
Abstract
The high regenerative capacity of liver contributes to the maintenance of its size and function when injury occurs. Partial hepatectomy induces division of mature hepatocytes to maintain liver function, whereas severe injury stimulates expansion of undifferentiated hepatic precursor cells, which supply mature cells. Although several factors reportedly function in liver regeneration, the precise mechanisms underlying regeneration remain unclear. In this study, we analyzed expression of nucleostemin (NS) during development and in injured liver by using transgenic green fluorescent protein reporter (NS-GFP Tg) mice. In neonatal liver, the hepatic precursor cells that give rise to mature hepatocytes were enriched in a cell population expressing high levels of NS. In adult liver, NS was abundantly expressed in mature hepatocytes and rapidly upregulated by partial hepatectomy. Severe liver injury promoted by a diet containing 3,5-diethoxycarbonyl-1,4-dihydrocollidine induced the emergence of NS-expressing ductal epithelial cells as hepatic precursor cells. NS knockdown inhibited both hepatic colony formation in vitro and proliferation of hepatocytes in vivo. These data strongly suggest that NS plays a critical role in regeneration of both hepatic precursor cells and hepatocytes in response to liver injury.
Introduction
T
Nucleostemin (NS) is a GTPase that binds to p53 and was originally reported to be highly expressed in stem cells from several tissues, including embryonic stem (ES) cells, immature hematopoietic cells, and neural stem/progenitor cells [7]. NS loss results in reduced cell proliferation and increased apoptosis in both ES cells and ES cell-derived neural stem/progenitor cells [8]. Structural comparisons have been used to isolate NS homologues in Caenorhabditis elegans [9], newt [10], Xenopus [11], mouse [7], and human [12]. In the regenerating newt lens, NS protein rapidly accumulates in nucleoli of dedifferentiating pigmented epithelial cells and multinucleate muscle fibers [10], suggesting that its expression correlates with undifferentiated status in newt cells. In contrast, the NS homologue in Caenorhabditis elegans (nst-1) is expressed in both proliferating and differentiated cells. Nst-1 mutants exhibit defects in larval growth and cell cycle progression in germline stem cells [9]. Because nst-1-mutant germ cells can still differentiate into mature sperm, nst-1 may play a critical role in germline stem cell proliferation but not in differentiation. In addition, NS is reportedly expressed at similar levels in non-proliferating muscle stem cells (satellite cells), rapidly proliferating precursor cells (myoblasts), and post-mitotic terminally differentiated cells (myotubes and myofibers) [13]. NS downregulation inhibits differentiation of myoblasts to myotubes, suggesting a role in post-mitotic terminal differentiation in this type. Thus, NS has pleiotropic effects on cellular function, and it is unclear how NS is involved in cell differentiation.
In a previous study, we generated a reporter system where the NS promoter drives green fluorescent protein (GFP) expression (termed NS-GFP) in vivo [14]. We successfully used this reporter system to identify a specific fraction of neonatal germ cells as spermatogonial stem cells with long-term repopulating capacity. We also combined the NS reporter system with a mouse brain tumor model and demonstrated the existence of an undifferentiated tumor-initiating cell (TIC) population in a highly aggressive brain tumor by analyzing GFP fluorescence intensity [15]. Consistent with our data, a recent report employing a bacterial artificial chromosome transgenic mouse line expressing GFP from the NS promoter showed that NS-enriched mammary tumor cells are highly tumorigenic in vitro and in vivo [16]. Further, another recent report showed that NS overexpression enhanced tumorigenicity of TICs, increased expression of genes that maintain undifferentiated status, and enhanced radioresistance [17]. These data suggest that NS functions to maintain stem cell properties in malignant cells.
In this study, we examined the expression and function of NS in liver. Interestingly, we found that NS contributes to the proliferation of hepatocytes after partial hepatectomy and to the regenerative capacity of hepatic precursor cells. Our data strongly suggest that NS is essential for injury-induced liver regeneration.
Materials and Methods
Animals
Mice used in this study were on a C57BL/6 background. NS-GFP Tg mice were generated as described previously [14]. Livers were collected at fetal (embryonic day 14.5: E14.5), neonatal (postnatal day 5: P5), and adult (8 weeks old) stages. For experiments involving severe liver injury, adult mice were fed a diet containing 0.1% DDC (Sigma-Aldrich, St. Louis, MO) for 2 weeks [6]. For partial hepatectomy, mice were anesthetized and 70% of the liver was resected. All procedures were performed in accordance with the animal care guidelines of Kanazawa University.
