
Abstract
Human adult stem cells (hASCs) have become an attractive source for autologous cell transplantation, tissue engineering, developmental biology, and the generation of human-based alternative in vitro models. Among the 3 germ cell layers, the mesoderm is the origin of today's most widely used and characterized hASC populations. A variety of isolated nonhematopoietic mesoderm-derived stem cell populations exist, and all of them show important differences in terms of function, efficacy, and differentiation potential both in vivo and in vitro. To better understand whether the intrinsic properties of these cells contribute to the overall differentiation potential of hASCs, we compared the global gene expression profiles of 4 mesoderm-derived stem cell populations: human adipose tissue-derived stromal cells, human bone marrow-derived stromal cells (hBMSCs), human (fore)skin-derived precursor cells (hSKPs), and human Wharton's jelly-derived mesenchymal stem cells (hWJs). Significant differences in gene expression profiles were detected between distinct stem cell types. hSKPs predominantly expressed genes involved in neurogenesis, skin, and bone development, whereas hWJs and, to some extent, hBMSCs showed an increased expression of genes involved in cardiovascular and liver development. Interestingly, the observed differential gene expression of distinct hASCs could be linked to existing differentiation data in which hASCs were differentiated toward specific cell types. As such, our data suggest that the intrinsic gene expression of the undifferentiated stem cells has an important impact on their overall differentiation potential as well as their application in stem cell-based research. Yet, the factors that define these intrinsic properties remain to be determined.
Introduction
H
Materials and Methods
Isolation and cultivation of hADSCs
Plastic surgical waste material (ie, abdominal fat) is obtained in cooperation with the Department Plastic Surgery of the UZ-Brussels (Belgium) and the ATLAS Kliniek (Belgium) upon informed consent of the patient and approval by the Ethics Committee of the UZ-Brussels. The median age of the donors is 39 years (n=7; F(3)/M(4); range 32–49). hADSCs are isolated and subcultivated as previously described [7]. Briefly, ∼125 g of processed adipose tissue is incubated for 90 min at 37°C in a dissociation medium (1:1) consisting of 1% (v/v) bovine serum albumin (BSA; Sigma-Aldrich) and 1 mg/mL collagenase A (Roche Applied Science) in phosphate-buffered saline (PBS). After 2 filtration steps, the filtrate is carefully brought on top of 15 mL of Histopaque®-1077 (Sigma-Aldrich). Upon centrifugation for 20 min at 1,000 g (4°C), the top layer is removed, and the hADSCs are collected in 50 mL PBS/BSA (1%). This procedure is carried out separately on 2 pieces of adipose tissue. Typically, 5–20×107 viable cells are obtained per 250 g of processed adipose tissue. The isolated hADSCs are then subcultured as a monolayer in an hADSC growth medium for 2 weeks, consisting of the Dulbecco's modified Eagle's medium (DMEM; Lonza) supplemented with 10% (v/v) fetal bovine serum (FBS; Perbio Hyclone), 50 μg/mL streptomycin sulfate (Sigma-Aldrich), 7.33 IU/mL benzyl penicillin (Continental Pharma), and 2.5 μg/mL fungizone (Life Technologies). Cell cultures are incubated at 37°C in a 5% (v/v) CO2-humidified atmosphere and passaged at subconfluency using TrypLE® express (Life Technologies). The growth medium is changed every 3 days. hADSCs between passages 3 and 5 are used for transcriptomic analyses.
Isolation and cultivation of hBMSCs
Bone marrow is either obtained by sternal puncture aspiration in healthy volunteers or by needle aspiration from the iliac crest of bone marrow transplant donors as previously described [26]. The median age of the donors is 26 years (n=10; F(4)/M(6); range 3–57). Informed consent is obtained from all donors or their parents. The Ethics Committee of the Jules Bordet Institute approved the use of the tissue material for this study. Briefly, mononuclear cells (MNCs) are isolated from bone marrow aspirates by density-gradient centrifugation (Linfosep; Biomedics) and washed in the Hank's buffered salt solution (HBSS; Lonza). MNCs are seeded at a cell density of 2×104 cells/cm2 in a low-glucose DMEM (DMEM-LG; Lonza) supplemented with 15% (v/v) heat-inactivated FBS, 2 mM L-glutamine, and 0.5% (v/v) antibiotic/antimycotic solution (all from Life Technologies). Cells are incubated at 37°C in a 5% (v/v) CO2-enriched humidified atmosphere, cultured up to 90% confluency, trypsinized (Tryple Select solution; Lonza), centrifuged, and subcultured at a lower density (5×103 cells/cm2) for all subsequent passages for 2 weeks. hBMSCs between passages 2 and 4 are used for transcriptomic analyses.
Isolation and cultivation of hSKPs
hSKPs are isolated and subcultivated as previously described [6]. Informed consent was obtained from both parents. The median age of the donors is 3 years (n=11; M(11); range 1–3). Briefly, freshly collected human foreskin samples are incubated with 25 mL of 0.2 mg/mL Liberase DH solution (Roche Applied Science) and incubated for 20 h at 4°C. Next, the epidermis is removed, and the tissue is incubated at 37°C for another 10–20 min depending on the sample size. After processing the samples, typically 5–15×106 viable cells are obtained per 5–8-cm2 foreskin. For cultivation, a cell density of 2×104 cells/cm2 is applied. The growth medium for hSKPs consists of a DMEM+GLUTAMAX/F12 Nutrient Mixture (3:1; Life Technologies) supplemented with 7.33 IU/mL benzyl penicillin (Continental Pharma), 50 μg/mL streptomycin sulfate (Sigma-Aldrich), 2.5 μg/mL fungizone, 2% (v/v) B27 Supplement (Life Technologies), 40 ng/mL basic fibroblast growth factor-2, and 20 ng/mL epidermal growth factor (both from Promega). Cell cultures are incubated at 37°C in a 5% (v/v) CO2-humidified atmosphere for 2 weeks. The growth medium is replenished every 2–3 days. hSKPs spheres are passaged every 2 weeks using 0.2 mg/mL Liberase DH solution (Roche Applied Science). hSKPs between passages 1 and 4 are used for transcriptomic analyses.
