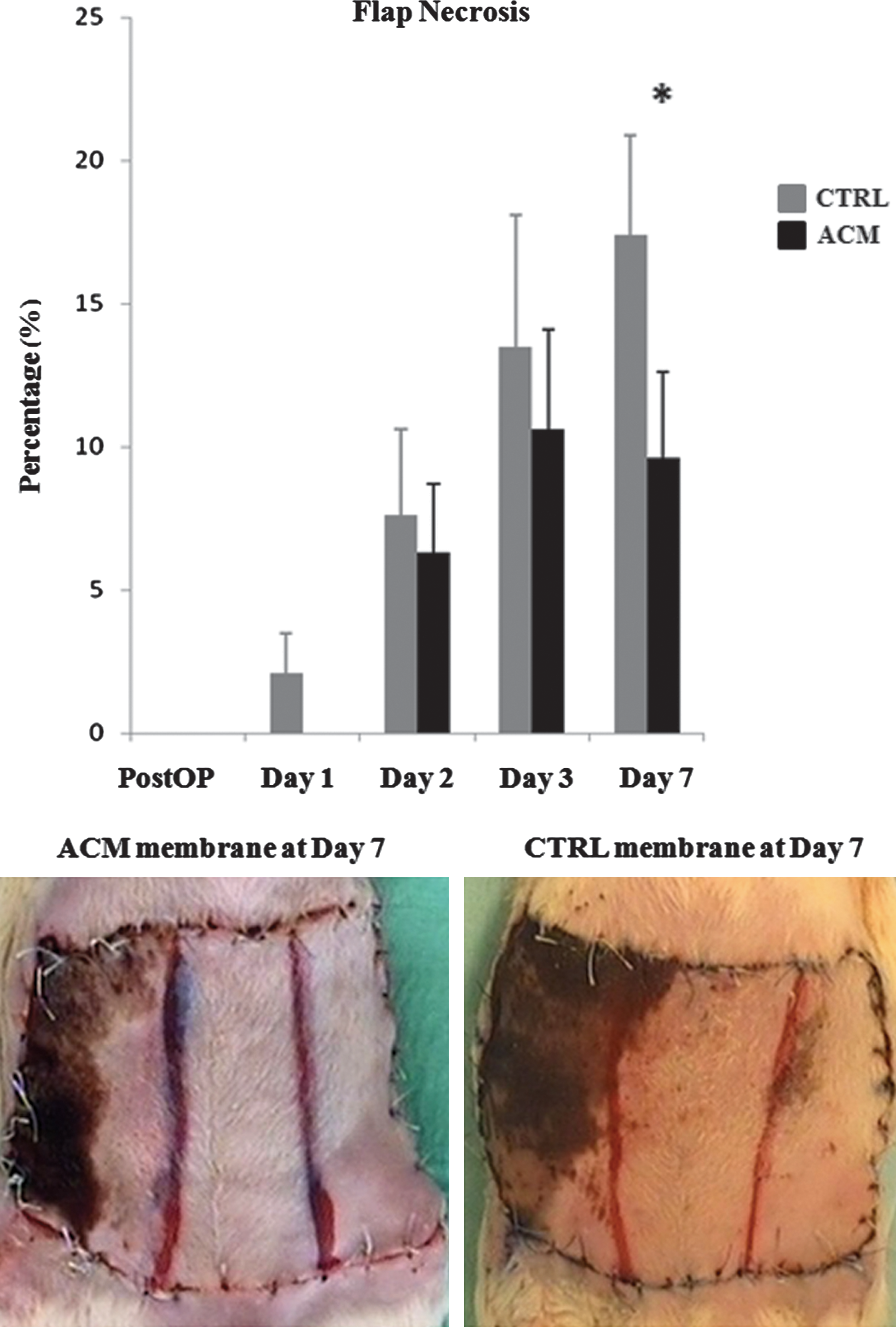
Necrosis development. Histograms represent the percentage of developed necrosis throughout the whole flap area immediately after the operation (postOP) and at different times after ligation of the epigastric bundle. Photos represent a macroscopic view of the ACM and CTRL-treated flap at day 7 postsurgery. *P value<0.05. Color images available online at
Histological analysis of cutaneous and subcutaneous structures
We excised full-thickness skin samples proximally to the necrotic area of flaps. Looking at the cutaneous structures of the CTRL group (Fig. 4A, upper right panel), it is of notice the thickness of the dead upper epidermal layer (orange), the disorganization of the collagen fibers (blue) in the papillary layer, and the dermal glands (pale pink) that appears less numerous and atrophied. Instead, the ACM-treated group (Fig. 4A, upper left panel), showed some detached dead stratum corneum, a normal epithelialization of the stratum spinosum and basale, and a correct arrangement of the collagen fibers and glands in the underlying dermis. In the subcutaneous tissues (Fig. 4A lower panels) where hypoderm and underlying fascia are visible, the vessel density was about 5-folds increased in the ACM group with respect to the CTRL group (Fig. 4B), and the presence of inflammatory infiltrates. The high vascularization, histologically detected in the ACM-treated subcutis, well explain the survival of the upper structures of the skin. Thickness distribution curves of dead cutaneous layers (Fig. 4C) are right side-located in the CTRL group because necrotic layers are thicker in not-treated rats, ranging from 300 to 700 μm. Instead, necrotic layers of ACM group have a distribution curve averaged on 35–100 μm (50% of layers).