Isolation of liver cells
For digestion of fetal or neonatal liver cells, livers were minced and dissociated with enzyme-based dissociation buffer (Invitrogen Life Technologies, Carlsbad, CA) as described previously [18]. For isolation of adult liver cells, a 2-step perfusion method was utilized [19]. Briefly, perfusion collagenase solution (0.5 g/L; Sigma-Aldrich) was administered to a sacrificed mouse via the portal vein. Nonparenchymal cells were separated from parenchymal cells by centrifugation (50 g, 1 min), and dead cells were removed by centrifugation through 25% Percoll solution (GE Healthcare, Tokyo, Japan).
Flow cytometry
Single-cell suspensions from fetal, neonatal, or adult liver were incubated with an anti-CD16/CD32 antibody on ice for 10 min, followed by incubation with phycoerythrin (PE)-conjugated anti-TER119 and anti-CD45 antibodies (BD Pharmingen, San Diego, CA) on ice for 30 min. Cells were washed thrice in staining solution [2% fetal calf serum (FCS)/phosphate-buffered saline (PBS)] and incubated with anti-PE microbeads (Miltenyi Biotech, Bergisch Gladbach, Germany). After 3 washes, CD45–TER119–cells were collected (MACS; Miltenyi Biotech). Fetal liver cells were incubated with a biotin-conjugated anti-Dlk antibody (MBL, Nagoya, Japan), followed by incubation with allophycocyanin-conjugated streptavidin antibody (BD Pharmingen). Dead cells were stained with propidium iodide. Fluorescence-labeled cells were analyzed and sorted with JSAN (Bay Bioscience Co, Kobe, Japan).
Hepatic colony forming assay
Cells fractionated by flow cytometry were inoculated at 2,500 cells/well into six-well dishes coated with type I collagen (0.3 mg/mL; Nitta Gelatin, Osaka, Japan). The culture medium included Dulbecco's modified Eagle medium (DMEM)/F-12 supplemented with 10% fetal bovine serum, 5 mmol/L HEPES (Wako, Osaka, Japan), 200 μmol/L L-glutamine (Invitrogen Life Technologies), 50 μmol/L 2-mercaptoethanol (Sigma-Aldrich), 10 mmol/L nicotinamide (Sigma-Aldrich), 10−7 mol/L dexamethasone (Sigma-Aldrich), 1 mg/L insulin (Wako), 1× penicillin/streptomycin (Invitrogen Life Technologies), 50 ng/mL HGF (Peprotech, Rocky Hill, NJ), and 20 ng/mL EGF (Sigma-Aldrich).
Immunohistochemical analyses
Adult liver tissues were fixed with 4% paraformaldehyde at 4°C overnight and embedded in paraffin. Frozen sections were sliced and then fixed with 4% paraformaldehyde. The following primary antibodies were used: goat anti-NS (1:200; R&D Systems, Inc., Minneapolis, MN), rabbit anti-GFP (1:500; Invitrogen Life Technologies), and mouse anti-Ki-67 (1:200; BD Pharmingen). Sections were incubated with primary antibodies for 16 h at 4°C, followed by incubation with the appropriate Alexa Fluor dye conjugated to anti-goat IgG, anti-rabbit IgG, or anti-mouse IgG secondary antibodies (all 1:200; Molecular Probes, Inc., Eugene, OR). Staining was visualized using confocal microscopy (FV1000; Olympus, Tokyo, Japan). For some experiments, primary antibodies were detected using peroxidase-conjugated secondary antibodies (GE Healthcare, Amersham, Buckinghamshire, UK) in combination with a 3, 3′-diaminobenzidine (DAB) Peroxidase Substrate Kit (Vector Laboratories, Burlingame, CA). Sections were counterstained with Mayer's hematoxylin and analyzed using a microscope (Ax80; Olympus).