Isolation and cultivation of hWJs
After receiving informed consent from the mothers involved, umbilical cords (n=9; F(5)/M(4)) are collected and processed according to the protocol of De Bruyn et al. [27]. Only full-term deliveries are included in this study. The MSCs are isolated from WJ without enzymatic digestion or dissection, as the procedure is based on the migratory and plastic adhesive properties of MSCs. Briefly, umbilical cord segments of 5–10 cm are cut longitudinally and plated for 5 days in DMEM-LG (Lonza) supplemented with 15% (v/v) heat-inactivated FBS, 2 mM L-glutamine, and 0.5% (v/v) antibiotic/antimycotic solution (all from Life Technologies). After removing the cord segments, the culture is pursued until subconfluence. Cell cultures are incubated at 37°C in a 5% (v/v) CO2-humidified atmosphere. After 48 h, nonadherent cells are removed by washing, and the medium is changed twice a week. When subconfluency (80%–90%) is achieved, adherent cells are harvested after detachment by 10-min incubation with the TrypLE® Select solution (Lonza) and expanded for all subsequent passages by re-plating at a lower density (1×103 cells/cm2) for 2 weeks. hWJs between passages 2 and 5 are used for transcriptomic analyses.
Isolation of RNA and reverse transcriptase-polymerase chain reaction
For quantitative real-time polymerase chain reaction (qPCR) analysis, total RNA is extracted from all samples using the GenElute Mammalian Total RNA Purification Miniprep Kit (Sigma-Aldrich), according to the manufacturer's instructions. The isolated RNA is quantified at 260 nm using a Nanodrop spectrophotometer (Thermo Scientific). Total RNA is reverse-transcribed into cDNA using an iScript™ cDNA Synthesis Kit (BioRad), followed by cDNA purification with the GenElute PCR clean up kit (Sigma-Aldrich). For microarray analysis, the RNA is extracted using TRIzol/chloroform and purified with RNeasy minicolumns, as recommended by the manufacturer's instructions (Qiagen).
Quantitative real-time PCR
cDNA products are used for quantitative amplification of the target genes. The gene expression assays used in this study are listed in Supplementary Table S1 (Supplementary Data are available online at
qPCR data analysis
The qPCR efficiency is estimated by iQ5 Optical System Software (Version 2), and the data are only used when the calculated PCR efficiency ranges 0.85–1.15. Moreover, for selecting reliable reference genes to normalize the qPCR data, we first evaluated the expression stability of 6 candidate reference genes: glyceraldehyde 3-phosphate dehydrogenase (GAPDH), beta-2-microglobulin (B2M), hydroxy-methylbilane synthase (HMBS), 18S, beta-actin (ACTB), and ubiquitin C (UBC). According to geNorm®, the optimal number of reference targets to be used in this experiment is 5 (V<0.15). As such, B2M, UBC, 18S, HMBS, and GAPDH were selected as the most stable reference genes in all 4 stem cell populations using qbasePLUS® software (geNorm®; Biogazelle). Thereafter, to compare the relative mRNA expression levels of the target genes, results are expressed as the fold changes normalized against the geometric means of all 5 reference gene mRNAs using qbasePLUS® software (Biogazelle). Statistical analyses are performed using a 1-way unpaired ANOVA with correction for multiple testing. Gene expressions with a fold change of at least 2-fold and a corrected P value≤0.05 are considered to be significantly different.
Microarray data analysis
All reagents and instrumentation pertaining to oligonucleotide microarrays are procured from Affymetrix (Affymetrix,
Results
Mesoderm-derived stem cells isolated from various tissues differentially express genes involved in bone and cardiovascular development
To evaluate the variation between the individual samples of each stem cell type, a PCA plot was first assessed (Supplementary Fig. S1). This analysis shows only minor differences between the samples of each investigated stem cell population, as they consistently group together. Yet, considerable differences between distinct stem cell populations can be observed (Supplementary Fig. S1). Therefore, expression patterns of genes involved in the ontogeny of mesoderm-derived tissues were used to evaluate the intrinsic mesodermal commitment of the 4 mesoderm-derived stem cell populations at a molecular level. Hierarchical clustering was carried out to give an overview of the differentially expressed transcripts known to be involved in mesodermal lineage development (Fig. 1). Genome-wide microarray analyses, using the Ward's hierarchical clustering method, show a close relation between different donors of the same cell type as they group together. Furthermore, when comparing hADSCs, hBMSCs, hSKPs, and hWJs, a closer proximity is observed between hADSCs and hSKPs than between other combinations (Fig. 1). These data are supported by qPCR analyses where no significant differences in gene expression could be observed between hADSCs and hSKPs for (i) MSC markers, including stem cell growth factor receptor (KIT), stromal antigen 1 (STAG1), thymocyte differentiation antigen 1 (THY1) (Figs. 1 and 2), and vimentin (VIM) (Figs. 3 and 4), and (ii) for genes related to mesodermal lineage specification, including bone morphogenetic protein 2 (BMP2), gap junction protein alpha 1 (GJA1), intercellular adhesion molecule 1 (ICAM1), runt-related transcription factor 2 (RUNX2), and vascular cell adhesion molecule 1 (VCAM1) (Figs. 1 and 2). Interestingly, hSKPs show a significantly increased gene expression associated with bone and cartilage development. These include genes encoding for the bone morphogenetic proteins BMP2 and BMP4 (Figs. 1 and 2), paired box 3 (PAX3), and twist homolog 1 (TWIST1) (Figs. 3 and 4). In addition, compared to the other mesoderm-derived stem cells, hSKPs maintain a similar expression level of other genes related to bone and cartilage development, including RUNX2 (Figs. 1 and 2) and the muscle segment homeobox (MSX) gene family members MSX1 and MSX2 (Figs. 3 and 4). Only hBMSCs show a significant higher expression of RUNX2 compared to hSKPs (Fig. 2). Furthermore, Gene Ontology (GO) analyses of hSKPs using IPA reveal a significant enrichment of functional gene classes associated with bone development (Table 1). Indeed, ∼13% of the genes involved in development of bone and formation of bone are significantly increased in hSKPs compared to the other mesoderm-derived stem cell populations (Table 1). Another interesting observation is the highly increased expression in hWJs of the genes coding for GATA-binding protein 6 (GATA6) (Figs. 5 and 6), the transcription factor heart and neural crest derivatives expressed 1 (HAND1), ICAM1, and VCAM1 (Figs. 1 and 2). GO analyses of hWJs show indeed significant enrichment of functional gene classes involved in cardiovascular system development, including angiogenesis, cardiogenesis, development of blood vessel, development of endothelial tissue, endothelial cell development, and vasculogenesis compared to the other mesoderm-derived stem cell types (Table 2). In addition, other genes involved in cardiovascular development, including endoglin (ENG), GJA1, VCAM1 (Figs. 1 and 2), and GATA6 (Figs. 5 and 6), are significantly increased in hBMSCs. GO analyses of hBMSCs show significant enrichment of functional gene classes involved in cardiovascular ontogeny, including angiogenesis, development of blood vessel, and vasculogenesis, compared to hADSCs and hWJs, but not toward hSKPs (Table 3).