Histological analysis.
Recruitment of endothelial progenitors
The flap area we chose for these studies is showed in Fig. 1E and described in Materials and Methods section. Subcutaneous tissues were excised from each animal and digested to obtain a single cell suspension (cells freshly isolated). Cells suspensions were analyzed at the cytofluorimeter for the presence of endothelial cell(EC) and endothelial progenitor cell (EPC) markers [19 –21], such as CD31, CD34, and Flk1 (VEGFR2) (Fig. 5). Cells isolated from ACM-treated flaps included about 10% of CD31, 23% of VEGFR2, and 15% of CD34-positive cells. Further, a double-positive population for CD31/CD34 (0.9% of the total amount of extracted cells) and a double-positive population for CD31/VEGFR2 (3.4% of the total) were isolated from the ACM group.
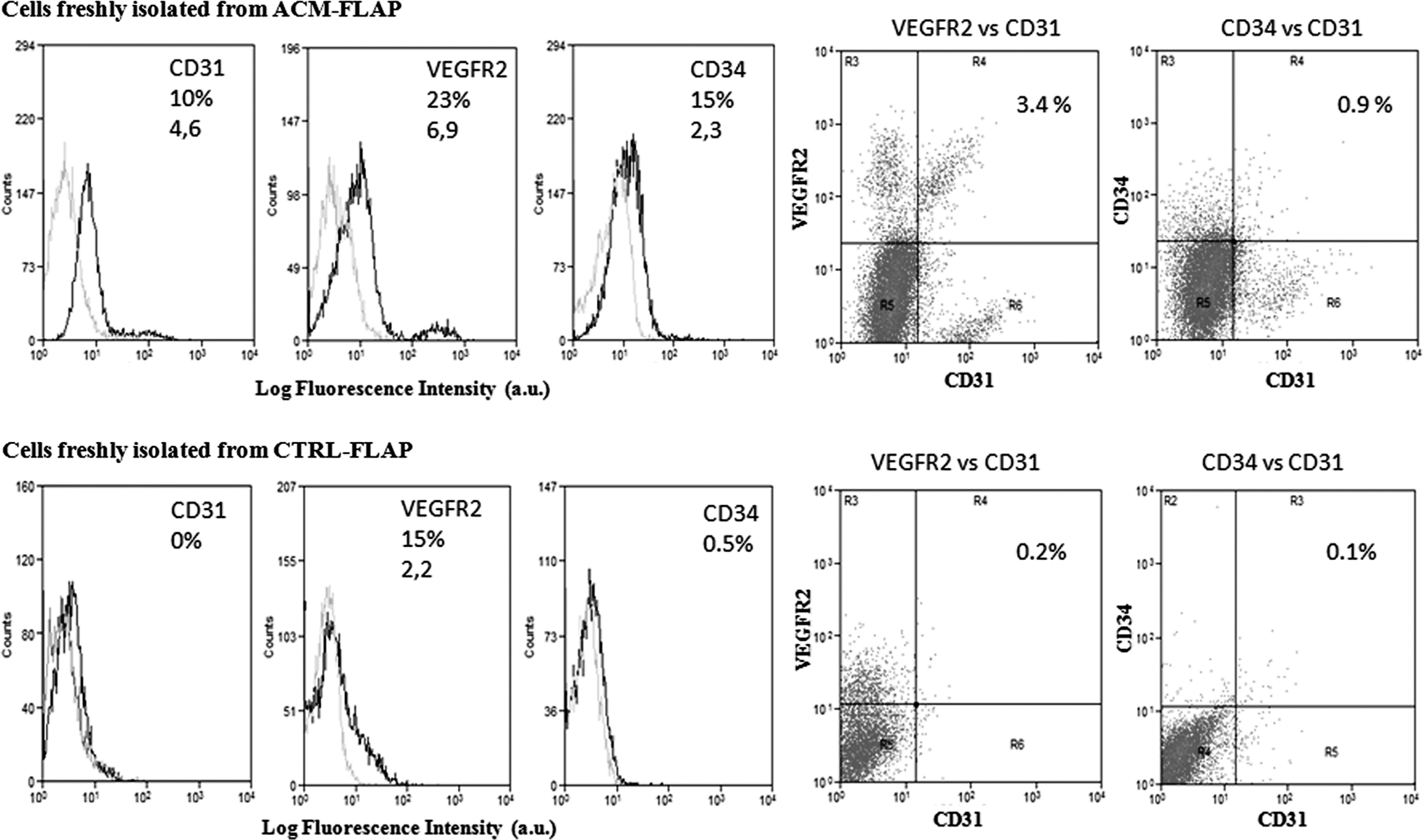
Vascular progenitor recruitment. Cytofluorimetric analysis of cells extracted from the ACM-treated (upper panel) and the CTRL-treated (lower panel) flaps. Histograms on the left show single staining analysis for the markers CD31, VEGFR2, and CD34 (gray histograms are the isotype-matched mAb used as control). Dot charts on the right show double staining analysis for CD31/VEGFR2 and CD31/CD34-positive cells. Percentage of positive cells and mean fluorescence intensity are shown as well. Shown results are representative of independent determinations performed on cells recovered from each animal. Data analysis with Summit 4.3.1 software.
