
Abstract
Gap junctional intercellular communication (GJIC) has been described in embryonic stem cells (ESCs) and various somatic cells. GJIC has been implicated in the regulation of cell proliferation, self-renewal, and differentiation. Recently, a new type of pluripotent stem cells was generated by direct reprogramming of somatic cells. Here, for the first time, we show that during reprogramming events GJIC is re-established upon reaching complete reprogramming. The opposite process of cell differentiation from the pluripotent state leads to the disruption of GJIC between pluripotent and differentiated cell subsets. However, GJIC is subsequently re-established de novo within each differentiated cell type in vitro, forming communication compartments within a histotype. Our results provide the important evidence that reestablisment of functional gap junctions to the level similar to human ESCs is an additional physiological characteristic of somatic cell reprogramming to the pluripotent state and differentiation to the specific cell type.
Introduction
P
Gap junctions are intercellular hydrophilic channels that allow direct diffusion of ions, signaling molecules, and metabolites between adjacent cells. These channels are formed of 2 hemichannels called connexons, and each of them is localized in the membrane of adjacent cells. Each connexon is composed of integral membrane connexin (Cx) proteins [8,9]. Expression of connexins Cx43 and Cx45 mRNA and dye coupling between undifferentiated ESCs were initially described in mouse cells [10]. It was shown that GJICs are maintained in hESCs propagated in vitro [11]. GJICs play a considerable role in the processes of embryonic development and in the induction of differentiation [12,13]. Cells in the preimplantation embryo are coupled with each other through gap junctions; however, as the first cell specialization occurs, ICM loses GJIC with trophectoderm cells [14]. Later in development, gap junctional coupling is only retained within each germ layer, and not across the boundaries between them, indicating that GJIC is maintained within the cell pools of the same lineage, although with a few exceptions [15,16]. However, little is known about the changes in gap junctions during differentiation of hESCs and iPSCs. Re-establishment of functional networks between differentiated cells of particular phenotype and forming the so-called communication compartment is the most likely basis for the beneficial outcome of successful differentiation to the particular phenotype in vitro because the ability of differentiated cells to form functional connections makes them more similar to their in vivo counterparts.
Given the critical importance of GJIC in cell type-specific, compartments we investigated GJIC formation during reprogramming to pluripotency and differentiation. Our data show that reprogramming accomplishment makes iPSCs indistinguishable from hESCs with respect to GJIC. We also demonstrate that during differentiation, GJIC is lost between pluripotent and differentiated cells, but re-established within differentiated cell types. These findings provide an additional proof that gap junction communication compartments mediate specific intercellular signaling between cells with similar cell fates.
Materials and Methods
Cell culture
The hESC lines hESM01 and hESM03 were previously described in [17]. Human umbilical vein endothelial cells (HUVECs) were derived as described [18]. The iPSC lines endo-iPS12 and endo-iPS10 were derived from HUVECs by lentiviral transfection with 4 transcription factors: KLF4, OCT4, SOX2, and C-MYC [19].
The hESC lines and endo-iPS12 were cultivated in the mTeSR1 medium (Stemcell Technologies) on Petri dishes coated with Matrigel (BD) according to the manufacturer's protocol. Cells were passaged every 5–7 days by exposure to 1 mg/mL dispase (Invitrogen) for 5–10 min at 37°C. To study gap junctions in iPSC clones during reprogramming, the endo-iPS10 cells at passage 6 and passage 26 [19], representing incompletely reprogrammed and fully reprogrammed cells correspondingly, were propagated in KnockOut Dulbecco's modified Eagle's medium (KO DMEM) supplemented with 20% Knockout SR, 2 mM L-glutamine, 1% MEM-nonessential amino acids solution (all from Invitrogen), 0.1 mM β-mercaptoethanol, and 4 ng/mL hrbFGF (Peprotech).
HUVECs were cultivated in DMEM/F12 with 15% fetal bovine serum (FBS), 5 ng/mL hrbFGF (Peprotech), 20 ng/mL hrVEGF (Peprotech), 1% nonessential amino acids, 2 mM L-glutamine, 50 units/mL penicillin, and 50 μg/mL streptomycin (all from Hyclone). All cell lines were maintained in 5% CO2 at 37°C.