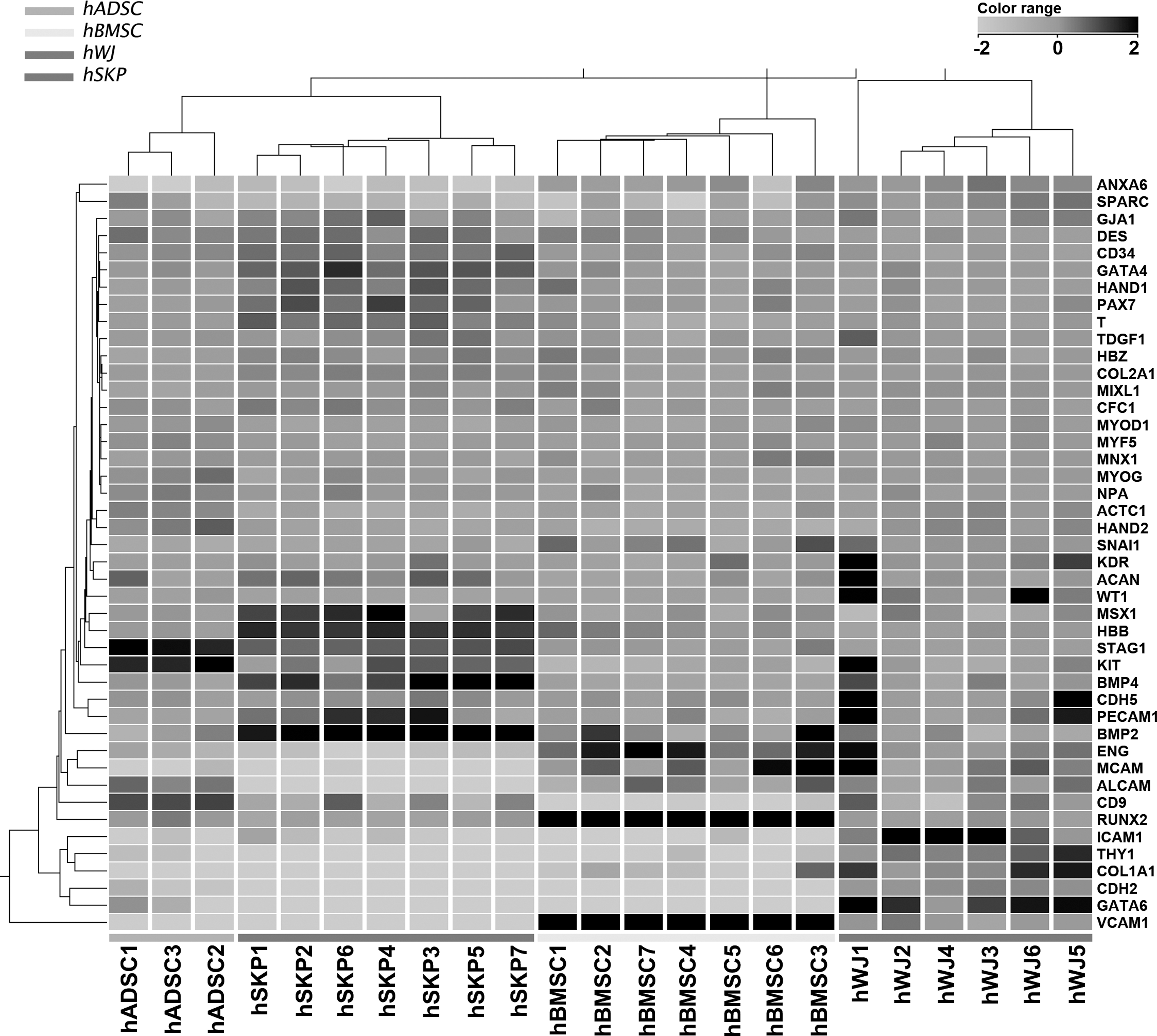
Microarray analysis shows differential expression of genes involved in bone and cardiovascular development in various mesoderm-derived stem cells. Heat map showing the relative expression levels of 44 genes involved in mesodermal lineage development and commitment in 3 independent hADSC, 7 independent hSKP, 7 independent hBMSC, and 6 independent hWJ cell isolates. Relative expression levels are color-coded as per the color key. Ward's hierarchical clustering shows a closer proximity between (i) hADSCs and hSKPs than (ii) hBMSCs and (iii) hWJs. hADSCs, human adipose tissue-derived stromal cells; hBMSCs, human bone marrow-derived stromal cells; hSKPs, human (fore)skin-derived precursor cells; hWJs, human Wharton's jelly-derived mesenchymal stem cells.