Immunocytochemical analyses
Hepatic colonies or cell lines were fixed with 4% paraformaldehyde for 10 min, followed by incubation with goat anti-albumin (1:100; Bethyl Laboratories, Montgomery, TX), rabbit anti-cytokeratin 19 (1:1,000, a gift from Dr. Atsushi Miyajima), chicken anti-GFP (1:500; AVES, Tigard, OR), and/or mouse anti-Ki-67 (1:200; BD Pharmingen) at 4°C overnight and then stained with Alexa 546–conjugated and/or Alexa 488–conjugated secondary antibodies.
Western blotting analyses
Liver samples were lysed with sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis (PAGE) sample buffer, sonicated, boiled, and used as total liver cell lysates. Protein concentrations were measured by the bicinchoninic acid (BCA) protein assay (Pierce, Rockford, IL), and equal amounts of protein were separated by SDS-PAGE and transferred onto polyvinylidene difluoride (PVDF) membranes. Membranes were blocked with 5% skim milk in PBS containing Tween 20 for 1 h at room temperature. Membranes were then incubated with a goat anti-NS antibody (1:1,000; Neuromics, Edina, MN) for 16 h at 4°C and a mouse anti-β-actin antibody (1:1,000; Sigma-Aldrich) for 1 h at room temperature. Immune complexes were detected using peroxidase-conjugated secondary antibodies (1:1,000; GE Healthcare and DAKO, Glostrup, Denmark) for 30 min at room temperature and the ECL Prime western blotting detection system (GE Healthcare).
Reverse transcription–polymerase chain reaction
An RNeasy Mini Kit (Qiagen GmbH, Germany) was used in accordance with the manufacturer's instructions to extract total RNA from nonparenchymal cells sorted by fluorescence-activated cell sorting from adult mice treated with DDC. The primers used were as follows: GAPDH (5′-ACCA CAGTCCATGCCATCAC-3′ and 5′-TCCACCACCCTGTTG CTGTA-3′), NS (5′-TCGGAGTCCAGCAAGCATTG-3′ and 5′-GCAGCACTTTCCACATTTGGG-3′), CK19 (5′-GTCCTAC AGATTGACATTGC-3′ and 5′-CACGCTCTGGATCTGTGA CAG-3′), EpCAM (5′-AGGGGCGATCCAGAACAACG-3′ and 5′-ATGGTCGTAGGGGCTTTCTC-3′), Prominin1 (5′-GTA CCTCAGATCCAGCCAGCAA-3′ and 5′-ATTCTTCCAGCT TGGGCAGC-3′), and CD44 (5′-GGCTTTCAACAGTACC TTAC-3′ and 5′-TGAAGCAATATGTGTCATAG-3′).
Lentiviral transduction of short hairpin RNA
To downregulate NS in hepatic precursor cells or mouse hepatic cell lines (Hepa1-6, a mouse hepatocellular carcinoma cell line and BNL C1. 2, a mouse embryonic liver cell line), lentiviruses carrying short hairpin RNA (shRNA) against NS was prepared as previously described [14]. Oligonucleotides encoding shRNA directed against mouse NS mRNA were synthesized as follows: NS #1: sense, AGTAGA AATTTGATGGGCA; antisense, AGCAGAAACTTGATAG GCA; NS #2: sense, GAGGAAAGTTGTTTCGTTA; antisense, GAAGAAAGTTGTTCCATTA. Hepatic colonies derived from Dlk+ fetal liver cells or cell lines were infected with lentivirus for 12 h, followed by washes with PBS, and incubation with culture medium. Cell lines were cultured with 10% FCS/DMEM (Invitrogen Life Technologies).
NS knockdown by hydrodynamic shRNA injection
In vivo transfection of shRNA plasmids into hepatocytes was performed by hydrodynamic injection using 6-week-old mice 3 days prior to partial hepatectomy, in accordance with a previous report [20]. A 27-gauge needle was used to inject 40 μg of plasmid in 2 mL PBS through the tail vein within 10 s. Three days later, the animals were sacrificed and the livers were fixed with 4% paraformaldehyde in PBS and embedded in paraffin for sectioning.
Statistical analyses
Statistical differences were determined using the unpaired Student's t-test for P values.