Differentiation of hESCs and iPSCs
To obtain partially differentiated colonies, pluripotent cells were cultivated for 8 days in feeder-free conditions as described above. The culture medium was then replaced with a medium containing 1 part mTESR medium and 1 part KO DMEM supplemented with 15% FBS. For later stages of differentiation, we continued cultivation for 3 additional weeks in the medium containing DMEM/F12, 15% FBS, nonessential amino acids, and 2 ng/mL bFGF.
Reverse transcription–polymerase chain reaction
Total RNA was isolated from cells using RNeasy Mini kit (Qiagen). Reverse transcription–polymerase chain reaction (RT-PCR) was performed using RNA (1–2 μg), random hexamers (Promega), and M-MLV reverse transcriptase (Promega) according to the manufacturer's recommendations. All RNA samples were adjusted to yield equal amplification of glyceraldehyde-3-phosphate dehydrogenase as an internal standard. Negative control RT-PCRs were carried out under the same conditions, without addition of M-MLV RT. Primers are listed in Supplementary Table S1 (Supplementary Data are available online at
Lucifer yellow injection
GJIC was determined by the dye coupling method as described previously [20]. Experiments were performed in Ca2+ Mg2+ PBS (Paneco). Fluorescent dye Lucifer yellow (LY) CH (dilithium salt, 4% w/v solution in 1 M LiCl; Sigma, L 0259, MW 457 Da) was injected using negative current (20 nA, 10 s) in a single cell of a cellular monolayer. Microinjection of rhodamine dextran (Invitrogene) into cells was used as negative control. The molecular mass of rhodamine dextran (10 kDa) is too big for transfer between neighboring cells through GJIC. The dye spread to the adjacent cells was monitored under the Axiolab fluorescence microscope (Zeiss) equipped with phase-contrast optics, a 40× water immersion lens, and CCTV camera (Panasonic). The number of LY-stained recipient cells by each donor cell was counted 2 min after dye injection. Twenty to 30 donor cells in a continuous field were measured in each sample. The data represent means±standard deviation.
Immunocytochemistry
For immunohistochemical studies, cells were fixed in 4% paraformaldehyde for 20 min at room temperature. Fixed cells were incubated for 1 h in blocking buffer. [phosphate-buffered saline (PBS), 0.2% Triton ×100, 0.1% Tween-20, 2% goat or donkey serum, and 2% FBS]. Primary antibodies were diluted in PBS, 0.1% Tween-20, and applied for 1–3 h at room temperature or overnight at 4°C. Antibodies used were as follows: Oct3/4 (Abcam; cat# ab19857), CD31 (Dako; cat #MO823), Gata 6 (Abcam; cat #Ab22600), MOC-31 epithelial cell adhesion molecule (EPCAM) (Dako; cat #M3525), and Nanog (Abcam cat # ab77095). Slides were washed 3 times with PBS and 0.1% Tween-20, and incubated with secondary antibodies: Alexa Fluor 546, Alexa Fluor 488-conjugated goat anti-mouse, or goat anti-rabbit or donkey anti-goat (all from Invitrogen) at a dilution 1:1000. Nuclei were stained with DAPI (Sigma). Slides were mounted in the Vectashield mounting medium (Vector Laboratories) and examined under the fluorescent microscope Zeiss AxioImager A1 (Zeiss).