While ACM contains soluble factors capable to chemoattract circulating progenitors and mediate the blood vessels growth, the not treated ischemia alone was not able to recruits EPCs in the subcutaneous tissues of the flap. In fact, cells isolated from the flaps of the CTRL group were negative for the markers CD31 and CD34, and only 15% of them expressed the VEGFR2. When we searched for double-positive cells, only 0.2% of CD31+/VEGFR2+ cells and 0.1% of CD31+/CD34+ cells were observed.
Isolation of endothelial progenitors
Cells derived from the subcutaneous tissues of ischemic flaps were cultured with a 2 steps procedure. First cells were preplated onto fibronectin-coated dishes for 48 h (Fig. 6A) in order to separate adherent from floating cells. Adherent cells extracted from the CTRL group grew slowly, while cells from ACM group got the 70% of confluence in 48 h and presented both polygonal and spindle morphology (Fig. 6A). The nonadherent cells were re-plated at a clonogenic density (25 cells/cm2) on fibronectin-coated dishes. After 7 days of culture colonies were analyzed (Fig. 6B). From ACM group we isolated several colonies (1.7 CFU/cm2) with the typical cobblestone morphology surrounded by radiating thin cells and characterized by a central cluster of rounded cells. These clusters were found positive for the vWF (Fig. 6C), confirming their endothelial nature. We did not obtain colonies from the CTRL group in agreement with the observed absence of double-positive progenitors in the freshly isolated subcutaneous cells from this animal group.
