
Abstract
Mesenchymal stem cells (MSC) have been isolated from several adult human tissues. Their propensity to differentiate into cell types of connective tissue, such as osteocytes, chondrocytes, and adipocytes, suggests that MSC may function as a reserve of progenitor cells that repair and maintain healthy adult tissues. Dysfunction of the trabecular meshwork (TM), a connective tissue at the anterior region of the human eye that regulates intraocular pressure, plays a major role in the pathogenesis of glaucoma. The mechanobiology and pharmacological aspects of the TM tissue have been relatively well studied in disease states. Less well understood is if there are progenitor cells within the TM that contribute to maintenance of this tissue. In this study, we have identified and characterized an expandable population of cells that have stem cell-like properties. In particular, these cells express the markers CD73, CD90, and CD105, which are typically associated with MSC. Thus, we have named these cells TM-MSC. As further evidence that these cells are MSC, they were differentiated in vitro into adipocytes, osteocytes, and chondrocytes. Through genomic characterization, we show that TM-MSC have gene expression patterns most similar to MSC derived from other tissues. TM-MSC express genes found on adult TM tissue, suggesting that TM-MSC are progenitor cells that serve to maintain a healthy TM.
Introduction
G
In contrast to the filtering portion of the TM, the nonfiltering Schwalbe's line cells are believed to function differently—previous studies have suggested that these cells possess progenitor-like properties [2 –4]. The importance of resident stem/progenitor cell populations within the eye has been well investigated, beginning with the identification of corneal limbal epithelial stem cells [5], which may hold promise in providing a self-renewing population of stem cells for corneal transplantation. Besides limbal stem cells, other stem cell types within the eye have been identified [6 –9]. That stem/progenitor cells exist in the TM and serve to maintain proper functioning of the TM is an attractive hypothesis. Therefore, the aim of this study was to identify and characterize a specific population of self-renewing progenitors within the TM that could provide a transplantable source of normal TM cells as a possible treatment strategy for patients with glaucoma. In this study, we have isolated and propagated TM-derived mesenchymal stem cells (TM-MSC). Importantly, we demonstrate that (1) TM-MSC resemble MSC morphologically and express MSC markers; (2) TM-MSC are multipotent, capable of differentiating into adipogenic, osteogenic, and chondrogenic lineages; and (3) TM-MSC are more similar to MSC than other cell types as indicated by gene expression signatures.
Materials and Methods
Isolation of human TM-MSC
Anterior eye segments were obtained from the Lions Eye Bank (Tampa) or from the Singapore National Eye Centre, following the institutional ethics board's guidelines. The corneoscleral rims were rinsed thrice in wash buffer (50% phosphate-buffered saline) with calcium- and magnesium-containing 25% Opti-MEM® 1, 25% human endothelial-SFM, Gentamycin reagent solution (25 μg/mL), penicillin–streptomycin (1×), and amphotericin B (1×) (all from Invitrogen). The rims were stained with 0.4% trypan blue (Invitrogen) for 1 min to visualize the TM and corneal endothelium, and then rinsed with phosphate-buffered saline (PBS). TM tissue was isolated from the iridocorneal angle using a technique adapted from Tripathi and Tripathi [10]. Under a stereomicroscope, the darkly stained corneal endothelium was peeled off and discarded, while the adjacent TM was gently separated from the underlying tissue using a crescent blade. This technique ensured that surrounding structures such as the corneal endothelium and scleral stroma were avoided. The meshwork was digested overnight in 2 mg/mL type I collagenase (Worthington Biochemical Corporation) dissolved in Dulbecco's modified Eagle medium (DMEM) containing 10% fetal bovine serum (FBS).
Culture and propagation of TM-MSC
The digested TM was further dissociated with 0.05% trypsin (Invitrogen; catalog no. 25300054) or Accutase (Millipore; catalog no. SCR005) to achieve a single-cell suspension. Where necessary, the cell suspension was triturated to aid in dissociation. The TM-MSC were cultured and propagated in DMEM (low glucose), containing 10% FBS, 4 mM L-GlutaMAX™, 1 mM sodium pyruvate, 1% nonessential amino acids, and 1% penicillin–streptomycin, on uncoated tissue culture plastic. Passaging was carried out using 0.05% trypsin–ethylenediaminetetraacetic acid.
Colony forming unit-fibroblast assay
The colony forming unit-fibroblast assay was performed as described [11]. Briefly, primary TM-MSC (at passage 0) were seeded at a density of 3,000 cells/well in a 24-well tissue culture plate and cultured in the MesenCult Proliferation Kit (Human) containing the MesenCult Basal Medium plus Stimulatory Supplements (StemCell Technologies). After 14 days, the cells were fixed for 5 min in methanol and stained in Giemsa (Sigma Fluka) for 5 min. The plate was rinsed, and the number of colonies under the brightfield microscope was enumerated and expressed as %-colony forming unit-fibroblast (CFU-F) [i.e., (number of CFU-Fs/3,000) ×100%].
Flow cytometry
Flow cytometric analyses were performed on TM-MSC that were isolated and cultured separately from pairs of eyes of 3 donors at passages 4–6. The harvested cells were washed in PBS, resuspended at the concentration of 106 cells/mL in an ice-cold PBS containing 10% FBS, and incubated for 30 min at 4°C. About 100 μL of the suspension was treated with conjugated antibodies for CD11b, CD34, CD45, CD73, CD79α, CD90, CD105, and HLA-DR at dilutions recommended by the manufacturer for 45 min at 4°C. Isotype IgG was used as a negative control. All antibodies were purchased from BD Pharmingen, except CD105, which was purchased from Abcam. For detailed information on the antibodies, see Supplementary Table S1 (Supplementary Data are available online at
Differentiation toward adipocytic, chondrocytic, and osteocytic lineages
Differentiation toward the 3 lineages was done in parallel on TM-MSC at a low-passage (p1 to p4) split from the same plate and were cultured up to 80%–90% confluence before induction. To induce adipogenic differentiation, the media were changed to the MesenCult Basal Medium with MesenCult Adipogenic Stimulatory Supplements (StemCell Technologies, 10% final volume) and cultured for 4 weeks, with thrice weekly media changes. Before fixing, TM-MSC were washed once with PBS. Fixation was conducted using 10% neutral-buffered formalin at room temperature for 30 min. The formalin was then aspirated and the well was rinsed twice with sterile water. After the rinse, 60% isopropanol was added, and the cells were incubated for 5 min. The isopropanol was aspirated, and the Oil Red O working solution of 1.8 mg/mL was added for 30 min. The stain was removed by rinsing with water, and red lipid vesicles were visualized under phase contrast.