Results
GJIC re-establishment during reprogramming to pluripotency
We determined the ability of pluripotent cells to communicate through gap junctions by measuring the number of cells that LY injected into 1 cell was transferred within the colony. LY was injected into colonies of feeder-free endo-iPSC and hESC lines. Colonies of fully reprogrammed endo-iPSC-12 cell line had a high level of GJIC, the number of stained cells was 65±2 cells/donor cell (n=23) (Fig. 1A, B). This value was comparable to the number of stained cells in hESCs, where the dye transfer occurred in 72±3 cells/donor cell (n=30) (Fig. 1C, D). HUVECs parental to endo-iPSCs were used to determine GJIC in somatic cells [19]. Dye transfer between somatic cells was slower than that in pluripotent, only 18±1 (n=20) cells/donor cell (Fig. 2). Acquisition of pluripotency may continue for several passages [21 –24]. To evaluate possible changes in GJIC during late stages of reprogramming, we used incompletely reprogrammed endo-iPSC10 cell line at passage 6 and completely reprogrammed cells of the same line at passage 26. Incompletely reprogrammed iPSCs were characterized by residual expression of endothelial-specific genes and reduced expression of pluripotency-related genes (Supplementary Fig. S1A). In incompletely reprogrammed cells, residual transgene expression was detected at passage 6 (Supplementary Fig. S1B). Nanog expression was observed in both incompletely and faithfully reprogrammed iPSCs (Fig. 2). Expression of TRA-1-60 antigen was utilized to determine faithfully reprogrammed cells [21,25,26]. Colonies of endo-iPSCs at different passages were injected with LY; after counting the number of stained cell, colonies were fixed for TRA-1-60 immunofluorescence detection. Surprisingly, the number of communicating incompletely reprogrammed cells was much lower than those in the parental HUVECs and did not exceed 5±5 cells/donor cell (Fig. 2). The lack or poor dye diffusion was well correlated with low TRA-1-60 antigen expression in these cells. At the later passages cells were strongly positive for TRA-1-60, demonstrating a high level of GJIC (Fig. 2).
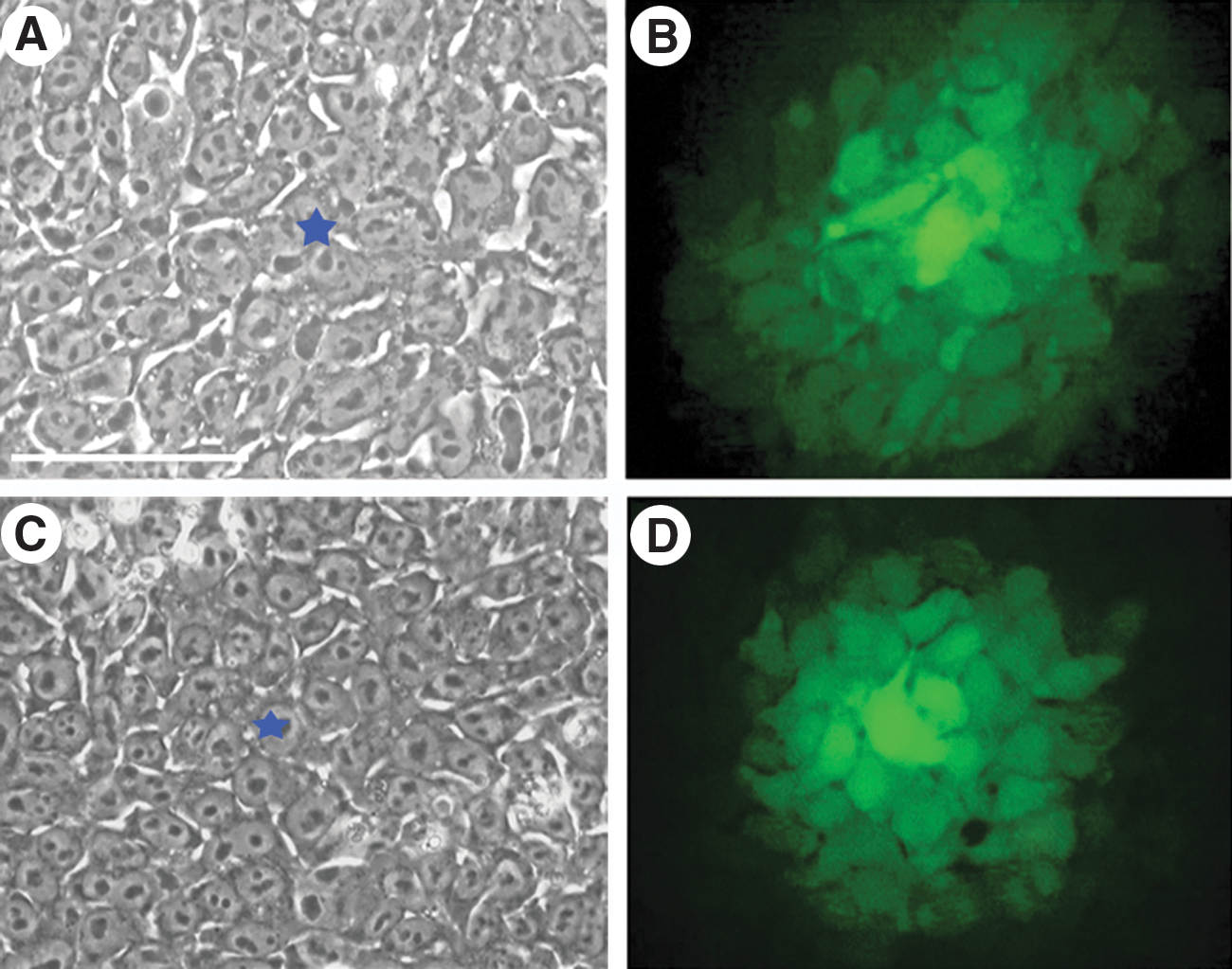
Distribution of Lucifer yellow (LY) dye in undifferentiated endo-induced pluripotent stem cell (iPSC) and human embryonic stem cell (hESC) colonies.

Loss and re-establishment of gap junctional intercellular communication (GJIC) during reprogramming. Cells subjected to LY injection are marked with the blue stars. Scale bars are presented in phase-contrast images. Scale bars represent 100 μm. TRA-1-60 staining is red, Nanog staining is green, 4′,6-diamidino-2-phenylindole (DAPI) staining is blue.
GJIC of each particular cell type may be characterized by connexins composition. We compared expression of connexins in HUVEC, endo-iPS-10, 12, and hESCs using transcriptome analysis (Supplementary Method and Fig. S2). We found that only connexin Cx37 and Cx43 expression varied significantly in the examined cell types. RT-PCR data confirmed microarray expression analysis (Fig. 3). In incompletely reprogrammed iPSCs, Cx37 and Cx43 were expressed at the level similar to HUVEC. In faithfully reprogrammed iPSCs, cells lacked characteristic for parental HUVEC connexin Cx37 expression, whereas Cx43 expression was increased 3–5-fold, reflecting both characteristic for pluripotent cells connexins composition and their number.