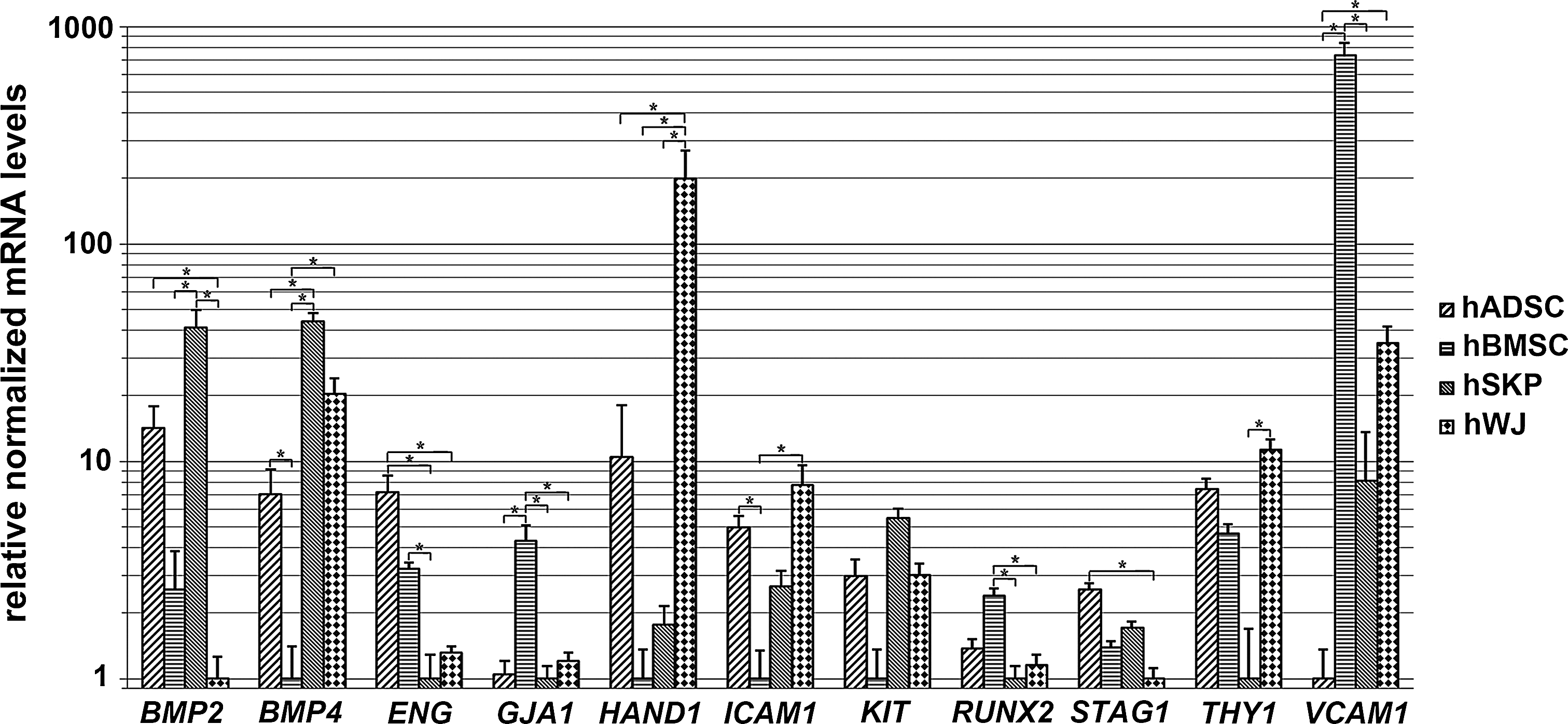
Differential expression of genes involved in bone and cardiovascular development is confirmed by quantitative real-time PCR analyses. Fold changes of genes involved in mesodermal development and commitment are determined for all 4 mesoderm-derived stem cell types (nhADSCs=4; nhBMSCs=3; nhSKPs=4; nhWJ=3). *Significantly increased or decreased mRNA expression between mutually compared stem cell types (fold change >2; P value<0.05). BMP, bone morphogenetic protein; ENG, endoglin; GJA1, gap-junction protein alpha 1; HAND1, heart and neural crest derivatives expressed 1; ICAM1, intercellular adhesion molecule 1; PCR, polymerase chain reaction; RUNX2, runt-related transcription factor 2; KIT, stem cell growth factor receptor; STAG1, stromal antigen 1; THY1, thymocyte differentiation antigen 1; VCAM1, vascular cell adhesion molecule 1.

Microarray analysis shows increased expression of genes implicated in neurogenesis and skin development in hSKP cells. Heat map showing the relative expression levels of 48 genes involved in ectodermal lineage development and commitment in 3 independent hADSC, 7 independent hSKP, 7 independent hBMSC, and 6 independent hWJ cell isolates. Relative expression levels are color-coded as per the color key. Ward's hierarchical clustering shows a closer proximity between (i) hBMSCs and hWJs than (ii) hADSCs and (iii) hSKPs, whereas hSKP cells show the highest expression of neural (crest)-related genes and hADSCs the lowest.