For osteogenic differentiation, TM-MSC were cultured in osteogenic differentiation media, consisting of the MesenCult Basal Medium with MesenCult Osteogenic Stimulatory Supplements (StemCell Technologies), dexamethasone (10−8 M), ascorbic acid (50 μg/mL), penicillin/streptomycin (1×), and gentamycin (50 ng/mL). Media were changed thrice a week. When multilayering was observed, osteogenic differentiation media containing β-glycerophosphate (StemCell Technologies; 3.5 mM) were added. After 4 weeks, cells were washed once with PBS and fixed with absolute ethanol for 30 min at room temperature. The ethanol was aspirated, and the wells were washed twice with water, before Alizarin Red (Sigma) was added for 30 min. The stain was rinsed off, and the stained cells were visualized under a brightfield microscope.
For chondrogenic differentiation, TM-MSC were trypsinized, and then aggregated as pellets containing 2×105 cells by centrifuging at 450 g, according to previous published protocols [12, 13]. The pellets were cultured in chondrogenic differentiation media containing the MesenCult Basal Medium, insulin–transferrin–selenium (ITS; Invitrogen; 1 mM), bovine serum albumin (PAA Laboratories; 1.25 μg/mL), linoleic acid (Sigma; 5.35 μg/mL), sodium pyruvate (Invitrogen; 1 mM), dexamethasone (Sigma; 10−7M), ascorbic acid (StemCell Technologies; 50 μg/mL), transforming growth factor beta 3 (TGF-β3; Peprotech; 10 ng/mL), gentamycin (Invitrogen; 50 ng/mL), and penicillin/streptomycin (Invitrogen; 1×). Media were changed thrice a week. After 5 weeks, the pellets were processed for histology and stained with type II collagen.
Immunocytochemistry and immunohistochemistry
TM-MSC were rinsed twice with PBS and fixed with 4% paraformaldehyde (PFA) on ice for 15 min. The PFA was aspirated, and the cells were washed 3 times with PBS containing 0.1% Tween®20 (PBT; Sigma) to permeabilize the cell membranes. The cells were then blocked for an hour at room temperature with PBT containing 10% FBS. The primary antibody was diluted in block. Antibody details are listed in Supplementary Table S1. Cells were incubated with the primary antibody overnight at 4°C, and then washed 3 times with PBS for 5 min each. Secondary antibodies were purchased from Invitrogen and details are listed in Supplementary Table S1. Cells were incubated with a secondary antibody diluted in block at room temperature for 1–2 h, and then washed with PBS thrice for 5 min each. A nuclear stain, 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen), was applied to the cells for 5 min, and then washed thrice with PBS. Where cells were cultured on coverglass slides, the slides were mounted in fluorescence mounting media (Dako) and viewed under a Zeiss Axio Observer D1 fluorescence microscope with a Zeiss AxioCam MRm high resolution microscope camera. Images were analyzed under Zeiss AxioVision 4.6 image analysis software and processed in Adobe Photoshop (Adobe).
Chondrogenic pellets were fixed with 4% paraformaldehyde at 4°C and embedded in wax blocks for sectioning into 5-μm sections. Slides were deparaffinized and rehydrated before staining. Epitope retrieval was achieved using a citrate buffer at pH 6.0 for 40 min at 100°C. Endogenous peroxidase was blocked with 3% hydrogen peroxide for 15 min at room temperature. Slides were treated with serum block for 30 min before incubation in collagen II primary antibody (Abcam; 1:500) for 45 min at room temperature. Horseradish peroxidase anti-rabbit secondary was applied for 10 min before detection with diamino benzidine for 3 min. The slides were visualized under a Leica DM LB2 light microscope; pictures were taken using a Leica DFC 320 camera and analyzed using Leica Application Suite software (Leica Microsystems GmbH).
RNA isolation, reverse transcription, and polymerase chain reaction
Total RNA was isolated using Trizol (Invitrogen) or the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems) according to the manufacturer's instructions, and precipitated with 70% ethanol. Reverse transcription of RNA into cDNA was conducted using the Superscript III First-Strand Synthesis System (Invitrogen) according to manufacturer's instructions. Reverse transcriptase–polymerase chain reaction (RT-PCR) and quantitative RT-PCR (qRT-PCR) primers were designed by Perl Primer software to be intron–exon spanning and, where possible, to have an annealing temperature of 55°C (Supplementary Table S2). Primers were synthesized by 1st Base Oligos (Singapore). cDNA was amplified in over 30 cycles (RT-PCR) and 40 cycles (qRT-PCR). RT-PCR was done in a MyCycler or a Tetrad2 PCR machine (both from Biorad), and qRT-PCR was done on the 7900HT Fast Real-Time PCR system and analyzed using SDS2.2.2 software (both from Applied Biosystems). The ΔΔCT method of comparative quantification was used. Markers were tested on 3 biological replicates; for quantitative RT-PCR, additional 2–3 technical replicates were tested.
Microarray and gene set enrichment analysis
RNA from TM-MSC was processed for microarray studies according to manufacturer's instructions (Ambion/Applied Biosystems). Complementary RNA (cRNA) resulting from the RNA amplification and purification steps above was hybridized onto Illumina HumanRef version-2 microarray chips (Illumina Inc.). The chips were scanned on the Illumina BeadArray Reader using Illumina BeadScan software (both Illumina), and the expression data were processed by BeadStudio (Illumina) followed by GeneSpring GX (Agilent). Housekeeping genes were filtered by using in-house scripts. TM-MSC were compared to a customized Gene Set Enrichment Analysis (GSEA) gene set comprising other types of stem cells, primary cell culture, and primary tissue gene sets, including bone marrow-derived MSC (BM-MSC), cortical bone, periosteum, auricular cartilage, neural progenitor cells, fibroblasts, smooth muscle, and eye-specific gene sets such as corneal derivatives and scleral tissue. This was done by first ranking the TM-MSC versus scleral fibroblast data set that was generated from our microarray results, according to expression (henceforth referred to as the TM-MSC data set). The ranked data set was then compared against published gene sets in Gene Expression Omnibus (GEO), using the GSEA program [14]. Gene sets from GEO used in this analysis are listed in Supplementary Table S3, and were compared against the TM-MSC data set for any enrichment. GSEA was performed using GSEA software (Broad Institute, MIT). The GSEA program calculated the normalized enrichment score (NES), nominal P-value, and the false-discovery rate q-value. Our data are deposited under GSE28679.