Expression of connexins Cx37 and Cx43 in human umbilical vein endothelial cell, hESCs, incompletely and fully reprogrammed iPSCs, evaluated by reverse transcription–polymerase chain reaction.
Thus, during the reprogramming process, the genetic and epigenetic conversion of endothelial cells to the pluripotent state is also accompanied by significant physiological changes, GJIC between cells is dramatically reduced, and cells became independent of their neighboring cells. Upon the accomplishment of the reprogramming process, GJIC between pluripotent cells is re-established de novo involving specific to pluripotent cells connexins.
Pluripotent stem cells are not connected with their differentiated derivatives by GJICs during early steps of spontaneous differentiation
To analyze changes in GJIC during the loss of pluripotency, the culture medium was replaced to initiate spontaneous differentiation of hESCs or iPSCs. Partially differentiated colonies in this case acquire typical bagel-like morphology (Fig. 4A). At the periphery of the bagel-like colony, significantly compacted ES-like cells formed a multilayer (Fig. 4B, yellow arrows), whereas the cells in the central part of the colony were larger and had different morphology (Fig. 4B, blue arrows) [27]. Immunostaining of these structures with Oct-4 antibodies revealed that only peripheral cells remained Oct-4 positive, indicating that the cells located at the center of the colony entered differentiation and lacked pluripotency, whereas the cells surrounding them were still pluripotent (Fig. 4B). The boundary between differentiated and undifferentiated cells was clearly visible within each colony; thus, dye injection could be preferentially performed in the differentiated or undifferentiated parts of the colonies. The first LY injection was performed into a pluripotent part of the colony (Fig. 4C, yellow stars); the second injection was performed into a differentiated part of the colony (Fig. 4C, blue stars). After the first injection, numerous pluripotent Oct-4-positive cells were well stained (>40 cells/donor cell), whereas after a second injection, we did not observe significant dye transfer out of the differentiated donor cell. However, it is worth to mention that the permeability of gap junctions for the dye in the differentiated part of the colony varied significantly, from a single stained cell up to 10 neighboring cells. Notably, the dye injected in any part of the colony was distributed only among the pluripotent or differentiated cells, but never crossed the boundary between these areas, indicating that pluripotent and differentiated cells represent 2 independent communication compartments (Fig. 4C).