Results
NS expression correlates with the colony-forming capacity of hepatic precursor cells in developing liver
To investigate NS expression in developing liver, we evaluated GFP intensity in liver cells of NS-GFP Tg fetuses (E14.5) and neonates (P5). Flow cytometry analysis of fetal liver cells showed that most CD45−Ter119− (nonhematopoietic) cells expressed high GFP levels (Fig. 1A). Although GFP levels were very high in Dlk+ cells, in which hepatic stem/precursor cells are enriched [21], those levels were only slightly higher than those in Dlk− cells (Fig. 1A). Thus, GFP expression was not indicative of a particular subpopulation in fetal liver. Interestingly, however, NS-GFP neonatal liver cells fell into distinct populations based on GFP fluorescence intensity (Fig. 1B). While most non-hematopoietic cells were GFPmid, we found a distinct GFPhigh population. The proportions of GFPlow and GFPneg cells were very small. To determine the potential functional significance of these subpopulations, we evaluated hepatic colony forming ability. GFPhigh cells generated colonies at higher frequency than did any other cell population (Fig. 1C). Most colonies derived from GFPhigh cells were CK19+ or albumin+ hepatocytes (Fig. 1D), suggesting that NS is an indicator of hepatic precursor cells in neonatal liver.
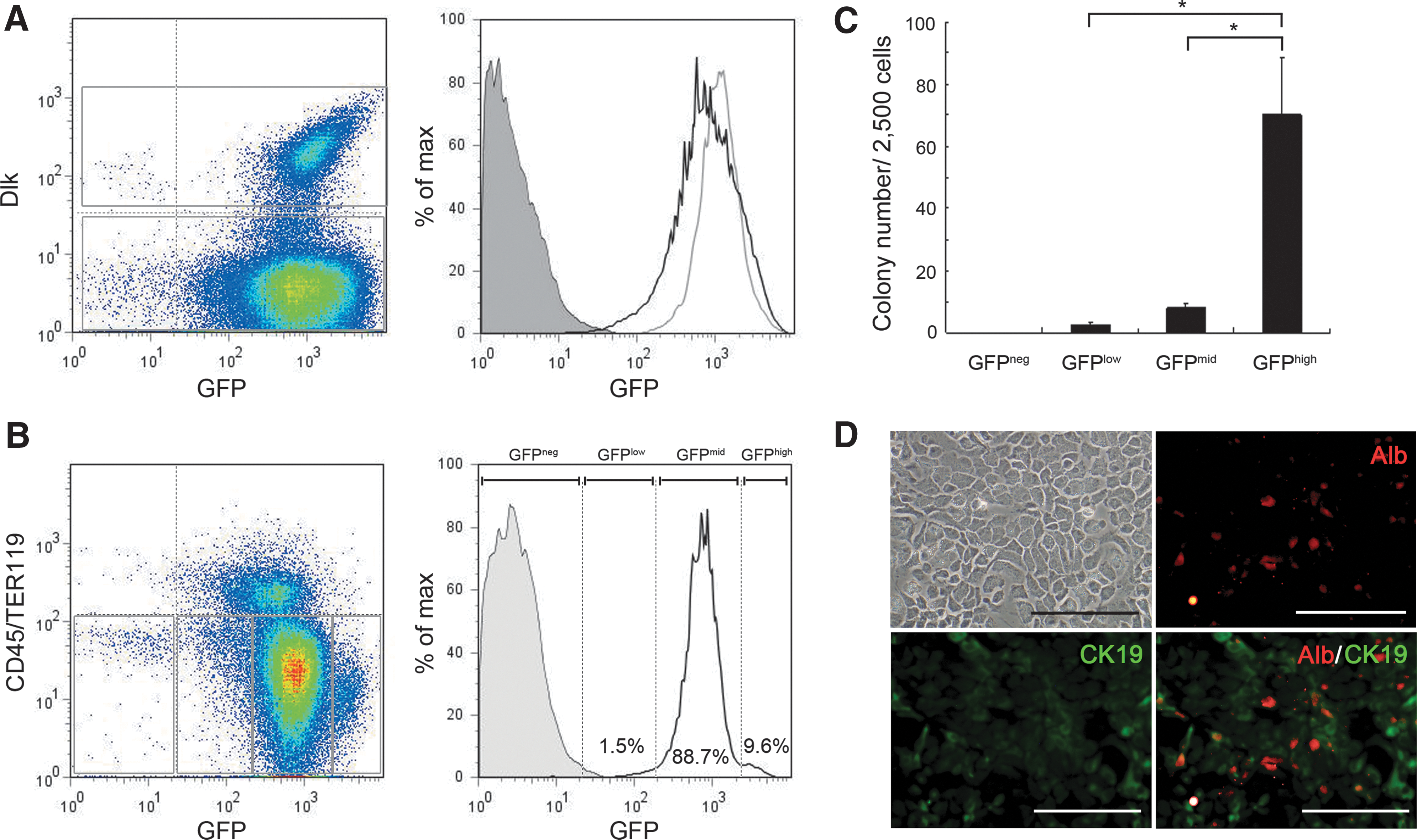
Correlation of nucleostemin (NS) expression with colony-forming capacity of hepatic precursor cells in developing liver.