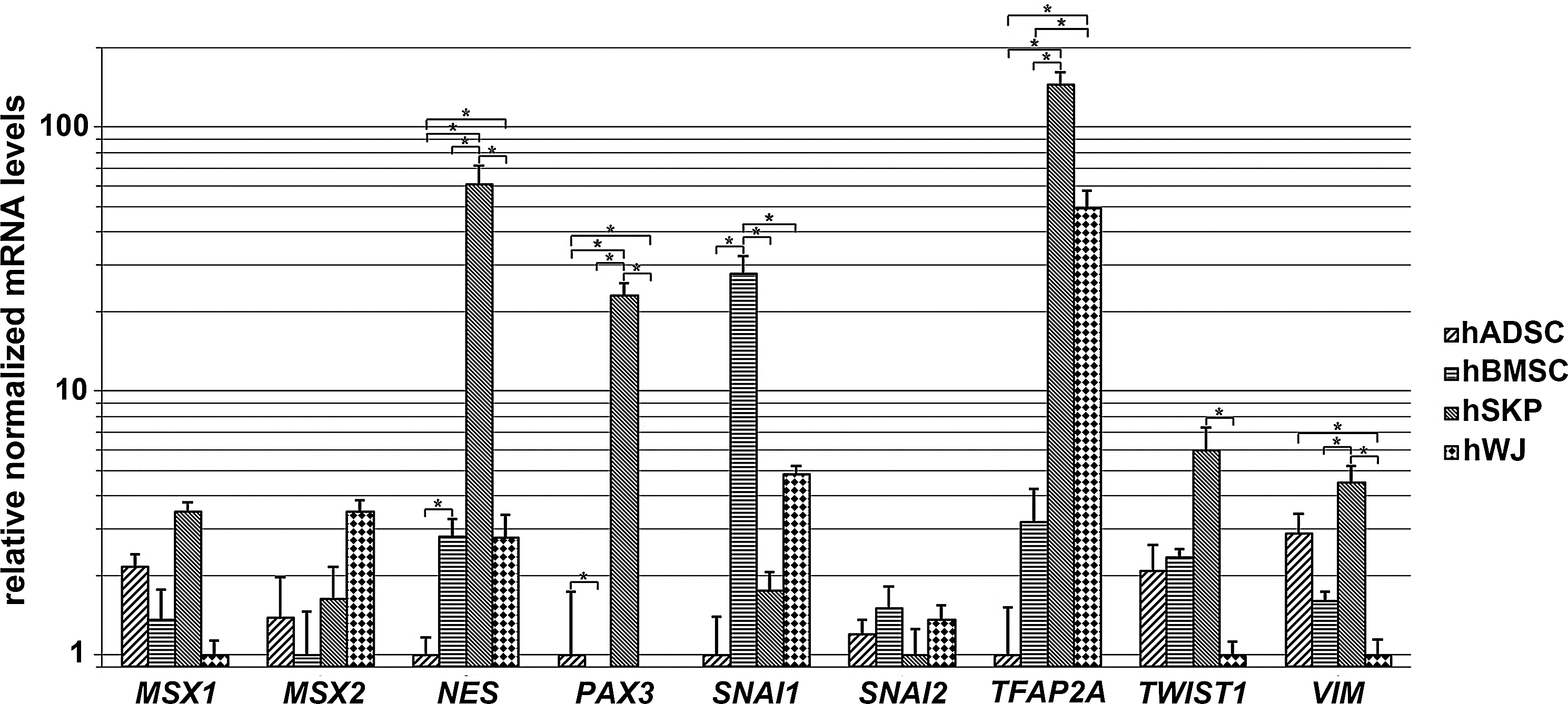
Differential expression of genes involved in ectodermal development is confirmed by quantitative real-time PCR analyses. Fold changes of genes involved in ectodermal development and commitment are determined for all 4 mesoderm-derived stem cell types (nhADSCs=4; nhBMSCs=3; nhSKPs=4; nhWJs=3). *Significantly increased or decreased mRNA expression between mutually compared stem cell types (fold change >2; P value<0.05). TFAP2A, activating enhancer-binding protein 2-alpha; MSX, muscle segment homeobox homolog; NES, nestin; PAX3, paired box 3; SNAI1, snail homolog 1; SNAI2, slug; TWIST1, Twist homolog 1; VIM, vimentin.

Microarray analysis shows increased expression of genes involved in endodermal development in Wharton's jelly-derived stromal cells. Heat map showing the relative expression levels of 48 genes involved in endodermal lineage development and commitment in 3 independent hADSC, 7 independent hSKP, 7 independent hBMSC, and 6 independent hWJ cell isolates. Relative expression levels are color-coded as per color key. Ward's hierarchical clustering shows a closer proximity between (i) hADSCs and hWJs than (ii) hBMSCs and hSKPs.
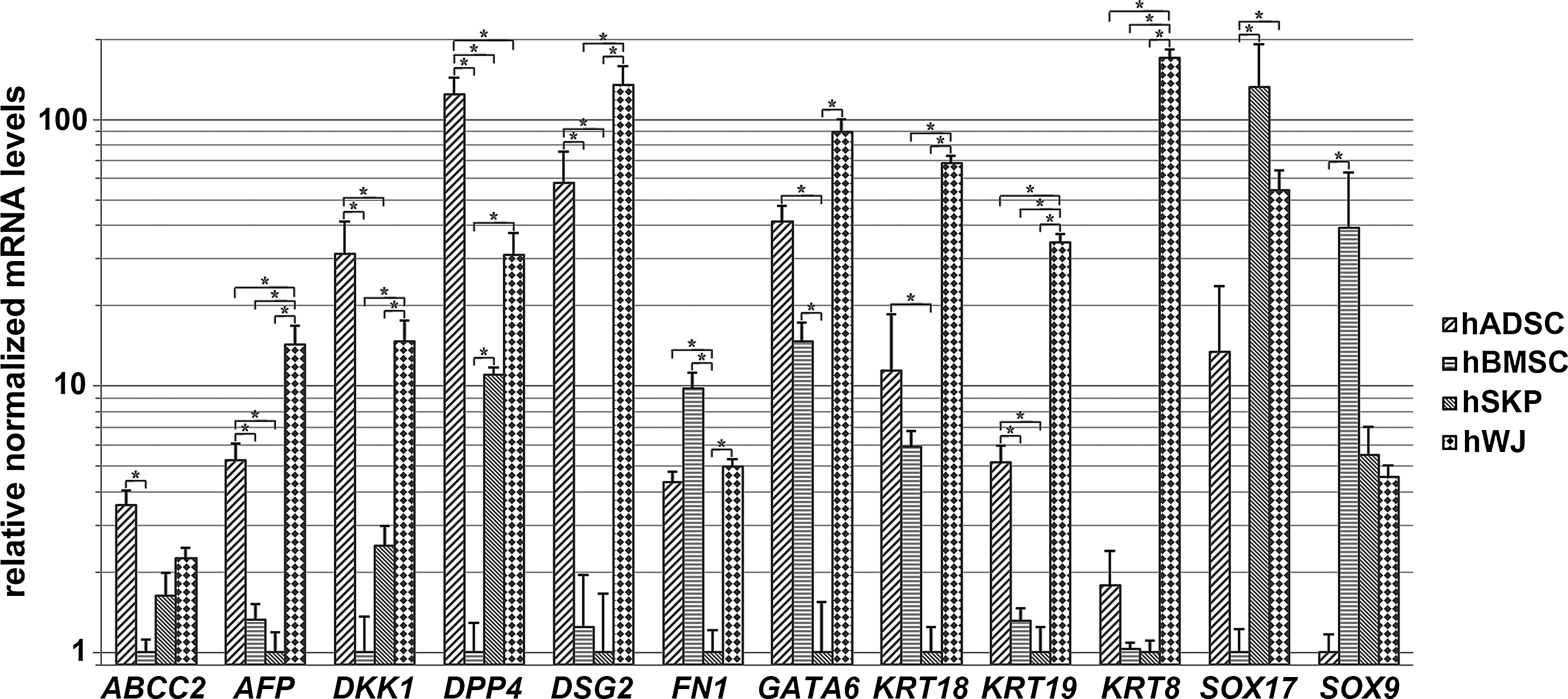
Differential expression of genes involved in endodermal development is confirmed by quantitative real-time PCR analyses. Fold changes of genes involved in endodermal development and commitment are determined for all 4 mesoderm-derived stem cell types (nhADSCs=4; nhBMSCs=3; nhSKPs=4; nhWJs=3). *Significantly increased or decreased mRNA expression between mutually compared stem cell types (fold change >2; P value<0.05). ABCC2, ATP-binding cassette sub-family C member 2; AFP, alpha-fetoprotein; DKK1, dickkopf homolog 1; DPP4, dipeptidylpeptidase 4; DSG2, desmoglein 2; FN1, fibronectin 1; GATA6, GATA-binding protein 6; KRT, keratin; SOX, sex-determining region Y-box.