Loss of GJIC between pluripotent and differentiating cells during early stages of spontaneous differentiation.
In differentiated cultures' GJIC is re-established between the cells of a similar origin
The partially differentiated colony represents an early step of a spontaneous differentiation process in in vitro culture of pluripotent stem cells. At this stage, GJIC within differentiated areas is weak or even absent. Morphological changes occur with further differentiation, and after 3–4 weeks of differentiation, cultures show a mixture of morphologically different cells, representing different histotypes. We wondered whether GJIC was re-established in de novo formed histological compartments. To address this question, LY was injected into cells located in the vicinity of an apparent morphological boundary between the cells presumably belonging to different histotypes (Fig. 5). Cells were then fixed, and immunocytochemistry with various antibodies was performed to identify the groups of cells coupled by GJIC. In few cases, we succeeded in identifying particular cellular lineages (Fig. 5). Notably, clusters of cells coupled by gap junctions had markers distinct from those of neighboring cells, where the fluorescent dye did not spread. As shown in Fig. 5A there are 2 groups of cells where the dye spread between cells in each group. Staining for CD31 confirmed that the cells of 1 group belonged to the same histotype (presumably, endothelial cells). Although CD31-negative cells were coupled by gap junctions within the cluster, they belonged to a communication compartment that differed from endothelial cells.

De novo re-establishment of gap junction communication compartments in differentiated pluripotent stem cells in vitro. Left column is a phase-contrast image of differentiated cultures of hESCs or iPSCs. Cells injected with LY are marked with stars. In the case of 2 injections of LY
A number of different cell types could be observed during differentiation. Two types of cells with presumably epithelial and fibroblast-like morphologies are shown in Fig. 5B. LY injections were performed in both groups of cells. First, LY was injected into the fibroblast-like cell. Within 2 min, LY spread into 2–3 neighboring fibroblast-like cells, but not to the cells of epithelial morphology. Similarly, the LY injection into the epithelial-like donor cell revealed GJIC between several epithelial-like cells but did not result in the fluorescent dye transfer into fibroblast-like cells. Immunostaining analysis for EPCAM confirmed that coupled cells belonged to the same tissue type (presumably epithelium). EPCAM-negative cells (fibroblast-like cells) were not coupled by GJIC with EPCAM-positive cells. Cells positive for GATA-6 also re-established GJIC within the cluster and did not communicate by GJIC with GATA-6 negative neighboring cells (Fig. 5C).
Discussion
Evidence for electric and dye coupling of embryonic cells has been demonstrated in a variety of species. In both mouse and human embryos, organization of gap junctions has been shown to begin at the 2–8-cell stage [12 –14]. Similar to cells of the ICM, undifferentiated ESCs express connexins Cx43 and Cx45 and display functional GJIC [11,28]. Most somatic cell types, in particular, HUVEC, express connexins and have functional GJIC [29]. iPSCs appear almost identical to hESCs and share the same pivotal features of self-renewal and pluripotency. However, it is still little known about intercellular communications during reprogramming process. In our study, we showed that during reprogramming events, GJIC is dramatically reduced between the cells undergoing reprogramming. This finding indirectly supports the hypothesis that mesenchymal-to-epithelial transition has to occur during reprogramming [30]. Weak or lack of GJIC is characteristic for epithelial stem cells, such as keratinocyte stem cells, breast epithelial, and neural-glial stem cells [31]. However, GJIC is also downregulated in cancer cells [32]. The stem cell theory of cancer is based on the postulate that, in contrast to differentiated cells, adult stem cells lacked GJIC and became the targets of carcinogenesis events [33]. iPSCs are now considered as a very promising source of cells for therapeutic purposes. Safety issues of cell reprogramming are very important. If GJIC is not completely reestablished in iPSCs cells and the cells are at least partially independent of neighboring cells, it possibly means that these cells are at the stage similar to oncogenic transformation. Thus, complete re-establishing of GJIC in iPSCs is an important physiological marker of the reprogramming accomplishment, and a prospective physiological indicator for evaluating the feasibility of a particular iPSC line for therapeutic purposes.