Statistical analysis
Data were expressed as mean±standard error of the mean (mean±SEM), and analyzed using unpaired 2-tailed equal variance t-tests unless otherwise specified.
Results
Isolation and propagation of an MSC-like cell type from human TM
We sought to culture stem cells isolated from the TM of the human eye. Human TM tissue was micro-dissected from the iridocorneal angle of cadaver eyes using an adaptation of previously reported methodology [10]. Digestion overnight in collagenase and subsequent trituration of the TM tissue, followed by further digestion with trypsin, resulted in a single-cell suspension. The TM cell suspensions were cultured under several conditions, including methods that favored propagation of embryonic stem cells and MSC under various adherent and suspension conditions. We observed clusters of cells growing in suspension conditions that resembled spheres, a cell type described previously [15], but we were unable to expand this population of cells (data not shown). However, under adherent culture conditions and, importantly, using serum that was qualified for human embryonic stem cell (hESC) growth, we were able to reproducibly expand a population of cells that had a distinctive morphology, one that is typical of mesenchymal stem cells [16]. We refer to this population of cells as TM-MSC and performed more detailed analyses of their MSC-like characteristics.
The TM-MSC we isolated adhered well to tissue culture plastic and consistently displayed a spindle-like morphology with elongated and smooth cell borders (Fig. 1A), similar in appearance to MSC derived from adipose tissue (hAd-MSC) (Fig. 1B). The nucleus was distinctive and positioned centrally, and the nuclear-to-cytoplasmic ratio of TM-MSC was somewhat variable. In contrast to published methods of culturing cells from TM explants on glass coverslips, the TM-derived cells we seeded directly onto uncoated tissue culture plastic were quickly established, typically within 2 to 7 days (data not shown). Cultured TM-MSC were routinely passaged for at least 10 passages, after which senescence was observed. At passage 4, the doubling time was about 60 h when grown on uncoated plates, with confluence reached within a week of seeding. At passage 8, confluence was typically observed after 2 weeks or in some cases longer with the same plating density (data not shown).
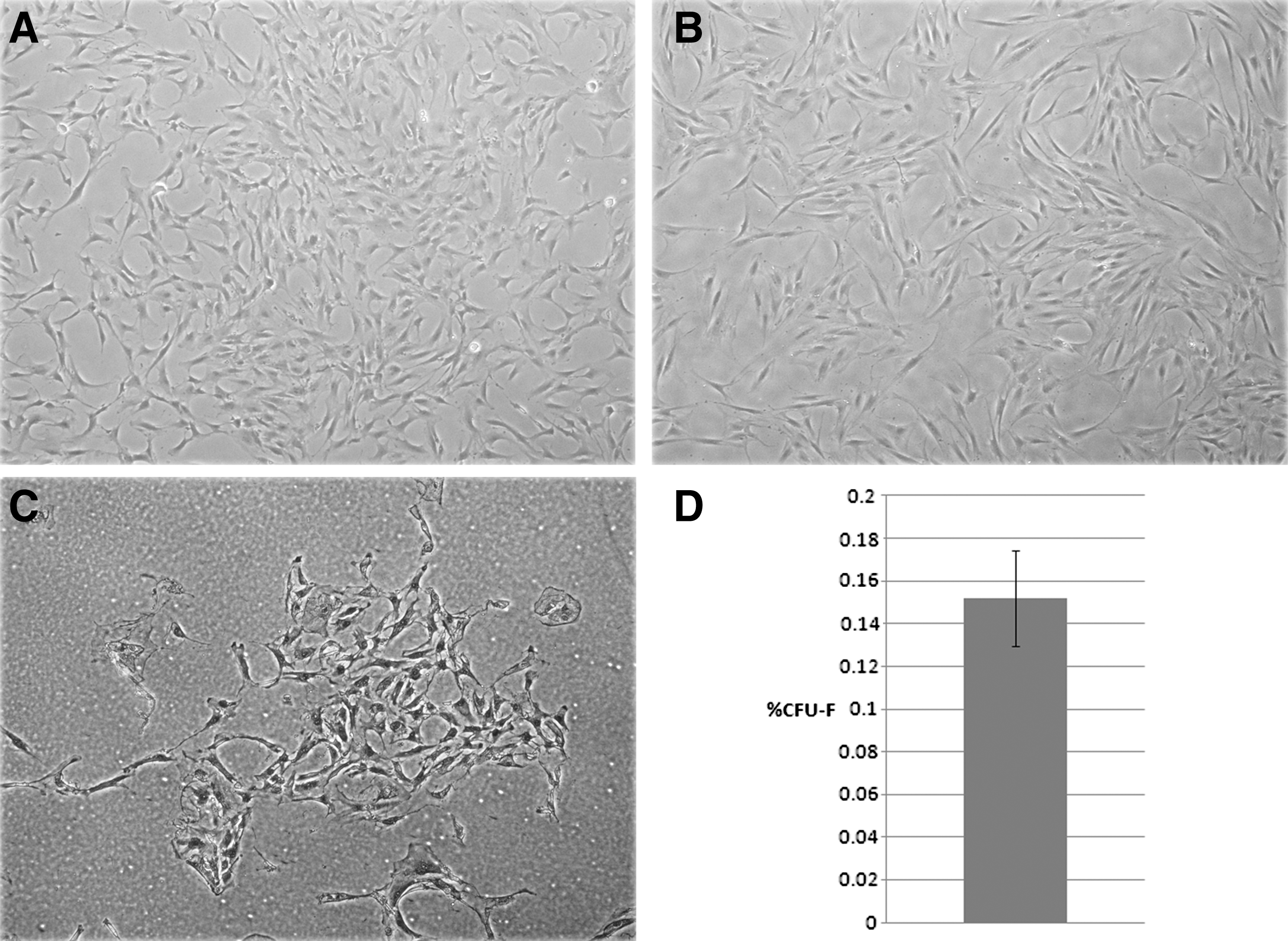
Morphology of early-passage TM-MSC (p2)
It has been demonstrated that a population of self-renewing fibroblast-like cells can be isolated from bone marrow stroma, termed “colony-forming units-fibroblast” (CFU-F), which are capable of differentiating into the nonhematopoietic tissues within the bone marrow stroma [11]. The CFU-F assay, a method of determining the frequency of stem/progenitor cells within a primary cell population, was undertaken on TM-MSC that had been freshly isolated from TM tissue before passaging. TM-MSC seeded at low densities [3,000 cells per well of a 12-well plate (3.8 cm2)] generated colonies after 14 days (Fig. 1C), indicative of the presence of proliferative cells within the population (Fig. 1D). Across 6 biological samples of TM-MSC analyzed for CFU-F formation, the mean frequency of CFU-F formation was 0.152%±0.022% SEM (Fig. 1D). Further investigation revealed that there was no significant correlation of CFU-F frequency with donor age (data not shown).