No significant gene enrichment in the functional gene class (B-H P value>0.05).
Functional analyses are performed using Ingenuity Pathways Analysis (IPA, version SEP 2011; Ingenuity Systems) using B-H multiple-testing-corrected P values (B-H P value<0.05) to identify enriched functional gene classes. Only significantly different expressed genes are used to identify the functional gene classes (fold change >2; P value<0.05). The ratio represents the number of genes increased in the function divided by the total number of genes involved in the function (%).
B-H P value, Benjamini–Hochberg multiple-testing-corrected P values; hADSC, human adipose-derived stromal cell; hBMSC, human bone marrow-derived stromal cell; hSKP, human (fore)skin-derived precursor cell; hWJ, human Wharton's jelly-derived mesenchymal stem cell.
No significant gene enrichment in the functional gene class (B-H P value>0.05).
Functional analyses are performed using Ingenuity Pathways Analysis (IPA, version SEP 2011; Ingenuity Systems) using B-H multiple testing-corrected P values (B-H P value<0.05) to identify enriched functional gene classes. Only significantly different expressed genes are used to identify the functional gene classes (fold change >2; P value<0.05). The ratio represents the number of genes increased in the function divided by the total number of genes involved in the function (%).
Functional analyses are performed using IPA (version SEP 2011; Ingenuity Systems) using B-H multiple-testing-corrected P values (B-H P value<0.05) to identify enriched functional gene classes. Only significantly different expressed genes are used to identify the functional gene classes (fold change >2; P value<0.05). The ratio represents the number of genes increased in the function divided by the total number of genes involved in the function (%).
Neonatal stem cells derived from Wharton's jelly express several major genes involved in liver development
The intrinsic endodermal commitment of the 4 mesoderm-derived stem cell populations has been determined by evaluating the expression profile of genes involved in the development of the liver, the pancreas, and the intestine. Genome-wide microarray analyses reveal a close proximity between on one hand hADSCs and hWJs and on the other hand hBMSCs and hSKPs (Fig. 5). Quantitative real-time PCR analyses show a significantly increased expression of genes involved in endodermal ontogeny in hADSCs and hWJs, including alpha-fetoprotein (AFP), dickkopf homolog 1 (DKK1), dipeptidylpeptidase 4 (DPP4), desmoglein 2 (DSG2), fibronectin 1 (FN1), GATA6, keratin 18 (KRT18), and KRT19 (Figs. 5 and 6). In addition, a significantly higher expression of the gene encoding for the hepatic drug transporter multidrug resistance-associated protein 2 [ATP-binding cassette subfamily C member 2 (ABCC2)] is found in hADSCs, whereas a significantly increased gene expression of KRT8 and sex-determining region Y-box 17 (SOX17) is observed in hWJs (Figs. 5 and 6). Furthermore, GO analyses of hWJs show significant enrichment of functional gene classes involved in liver development. Approximately 25% of the genes involved in proliferation of hepatocytes and 40% of the genes involved in development of liver are significantly increased in hWJs compared to hADSCs and hSKPs, respectively (Table 2). In contrast, GO analyses of hBMSCs compared to hWJs show a significant enrichment of functional gene classes involved in liver regeneration and glucose metabolism, including proliferation of liver cells, regeneration of liver, and gluconeogenesis of liver (Table 3). In addition, increased expression of SOX9 is observed in hBMSCs (Fig. 6).
Skin-derived precursor cells express high levels of genes involved in neurogenesis and skin development
To determine the intrinsic ectodermal commitment of the 4 mesoderm-derived stem cell types, gene expression signatures for neural (crest) development and commitment were used. As such, genome-wide microarray analyses reveal a closer proximity between hBMSCs and hWJs than between any other combination. hSKPs show the highest expression of neural (crest)-related genes, whereas hADSCs the lowest (Fig. 3). Quantitative real-time PCR analyses point to a significantly increased expression of genes involved in ectodermal ontogeny in hSKPs, including nestin (NES), PAX3, activating enhancer-binding protein 2-alpha (TFAP2A), TWIST1, and VIM (Figs. 3 and 4). In addition, compared to the other mesoderm-derived stem cell populations, hSKPs maintain a similar expression level of other genes related to neural (crest) and skin development, including MSX1 and MSX2, snail homolog 1 (SNAI1), and SNAI2 (Figs. 3 and 4). Only hBMSCs show a significantly higher expression of SNAI1 (Figs. 3 and 4). Furthermore, GO analyses of hSKPs reveal a significant enrichment of functional gene classes associated with neural and skin development, including development of brain (15%), neurogenesis (10%), outgrowth of neurites (11%), skin development (14%), proliferation of skin cell lines (35%), and proliferation of keratinocytes (15%) (Table 1). Additionally, GO analyses also show a significantly decreased expression of genes involved in neural development in hADSCs, including neurogenesis (17%), development of brain (24%), growth of neurites (17%) and outgrowth of neurites (17%), compared to hBMSCs (Table 3), and neurogenesis (22%), growth of neurites (22%), and outgrowth of neurites (23%), compared to hWJs (Table 2).