Expression levels of connexins Cx43 and Cx45 are considered to be a marker of undifferentiated hESCs state [11]. Downregulation of Cx43 and Cx45 and GJIC during hESC differentiation into squamous epithelial cells was recently described in [34]. In our study we demonstrated that completely reprogrammed iPSCs have strong GJIC and express high levels of connexin 43. Surprisingly, we did not observe any significant level of connexin 37, neither in hESCs nor in iPSCs, which is not consistent with the data previously published in [35]. Moreover, connexin 37 was strongly expressed in parental HUVEC, but underwent silencing upon complete reprogramming. This observation could be explained by different culture conditions. In our study pluripotent stem cells were cultured in an defined medium without feeder cells, thus presumably indicating that Cx37 expression far most reflects culture conditions rather than pluripotent stem cell marker. In contrast to the connexin Cx37 expression, the level of connexin Cx43 transcription augments dramatically during reprogramming, indicating the importance of connexin Cx43 for the accomplishment of reprogramming or maintenance of a pluripotent state. Therefore, connexin 43 expression could be suggested as an additional molecular marker to validate complete reprogramming.
At the earliest stages of spontaneous differentiation of hESCs and iPSCs, the GJIC between the cells that undergo differentiation and the cells remaining pluripotent is completely disrupted in both directions. We did not observe dye diffusion from the pluripotent cells to the differentiating cells, or from the differentiating to pluripotent cells. Similar situation is observed in vivo when cells of ICM remain coupled but the spread of fluorescent dye to trophoblast cells dramatically decreases [14]. GJIC within the pool of differentiating cells may vary significantly, from 10 neighboring cells to complete absence of dye distribution. At the same time, a high level of GJIC always remains between the pluripotent cells. A high level of cooperation between pluripotent cells ensures high stability in maintaining the biological status of these cells, because any metabolic change in a single cell (eg, in case of various deviations in the genome function) will be minimized by strong fluxes of compounds entering from neighboring cells. Therefore, the loss of GJIC at the beginning of hESCs and iPSCs differentiation could be a key event toward the progression of differentiation in a pluripotent cell culture.
Further studies are required to clarify the reasons of GJIC variability at the early steps of differentiation. Possibly, the presence or absence of GJIC in this case is not crucial for further progression of ongoing differentiation. However, it is more likely that the loss of GJIC between cells is physiologically functional and important for the individual genetic program realization and subsequent differentiation events leading to the particular histotype formation. We demonstrated that at the later stages of pluripotent stem cell differentiation, cells gain GJIC and these connections are restricted by the cells of the same histotype. Therefore, during differentiation and cell commitment in vitro, re-establishment of GJIC occurs within a particular communication compartment between cells of a similar fate. We cannot exclude that cells of distinct histotypes could establish GJIC during further differentiation in vitro, like it occurs during more advanced steps of embryonic development and in adult organism; however, we did not observe it in our cell model and time points. Loss of GJIC at the beginning of pluripotent stem cell differentiation and then re-establishment of communications between the cells of the same histotype are processes similar to those during early embryonic development, when the embryo cells form communication compartments in which GJIC remains only between the cells of the same germ layer [15]. We believe that our study shows that such a process occurs in human pluripotent cell cultures.
Conclusion
We demonstrate that reprogramming to pluripotency disrupts existing in somatic cells GJIC; however, reprogramming accomplishment restores the GJIC level and connexin expression pattern to that found in hESCs. The results enable the conclusion that GJIC might be an additional important characteristic of faithful cell reprogramming.
Footnotes
Acknowledgments
This work was supported by Russian Foundation for Basic Research, Moscow Anticancer Program and Russian Ministry of Education and Science.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.