Partial hepatectomy upregulates NS expression in hepatocytes
We next examined NS-GFP expression in adult liver. We found that NS-GFP is highly expressed in hepatocytes. In particular, hepatocytes near central veins showed higher levels of NS-GFP expression than did those in the portal area (Fig. 2A–C). In addition, we found that NS-GFP is also highly expressed in bile duct epithelial cells (Fig. 2D, E). Next, we asked whether expression levels of NS-GFP are altered by liver injury. After partial hepatectomy, NS-GFP was upregulated within 24 h (Fig. 3A i–iv). Immunohistochemical analyses showed that NS protein expression was remarkably increased in nucleoli 24 h after hepatectomy. Although there were variations among samples, NS protein levels appeared to remain elevated at day 3 (Fig. 3B) and reverted to baseline levels at day 7, whereas NS-GFP expression was already downregulated at day 3. Western blotting analysis using an anti-NS antibody consistently showed an increase in endogenous NS protein on day 1 and day 3 after partial hepatectomy (Fig. 3C). The discrepancy between NS-GFP and endogenous NS is possibly due to differences in protein stability stemming from different post-translational modification of these molecules. Interestingly, partial hepatectomy induced a variant form of NS that is reported to be expressed in particular tissues [22,23]. Hepatocytes that had begun to proliferate showed a small increase in the expression of Ki-67, a marker of cell proliferation, at day 1, and further increases in Ki-67 expression were observed at day 3 (Fig. 3A v–viii). These data indicate that NS gene expression is rapidly upregulated before the start of cell division in response to partial hepatectomy.

Expression of NS-GFP in hepatocytes and bile duct epithelial cells of adult liver.

Upregulation of NS-GFP in adult hepatocytes in response to partial hepatectomy.

A DDC diet induces emergence of ductal epithelial cells expressing NS-GFP
DDC treatment inhibits the capacity for hepatocytes to regenerate, while inducing ductal proliferation in mice in what is known as the oval cell response [6]. We found that DDC treatment reduced expression of NS-GFP in hepatocytes (Fig. 4A i, ii) compared to the expression in untreated hepatocytes. We also found that bile duct-like NS-GFP-positive cells emerged in the portal zone following DDC treatment (Fig. 4A iii, iv). Interstitial cells surrounding ductal cells did not express NS-GFP. We confirmed by immunofluorescence that NS-GFP was expressed in CK19+ ductal epithelial cells (Fig. 4B). To investigate the regenerative capacity of NS-GFP-expressing cells in DDC-treated liver, we evaluated NS-GFP intensity in nonparenchymal cells, since oval cells reportedly reside in that population [6]. Flow cytometry analysis showed that most CD45−Ter119− nonparenchymal cells were GFP-positive (Fig. 5A), although the intensity of NS-GFP expression in non-parenchymal cells in adult mice appeared lower than that seen in developing liver cells. These NS-GFP-positive cells fell into GFPhigh and GFPlow populations. GFPhigh cells were relatively rare, but only GFPhigh cells showed hepatic colony forming ability (Fig. 5C). In contrast, no colonies were generated from GFPlow or GFPneg cells. Severe liver injury promoted by a DDC diet increased the proportion of GFPhigh cells relative to the proportion in untreated mice (Fig. 5B). Because we found that ductal cells express NS-GFP (Figs. 2 and 4), we assumed that the DDC diet increased the number of ductal cells, resulting in an increase in the proportion of GFPhigh cells. Hepatic colonies were generated only from GFPhigh cells (Fig. 5D). GFPhigh cells expressed higher levels of NS mRNA, indicating that GFP expression corresponded with that of endogenous NS, and also expressed several genes reportedly expressed in oval cells [19] (Fig. 5E). These data indicate that hepatic precursor cells induced by severe liver injury express NS.