TM-MSC express mesenchymal stem cell markers
Based on our observations that TM-MSC morphologically resembled MSC (Fig. 1A, B) and that they were potentially a stem cell population with proliferative capacity (Fig. 1C, D), we analyzed TM-MSC for their expression of the MSC markers: CD73 (NT5E), CD90 (THY-1), and CD105 (endoglin). Expression of these markers has been widely adopted as the minimal criteria of defining MSC [17]. By flow cytometry, most TM-MSC expressed CD73 (99%), CD90 (93%), and CD105 (95%) (Fig. 2A). Similarly, by immunocytochemistry, nearly all the TM-MSC stained uniformly positive for CD73, CD90, CD105, and CD146 (Fig. 2B). When co-labeled with CD105 and CD90, TM-MSC were double-positive for both MSC markers (Fig. 2B). We also tested TM-MSC for markers that would not be expected to be expressed on MSC. Indeed, this was the case as TM-MSC were negative for a leukocyte marker (CD11b), a hematopoietic stem cell marker (CD34), a pan-hematopoietic lineage marker (CD45), and a B-cell marker (CD79α), and were low (2%) for major histocompatibility antigen (HLA-DR) (Fig. 2A).
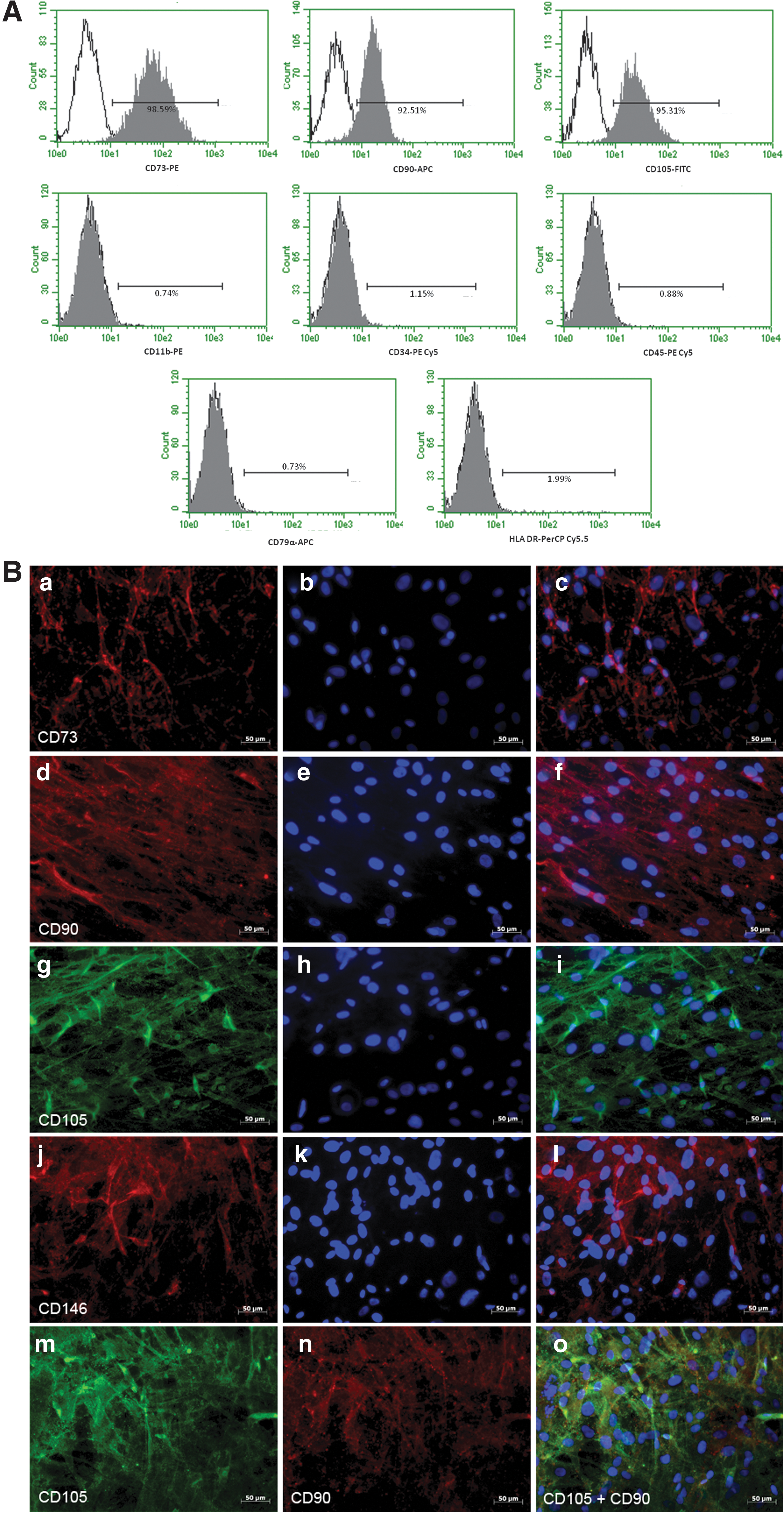
Marker expression in TM-MSC was confirmed by RT-PCR (Supplementary Fig. S1). We observed that TM-MSC, like hAd-MSC, also expressed several other stem cell markers such as MYC, NANOG, OCT4, KLF4, REX1, and NESTIN, but not SOX2 (Supplementary Fig. S2). Taken together, these results indicate that TM-MSC meet the established criteria of marker expression defining an MSC population.