Discussion
The mesoderm, the middle of the 3 germ cell layers of a developing embryo, forms the origin of today's most widely used and characterized hASC populations. Bone marrow- and adipose-derived stromal cells, hSKPs isolated from ventral trunk skin, and Wharton's hWJs trace back to the extraembryonic mesoderm [4,17,18]. Due to their multipotent character as well as their favorable immunological properties, these nonhematopoietic mesoderm-derived hASCs have become an attractive cell source for tissue regeneration and engineering, and for developmental biology studies in which the formation of tissues and organs is investigated [14 –16,33]. A need for reliable in vitro models from human origin also exists in pharmacotoxicological research and testing. Although these 4 stem cell populations share a similar developmental mesoderm background and are able to differentiate into multiple cell types, variations in their functionality, clinical efficacy, and differentiation potential may relate to diverse intrinsic properties specific for each defined stem cell population [5 –7,19 –21]. Therefore, the differential expression of genes and functional gene classes involved in mesodermal, endodermal, and ectodermal development and commitment were investigated in the present study by in-depth comparative transcriptome analyses. First of all, GO analyses revealed no significantly enriched functional gene classes in hADSCs related to mesodermal, endodermal, or ectodermal development (data not shown). Moreover, recent research showed that the expression of genes related to tissue and organ development is significantly lower in hADSCs compared to hBMSCs [34]. This may be explained by the restrictive differentiation potential of hADSCs due to hypermethylation of nonadipogenic lineage-specific promoters [35,36]. Indeed, several reports showed that epigenetic modification of hADSCs is required to cross cell lineage restrictions [37,38]. The necessity to use epigenetic modifiers such as histone deacetylase inhibitors (HDAC-I) and/or DNA methyltransferase inhibitors (DNMT-I) makes the use of hADSCs as a potential stem cell source for clinical treatment less likely.
Bone development
Genome-wide transcriptome and GO analyses revealed that hSKPs are characterized by a significantly increased expression of genes and a significant enrichment of functional gene classes associated with bone development compared to the other 3 mesoderm-derived stem cell populations. Indeed, Lavoie et al. showed that SKPs can generate functional chondrocytes and osteoblasts in vitro and contribute to bone repair in vivo [20]. This is supported by our observations. More specifically, increased expression of genes involved in bone and cartilage development such as BMP2 [39,40] and BMP4 [41,42] was observed. These are both part of the transforming growth factor β (TGFβ) superfamily and play an important role in the development of bone and cartilage and bone mineralization. Also, the neural crest transcription factors PAX3 and TWIST1 have a higher expression. These are involved in the development of craniofacial structures [43,44]. In addition, TWIST1 is also a regulator of RUNX2, a transcription factor that is critical for osteoblast differentiation and skeletal morphogenesis [45,46]. Other transcription factors involved in osteoblast differentiation such as RUNX2 [46], and those involved in craniofacial development such as MSX1 and MSX2 were found to be expressed at comparative levels in the 4 mesoderm-derived stem cells [47,48].
Nervous system and skin development
hSKPs showed a significantly increased expression of genes and enrichment of functional gene classes involved in the development of the nervous system and skin. More specifically, the neural stem cell markers NES and VIM and the neural crest transcription factors PAX3, TWIST1, and TFAP2A, involved in the development of the nervous system and melanocytes, were found to be significantly increased in hSKPs compared to the other mesoderm-derived stem cell populations [49,50]. Interestingly, although hSKPs isolated from trunk skin originate from the lateral-plate mesoderm, it was previously shown that they acquire in vitro properties similar to neural crest-derived hSKPs isolated from facial skin. This includes differentiation into functional osteocytes, chondrocytes, neuronal, and glial cells such as Schwann cells (SCs) [4,20,51,52]. Upon neurogenic stimulation in vitro, SKPs follow an appropriate pattern and time course of neuronal differentiation, with proliferating nestin-expressing SKP-generating postmitotic neuronal cells that coexpress pan-neuronal and peripheral autonomic lineage markers such as βIII-tubulin, peripherin, p75, and tyrosine hydroxylase. These SKP-derived neuron-like cells survive and maintain their peripheral phenotype for at least 5 weeks when transplanted into the central nervous system of a rodent [53]. After transplantation of SKPs in a Parkinson's disease rat model, increased dopamine levels in the striata of the rats were observed, and SKP derivatives found along the tract of transplantation were positive for tyrosine hydroxylase and the dopamine transporter Slc6a3 [54]. SKP also respond to neural crest stimuli such as neuregulins to generate peripheral nervous system glial cells such as SCs [51,55]. These SKP-derived SCs proliferate and induce myelin proteins when in contact with sensory neuron axons in culture and also generate SCs that myelinate central nervous system axons when transplanted into the dysmyelinated brain of neonatal shiverer mice [55]. In addition, SKP-derived SCs survive well within the injured spinal cord, reduce the size of the contusion cavity, myelinate endogenous host axons, and recruit endogenous SCs into the injured cord [51]. They may also bridge the lesion site, increase the size of the spared tissue rim, myelinate spared axons within the tissue rim, reduce reactive gliosis, and provide an environment that is highly conducive to axonal growth [51]. It could also be demonstrated that dermal stem cells, although generated by distinct convergent developmental pathways in vivo, produced SKPs with almost indistinguishable phenotypes [4].
These data suggest that hSKPs, isolated from ventral trunk skin, are primed to differentiate into cells involved in bone, skin, and nervous system formation due to their default intrinsic properties. This confirms the neural-crest-related differentiation potential of (fore)skin-derived hSKPs. More specifically, the neural crest is a transient embryonic structure that gives rise to more than 100 different cell types, including the peripheral nervous system (peripheral neurons and glial cells such as SCs) and several non-neural cell types such as smooth muscle cells of the cardiovascular system, melanocytes of the skin, craniofacial bone, cartilage, and the adipose tissue of the salivary glands [56].