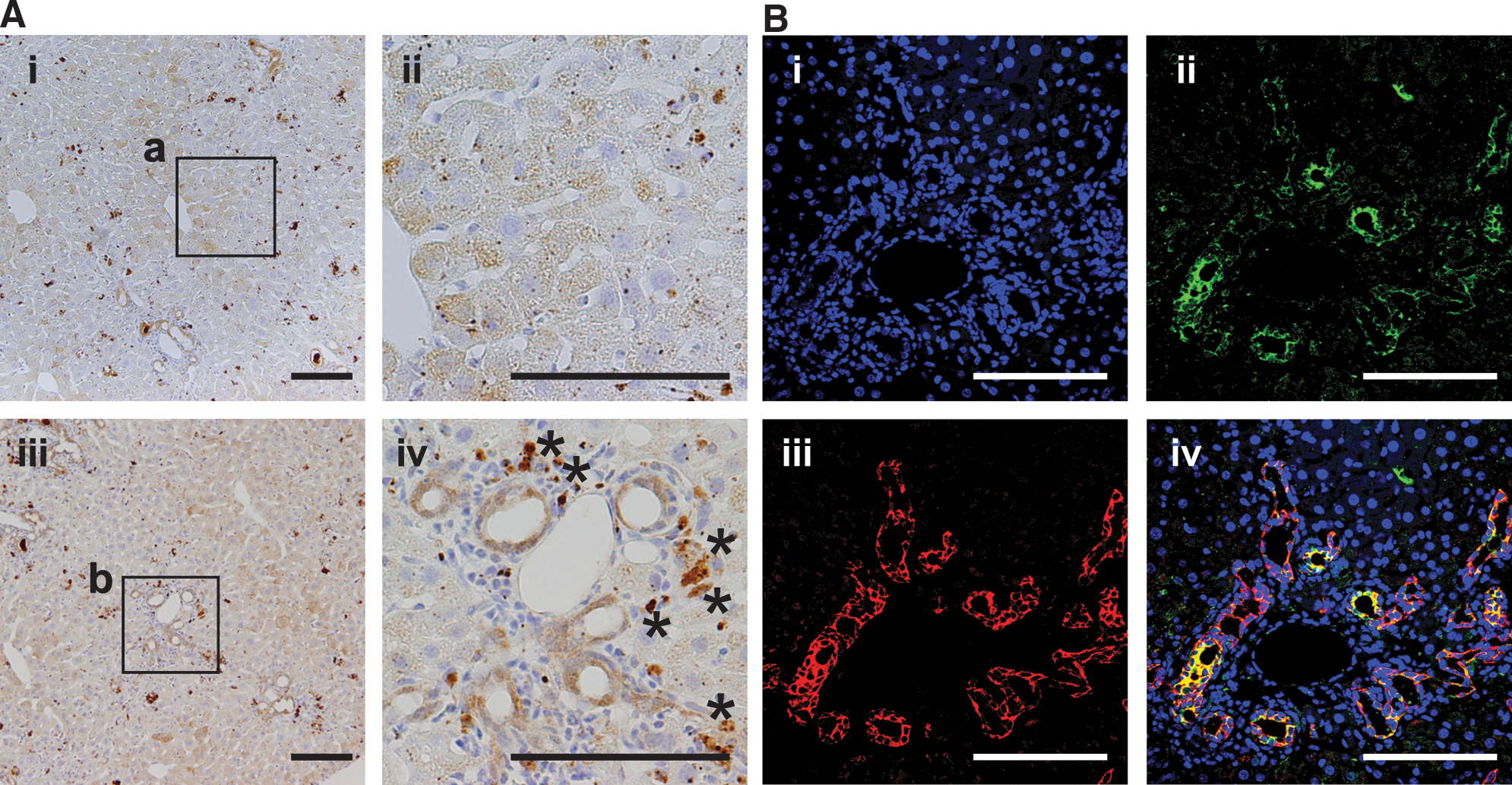
Duct-like cells express NS-GFP in liver of adult mice fed a 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) diet.