TM-MSC are multipotent, being able to differentiate into adipocytes, osteocytes, and chondrocytes
The marker expression data indicated that the TM-MSC were similar to MSC. Thus, we investigated whether TM-MSC were capable of multilineage differentiation, a defining characteristic of all MSC. To this end, low-passage TM-MSC were differentiated under adipogenic, chondrogenic, and osteogenic conditions. Although these widely used methods do not produce homogenous populations of these cells types, they are, nonetheless, useful to establish the potential of MSC to give rise to such lineages.
Under conditions that favored adipocyte differentiation [18], TM-MSC gave rise to Oil Red O–positive cells that tended to cluster together (Fig. 3A). In addition, adipogenic markers peroxisome proliferator-activated receptor gamma (PPARγ), fatty acid binding protein 4 (FABP4), and lipoprotein lipase (LPL) were upregulated in TM-MSC differentiated under adipogenic conditions, compared to undifferentiated control TM-MSC (Fig. 3B).
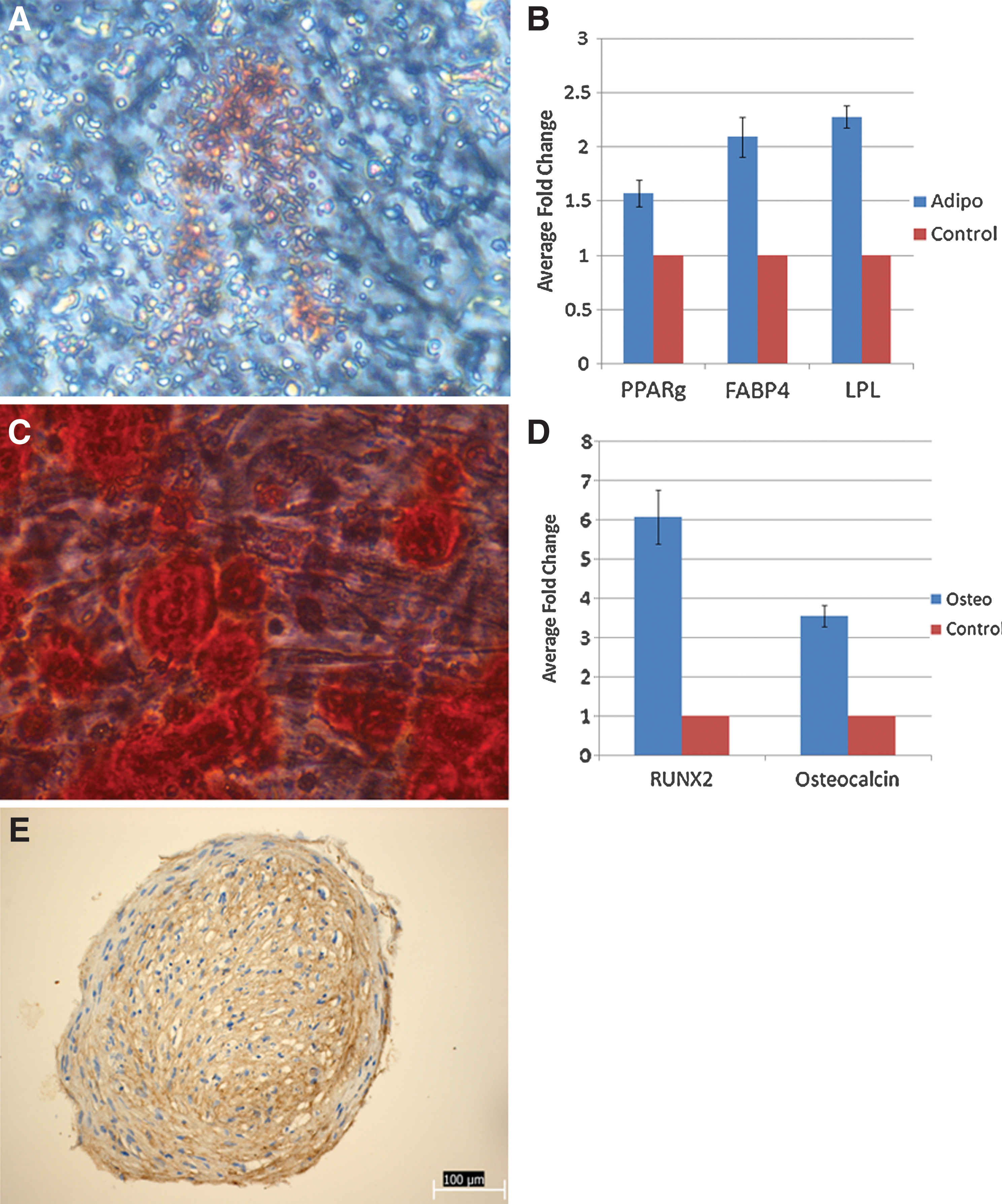
Multilineage differentiation of TM-MSC into the adipogenic
Osteocytic differentiation [19] of TM-MSC was conducted over 4 weeks, and the TM-MSC were subsequently stained with Alizarin Red for the presence of intracellular calcium deposits (Fig. 3C). Clusters of differentiated (Alizarin Red-positive) TM-MSC were visible with the naked eye (data not shown) as well as under the microscope (Fig. 3C). When analyzed by qRT-PCR, osteogenic markers RUNX2 and osteocalcin were seen to be upregulated in TM-MSC differentiated under osteogenic conditions (Fig. 3D).
To analyze the potential of TM-MSC to become chondrocytes, TM-MSC were pelleted in an aggregation culture assay as previously described [12, 13]. After 5 weeks, pellets were collected and stained for type II collagen, a general chondrocytic marker [20, 21] (Fig. 3E). The type II collagen staining appears to be stronger in the inner core rather than in the outer fibrous layer.