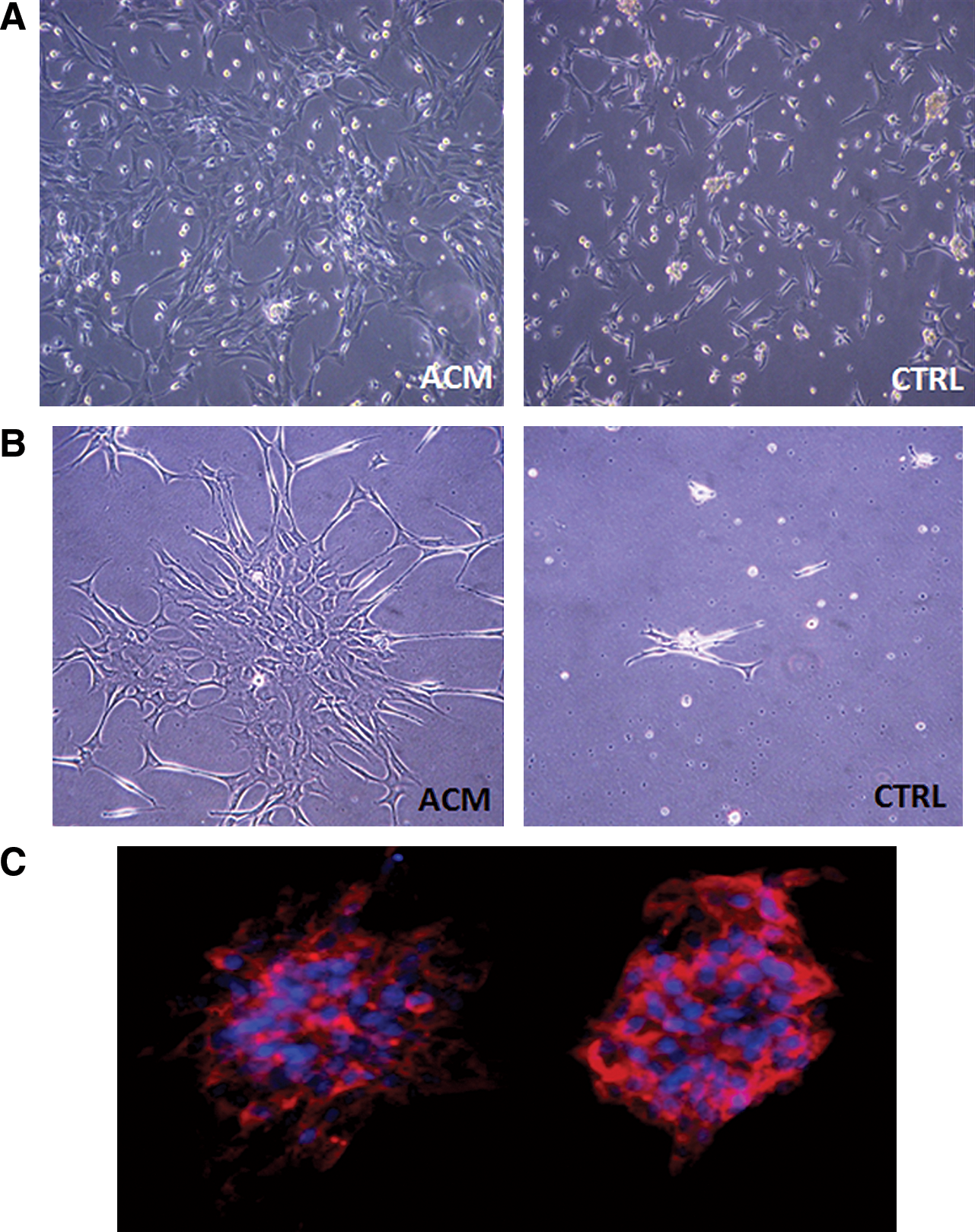
Isolation of endothelial progenitor cells.
Discussion
Ischemia of the operated tissues is a very common and difficult to handle complication of many surgeries. Necrosis of tissues occurs secondary to an early reperfusion injury or a prolonged absence of blood supply. The animal model we chose provides a very serious (half flap-extended and 1 week-protracted) ischemic damage, since the vascularization of the flap is prevented by the chronic epigastric artery ligation and the complete elevation of the whole flap area. Several studies have used angiogenic growth factors [22], in local or systemic administration [23], as recombinant proteins or by gene delivery [24], to pursue a therapeutic angiogenesis of ischemic flaps. We previously demonstrated [14] that the AFSC secretome is responsible for the paracrine vascularizing effect that these cells showed in vivo. We described the conditioned media of human AFSC as a potent cocktail of combined chemokines [such as monocyte chemotactic protein-1 (MCP-1), interleukin (IL)-8, stromal cell derived factor-1α (SDF-1α)] and growth factors (such as VEGF, TGFβ), synergically promoting the neovascularization of ischemic tissues. Here we used 5 mL of 16 h conditioned media (each mL from 800,000 AFSC) for each 25 cm2 gelatin membrane (secretome from 30,000 AFSC per cm2). This low ACM concentration was demonstrated to be effective in this formulation. The membrane we used, offers a good solid support for the local delivery of the ACM at the injury site. This approach was useful to potentiate [10] and at the same time topically limit the neovascularization, thus avoiding clinical concerns linked to a systemic administration of angiogenic growth factors, such as the targeting of nonspecific tissues or, on the other hand, the blood diluting effect and the reduced half-life of circulating proteins.