Furthermore, differentiation of hBMSCs into neuron-like cells expressing markers typical for mature neurons has also been reported [57,58]. However, the lack of voltage-gated ion channels necessary for generation of action potentials suggests that these cells actually may not be classified as true neurons [59]. A similar (limited) neuronal differentiation capacity is observed for hWJs [60]. In contrast, hADSCs showed a significantly decreased expression of genes involved in neural development, making them a less interesting source to produce functional neuronal and glial cell types when compared to the other investigated adult stem cell populations. Indeed, under neuroinductive conditions, hADSCs can differentiate into cells expressing the neuronal differentiation marker βIII-tubulin [61] and are able to develop a neuronal phenotype and a positive staining for glial fibrillary acidic protein, S100B, p75, nestin, NeuN, and intermediate filament M [62]. Yet, no functionally mature neurons or glial cells have been generated from hADSCs so far [63].
Cardiovascular system development
Another interesting observation was the significantly increased expression of genes and the enrichment of functional gene classes involved in cardiovascular system development in both hBMSCs and hWJs. More specifically, ENG, a member of the TGFβ receptor superfamily [64], GJA1, the major gap-junction protein in the heart [65], VCAM1, which is involved in the development of vascular endothelium [66], and GATA6, a critical transcription factor for cardiac development [67] are all significantly increased in hBMSCs. Bone marrow stromal cells have indeed been reported of being capable to generate myoblasts that fuse into rhythmically beating myotubes upon treatment with DNMT-I 5-azacytidine [68] and have a potential in vascular remodeling and angiogenesis [69]. On the other hand, a highly increased expression of GATA6, HAND1, which plays a critical role in heart development [70], ICAM1, which is involved in the ontogeny of endothelial cells [71], and VCAM1 is observed in hWJs. Although, very few data are currently available showing the capacity of hWJs to differentiate into cardiomyocyte-like cells [72]. The cardiogenic differentiation potential of hWJs has been indicated so far by the formation of myotube structures; [73] the synthesis of sarcomeric actinin, troponin I, troponin T, and N-cadherin [74]; and spontaneous beating after 21 days of differentiation. In addition, it was recently shown that hWJs can transdifferentiate toward vascular endothelial cells [75]. These data support our findings, suggesting that hBMSCs and hWJs could be the most appropriate mesoderm-derived stem cells to differentiate into cells involved in cardiovascular system development such as cardiomyocytes and endothelial cells.
Liver development
Genome-wide transcriptome and GO analyses also revealed that hWJs have a significantly increased expression of genes and a significant enrichment of functional gene classes associated with liver development. These genes include AFP, the major plasma protein produced in the liver during fetal life [76], DKK1, an inhibitor of the WNT-signaling pathway critical for endodermal development [77], DPP4, an early endoderm marker that is also involved in glucose metabolism [78], DSG2, an important component of desmosomes in the colon and other epithelial cell types [79], and FN1, which plays a role in liver progenitor migration during embryonic development [80]. Furthermore, GATA6, a transcription factor also critical for embryonic liver development [76], the cytokeratins KRT8, KRT18, and KRT19, which encode for the major intermediate filament proteins of epithelial cells, including the liver, pancreas, and bile duct [81], and SOX17, a key transcription factor required for normal development of the definitive gut endoderm [82] are also found to be significantly increased in hWJs. In addition, differentiation of hWJs into hepatocyte-like cells was reported by several research groups [83 –86]. More specifically, Campard et al. showed the successful hepatic differentiation of hWJs that exhibited a hepatocyte-like morphology, upregulated hepatic markers, stored glycogen, produced urea, and exhibited an inducible cytochrome P450 3A4 activity, without the use of epigenetic modifiers [83]. In contrast, epigenetic modification is required to differentiate hADSCs [37,87,88] and hBMSCs [2] into functional hepatocyte-like cells, whereas no mature hepatocyte-like cells have been generated from hSKPs so far [5]. These data support our findings, suggesting that hWJs could be the most promising mesoderm-derived stem cells to differentiate into hepatocyte-like cells.
Conclusion
In conclusion, 4 different mesoderm-derived stem cells were isolated and cultured using the most generally applied isolation methods and culture conditions that are considered to be specific for each of the investigated stem cell types. Using this methodology, hSKPs came out as being the most appropriate stem cells for bone-, nervous system-, and skin-based applications. hWJs seem promising stem cells for angiogenesis, cardiogenesis, and hepatogenesis applications, whereas hBMSCs seem promising for angiogenesis and, to some extent, hepatogenesis. For hADSCs, no well-defined intrinsic preferences for differentiation-specific cell types could be observed. In our opinion, these data suggest that the intrinsic gene expression of the undifferentiated stem cell type has an important impact on its overall differentiation potential. The factors that define these intrinsic properties, however, still remain to be determined. It can be argued that the tissue of origin, the isolation protocol as well as the expansion medium, and to a limited extent, the donor's age and gender will have a significant impact. To understand what factors seem to be of most importance, future experiments in which (i) the isolation method and (ii) the expansion medium are harmonized and standardized are required. To determine the most suitable differentiation strategy to generate the human target cells of interest from adult stem cells, several aspects need to be taken into account. These consist of careful selection of the most suitable postnatal stem cell population based on its intrinsic properties, that is, the expression of key transcription factors and growth factor receptors, related to embryonic development and lineage commitment. Based on this knowledge, the most suitable differentiation protocol should be designed in such a way that it can be tailored to the selected stem cell type.
Footnotes
Acknowledgments
The authors thank Prof. Dr. P. Wylock (UZ-Brussels, Department of Plastic Surgery) and Dr. P. Willekens and V. Van den Borre (ATLAS kliniek) for kind donation of human adipose tissue samples and Dr. V. De Boe (UZ-Brussels, Department of Urology) for donation of human foreskin tissues, upon informed consent of the involved patients or parents when applicable.
Financial Support
Joery De Kock is a doctoral research fellow of the Institute for the Promotion of Innovation through Science and Technology in Flanders (IWT-Vlaanderen). The research leading to these results has also received funding from Grants of the Research Council (OZR) of the Vrije Universiteit Brussel and the European Community's Seventh Framework Programme (FP7/2007–2013) under the grant agreement no. 20161 (ESNATS) and from ISRIB (Brustem) and BELSPO (IAP-HEPRO).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.