In summary, our differentiation results demonstrate that TM-MSC are multipotent stem cells that give rise to the adipogenic, osteogenic, and chondrogenic lineages. Taken together with the marker expression pattern of these cells, it can be concluded that these cells derived from TM possess the defining characteristics of MSC.
Gene expression profiling of TM-MSC
To test our hypothesis that TM-MSC are similar to MSC, we conducted a detailed microarray study. Gene expression profiles were generated for TM-MSC and compared to several other cell types. Briefly, our TM-MSC expression data were rank-ordered according to differential expression values and compared to published gene sets using the GSEA program [14]. Gene sets used for comparison were selected from a wide variety of stem and somatic cell types, including bone marrow-derived MSCs (BM-MSC), cortical bone, periosteum, auricular cartilage, neural progenitor cells, fibroblasts, smooth muscle, and eye-specific gene sets such as corneal derivatives and scleral tissue. Figure 4A shows a graph of the various stem/somatic cell and tissue types arranged according to how similar they were to our TM-MSC data set. The P-value denotes the family-wise error rate computed by the GSEA program and corrected for multiple hypotheses testing. For clarity, the dotted red line on the graph represents the threshold by which values above the line are considered significant (P<0.05), and values below the line are considered not significant (P>0.05). Based on our GSEA results, we found that 3 separate BM-MSC gene sets were significantly enriched in TM-MSC (P<0.05) (Fig. 4A). In addition, gene sets representing mesenchymal-derived tissues such as cortical bone, periosteum, and cartilage were significantly enriched in the TM-MSC data set, indicating that the TM-MSC share some similarity to tissues of mesenchymal origin. Interestingly, gene sets derived from corneal derivatives (such as limbal epithelial stroma and corneal tissue) were not significantly enriched in our TM-MSC data set. Since MSC/mesenchymal gene sets were enriched significantly in TM-MSC, we conclude that TM-MSC are more similar to mesenchymal derivatives than they are to corneal limbal epithelium or neural progenitors.
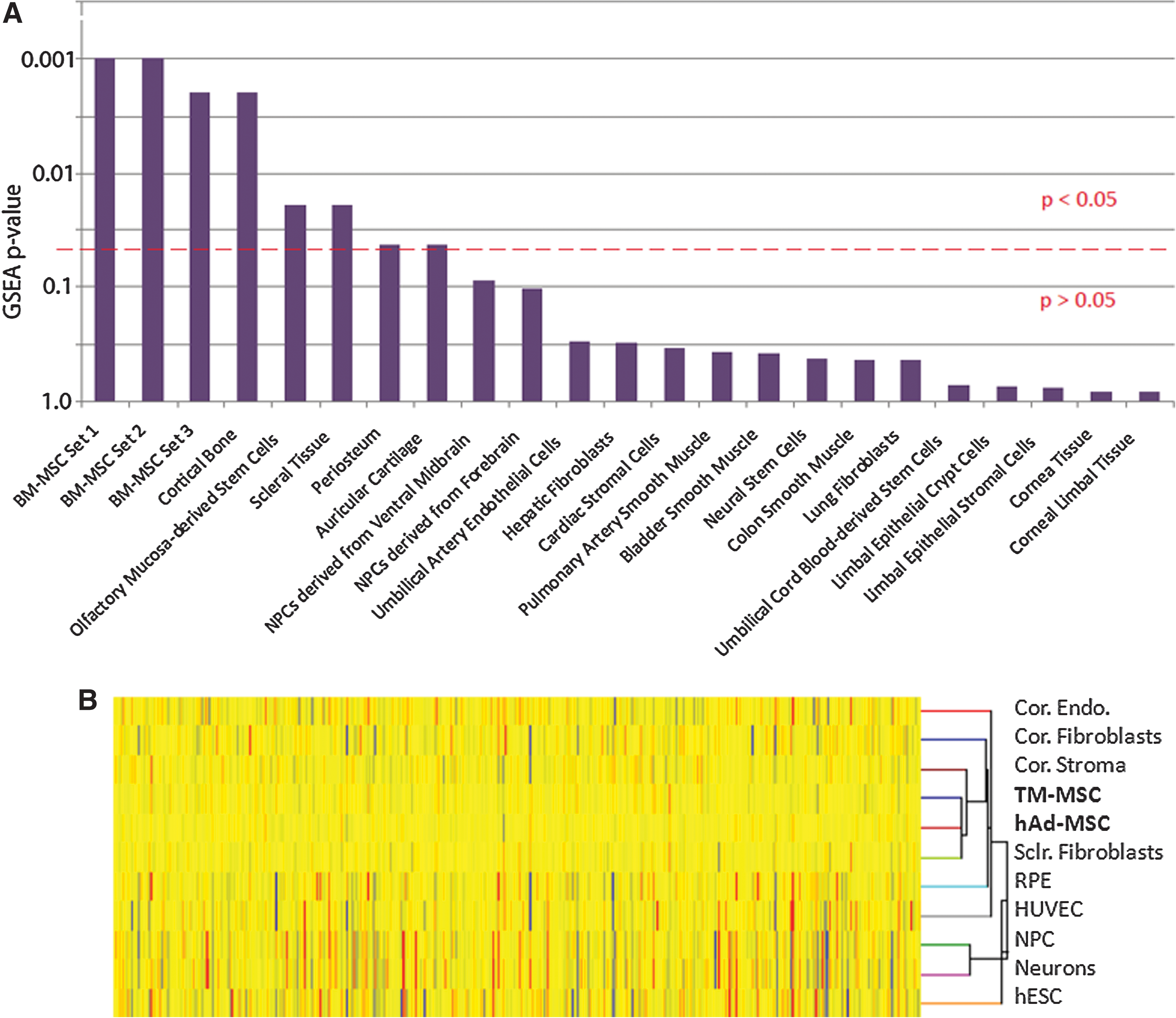
Microarray and GSEA analyses of TM-MSC.