The support membrane that we chose, made of gelatin, (1) is a biodegradable, biocompatible, and nonimmunogenic product, suitable for many medical applications; (2) its hydrophilic nature enables absorption of ACM; (3) in the adopted formulation it can undergo a gel/sol transition at body temperature thus releasing proteins in a controlled way. The gelatin membrane alone, soaked with saline, was implanted in rats of the CTRL group, where no recovery of the ischemic sector of the flap occurred. Postnatal growth of new vessels is activated during pathological conditions, such as wound healing, tumorigenesis, and tissue ischemia [25]. In our CTRL group the ischemic injury per se triggered a neovascularization, as the compensative hyperperfusion effect in the vital and border zones showed, nevertheless this was not sufficient to re-establish a normoperfusion in the ischemic zone within the first week. The initial baseline level of perfusion was never achieved, the subsequent necrosis reached the 18% of the whole flap area with peaks of 23%, and no progenitors were detected (cytofluorimetric analysis) or isolated (CFU analysis) from the ischemic subcutis.
The postinjury vascularization can be enhanced by the administration of growth factors and cytokines [7] that recruit EPC into the injury site, thus accelerating the wound healing. In the ACM group, at day 7, the ischemic area was re-vascularized with a perfusion level 50% higher than the initial baseline and, as a consequence, presented a less developed necrosis with respect to the CTRL group. The histology of ACM-treated flaps revealed a thin dead stratum corneum, a normal arrangement of the epidermal and dermal structures, and a high density of vessels in the subcutaneous tissues. More important, we showed that ACM recruited CD31+/VEGFR2+ and CD31+/CD34+ cells into the ischemic subcutaneous tissues, where growth and sprouting of new vessels occurred. Since subcutaneous tissues contain blood, adipose cells, and other derivatives of mesodermal origin, we used a double staining analysis for the quantification of endothelial progenitors from the freshly isolated cell population. In fact, CD31 cannot be considered a specific marker of ECs, since it is expressed also by circulating monocytes [26]. Similarly, CD34 cannot be considered a solely marker of EPC since CD34-positive cells can be abundantly isolated also from the total nonendothelial population within the SVF of adipose tissues [27]. Even the VEGFR2 is not a unique marker for hematovascular lineage cells, since it is expressed also by developing and mature mesoderm derivatives [28]. We found that out of the 23% VEGFR2-positive cells and the 15% CD34-positive cells extracted from the ACM-treated flap, 3.4% were CD31/VEGFR2 double positive and 0.91% were CD31/CD34 double positive. To confirm the ACM-mediated recruitment of EPCs, we identified vWF-positive cell clones derived from the free floating cell fraction obtained from the digestion of subcutaneous tissues.
Given the extraction of a high percentage of CD34 single-positive and VEGFR2 single-positive cells from subcutaneous tissues of ACM group, together with the isolation of a high proliferative SVF, one could speculate that the ACM also stimulated stem/progenitor cells residing in the subdermal adipose tissue and dermis [29]. Many studies have demonstrated that a variety of growth factors and cytokines are involved in the cutaneous wound healing; IL-6 is necessary for tissue granulation, VEGF for the wound angiogenesis, TGF-β for the reepithelialization. ACM contain these growth factors [14] together with other soluble factors, such as chemokines (IL-8, MCP-1, SDF-1α) responsible for the migration of inflammatory cells and for the mobilization, homing, and incorporation of endothelial progenitors into the vasculature [30]. Again, since AFSC secrete the mentioned growth factors and chemokines, as our and other groups have demonstrated [14,31 –35], we hypothesize that the endogenous repair triggered by the AFSC secretome could involve both the recruitment of EPC and the activation of local niches.
Conclusion
ACM, as a cocktail of “navigation cues and niche signals” [36], are effective, safe, simple, and reproducible to formulate, convenient, easy to deliver, being good candidate for a translational treatment that could be: (1) in situ-directed; (2) based on the activation of endogenous mechanisms of repair; (3) cell-free, thus avoiding the ex vivo manipulation of autologous or heterologous cells.
Footnotes
Acknowledgments
We thank the Galliera Genetic Bank (Network of Telethon Genetic Biobanks, Genoa, Italy) for providing amniotic fluid cells, Anna Hofmann, Matthias Layr, Sabine Pfeifer, Rainer Mittermayr (Ludwig Boltzmann Institute for Experimental and Clinical Traumatology, Vienna, Austria) for developing the animal model, and Dr. Roberto Benelli (National Institute of Cancer, Genoa, Italy) and Prof. Heinz Redl (Ludwig Boltzmann Institute for Experimental and Clinical Traumatology, Vienna, Austria) for the very kind scientific support. Financial support by research funding from the EU's Seventh Framework Program (Angioscaff).
Authors Disclosure Statement
None to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.