To further investigate the similarities between the TM-MSC populations and other cell types that are not available in the GEO database, we conducted a microarray study comprising other cell types such as corneal fibroblasts, retinal pigmented epithelial (RPE) cells, corneal stroma, hESC, and neurons differentiated from hESC. The dendrogram in Fig. 4B shows the hierarchical clustering of TM-MSC with these cell types. Of note is that TM-MSC cluster with hAd-MSC and scleral fibroblasts, demonstrating that TM-MSC are similar to these 2 cell populations. TM-MSC appear to be less related to corneal fibroblasts, RPE, and share even less similarity to human umbilical cord endothelial cells, hESC, neural precursor cells, and mature neurons. Taken together, our results demonstrate that by GSEA and hierarchical clustering (1) TM-MSC share similarity with mesenchymal gene sets and (2) TM-MSC share little similarity with derivatives from other parts of the eye, such as from the cornea and retina.
TM-MSC express markers of the TM
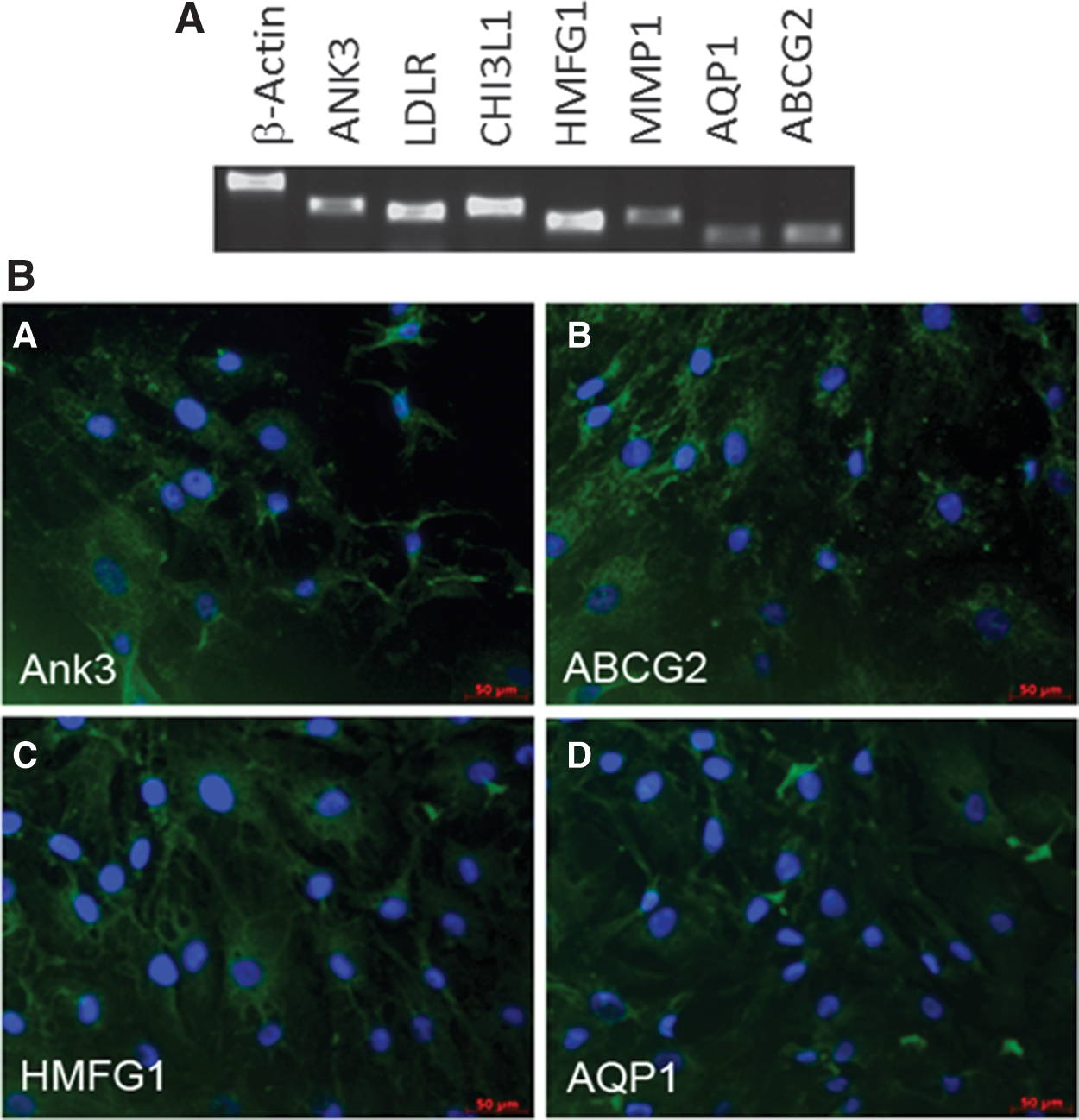
The isolation of MSC from TM tissue raises the distinct possibility that these cells can give rise to TM tissue. We hypothesized that if TM-MSC are progenitors of TM, they would express certain genes also found in mature TM cells. Thus, we examined low-passage TM-MSC for expression of known TM markers (Fig. 5). By RT-PCR analysis, we detected expression of several previously reported TM markers, including Ankyrin3 (ANK3), Low Density Lipoprotein Receptor (LDLR), Chitinase3-Like-1 (CHI3L1), Human Milk Fat Globule Globule 1 (HMFG1), Matrix Metalloproteinase (MMP1), and Aquaporin 1 (AQP1). TM-MSC were also positive for ABCG2, a marker for a side population of stem cells [22]. Although some of these markers may be expressed on other cell types, collectively the marker expression data indicate that TM-MSC have TM cell characteristics, suggesting that they may be progenitors of the meshwork tissue. Directed differentiation experiments that establish the developmental potential of TM-MSC are needed to confirm that they give rise to TM tissue.
To confirm that our TM-MSC isolation and culture method were not propagating a cell population of epithelial, endothelial, or neural crest origin, we analyzed TM-MSC for expression of vascular endothelial markers (CD31 [platelet-endothelial cell adhesion molecule; PECAM] and vascular-endothelial cadherin [VE-Cad]); epithelial markers (cytokeratin 3 [KRT3], cytokeratin 12 [KRT12], and α9-integrin [ITGA9]); as well as neural crest progenitor marker p75 (Supplementary Fig. S3). All markers were absent, excluding the possibility of Schlemm's Canal endothelial cells and corneal endothelial cells persisting in the TM-MSC cultures. TM-MSC expressed perivascular markers such as NG2, platelet-derived growth factor receptor-beta (PDGFRβ), and regulator of G protein signaling 5 (RGS5) (data not shown), the latter of which has been shown to inhibit endothelial cells in culture [23]. Taken together, these results demonstrate that TM-MSC do not express epithelial nor vascular endothelial markers. Our results also suggest that our TM-MSC culture conditions lack contamination from other cell types such as epithelial cells, keratocytes, and endothelial cells.
To ensure that the MSC population originated from the TM itself and not from the adjacent sclera and cornea, the sclera and cornea were dissected and digested separately with type I collagenase in DMEM containing 10% FBS. Scleral and corneal stromal cell cultures were then set up using the same culture conditions as TM-MSC. The resulting cells from both tissues appeared more elongated with a small and uniform nuclear-to-cytoplasmic ratio compared to TM-MSC (Supplementary Fig. S4A–C). They grew in swirls, a characteristic of cultured fibroblasts. Several TM markers (LDLR, CHI3L1, MYOC, and MMP1) were significantly downregulated in the corneal and sclera stromal cells compared to TM-MSC (Supplementary Fig. S4D). This suggests that TM-MSC were derived from the TM, not from the sclera or cornea.
Discussion
The study here demonstrates that the TM harbours a mesenchymal stem cell-like population. We established a procedure whereby a population of resident stem cells, TM-MSC, could be isolated and expanded as adherent cells in culture. These TM-MSC expressed typical MSC markers CD73, CD90, and CD105, but not hematopoietic markers. In addition, TM-MSC have the defining developmental potential of MSC—being able to differentiate into adipocytic, chondrocytic, and osteocytic lineages. Further evidence of their similarity to MSC was demonstrated by the overall similarity of gene expression profiles between TM-MSC and mesenchymal cells.
Primary cells derived from the TM have been reported previously [10, 24], but it is unclear if those cells possess stem cell-like properties. Those cells were described as having elongated cell bodies with centrally located, bulging nuclei, similar to our observations, but were not tested for the presence of MSC cell marker or differentiation potential as we have demonstrated (Fig. 1A). Other contaminating cells such as histiocytes, clump cells from the iris, remnants from corneal endothelium, and epithelial-like cells were also observed in those cultures [24]. As we largely based our dissection technique on Tripathi's and Grierson's methods, we also noticed some corneal endothelial cells growing in our initial TM-MSC culture as observed by their distinctive compact box-like morphology, but they did not survive subsequent passages (data not shown). To select specifically for plastic-adherent dividing cells and not terminally differentiated cells in our TM-MSC cultures, and to further assess clonogenic potential of our primary TM-MSC, the CFU-F assay was used [11]. We observed that 0.15% of seeded TM-MSC were able to form adherent colonies (Fig. 1D). Interestingly, this percentage is higher than that of age-corrected bone marrow-derived MSC (BM-MSC) [25]. In addition, as fibroblasts do not survive the CFU-F assay nor are they able to differentiate into mesenchymal lineages [26] and because our TM-MSC cultures were established from CFU-F colonies, the CFU-F assay served as a further purification step of selecting for proliferative MSC and not terminally differentiated fibroblasts. Therefore, the distinguishing difference between our culture method and those previously reported is that we have positively selected for progenitor cells within the TM, which we hypothesize are able to differentiate into the TM tissue proper.
That TM-MSC are progenitors of the mature TM is an attractive hypothesis. Our results have demonstrated conclusively that TM-MSC are multipotent: it is intriguing to suggest that due to their location within the corneoscleral limbus, TM-MSC may additionally be capable of differentiating into functional TM. Their similarity to MSC rather than other eye derivatives, such as the cornea, as demonstrated by gene expression profiling, indicates that this possibility may indeed be true. In addition, our results demonstrating that TM-MSC also express markers of mature TM such as ANK3, MYOC, and CHI3L1 are indicative that we have isolated a population of TM-MSC that will give rise to the TM.
That stem/progenitor cells exist in the TM and repopulate the TM to maintain physiological IOP is an attractive hypothesis. Age is one of the risk factors for primary open angle glaucoma (POAG), and TM decellularization has been observed in patients with glaucoma. In nonglaucomatous eyes, it was reported that TM cellularity decreased more than half (58%) and absolute cellularity decreased by 47% from newborn to 81 years of age [27]. Another study reported an average loss of 6,000 cells per year [28]. Indeed, sheath-derived plaques comprising thickened elastic fibers within the ciliary muscle due to the increased deposition of extracellular matrix (ECM) are present in greater numbers in POAG as compared to age-matched nonglaucomatous controls [29]. Not only is ECM deposited in greater amounts in glaucoma, but also matrix remodelling molecules and phagocytosis are decreased in glaucoma models [30, 31]. Thus, it is possible that the age-related loss of TM function linked to glaucoma could in part be due to a depletion of progenitor cells that maintain this tissue.
In summary, cells derived from the human TM have been widely studied, but there are currently no reports confirming the presence of stem/progenitor cells within the TM that may play a key role in regenerating diseased TM tissue and serve as a model for understanding the normal biological function of TM tissue. If we were to confirm that the TM-MSC are a progenitor cell type that functions to replace diseased TM tissue, our culture system of propagating TM progenitor cells would be a reproducible and robust alternative to currently available in vitro models of glaucoma, because we are able to utilize this TM-MSC culture system to investigate the pathogenesis of glaucoma from a stem cell perspective. Subsequent challenges would be to culture TM-MSC from glaucoma patients, and to investigate the hypothesis that glaucoma is a stem-cell disorder caused by TM-MSC depletion. More importantly, because glaucoma is a disorder afflicting nearly every ethnic group in the world, the ability to grow TM-MSC in high-throughput, individualized culture systems that can be easily manipulated, expanded, and differentiated ex vivo would radically transform current glaucoma therapeutic strategies.
Footnotes
Acknowledgments
The authors gratefully acknowledge helpful discussions with A. Bhinge, L. Lim, L.-F. Seet, and R. Wagey. The funding support for this work was from the Agency for Science, Technology and Research (A*STAR), and the National Medical Research Council of Singapore.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.