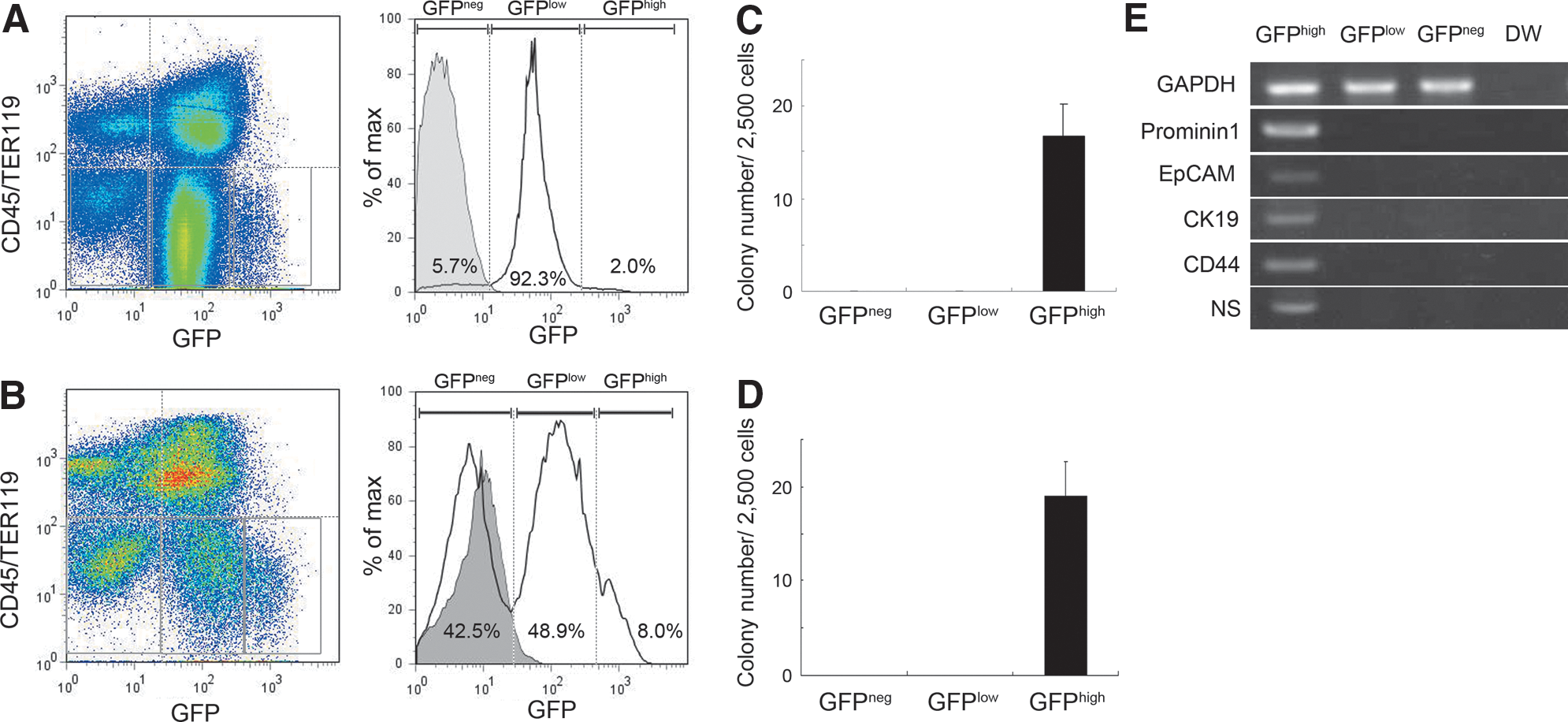
Increased ratios of NS-GFPhigh hepatic precursor cells are seen following a DDC diet.
NS downregulation inhibits proliferation of hepatic precursor cells
Next, to address whether NS is required for regeneration of hepatic precursor cells, we downregulated NS expression in hepatic cell line and primary fetal liver cells in vitro. Previously, we successfully suppressed NS in a germ cell line by infection with a lentivirus carrying NS shRNA [14]. In this system, infected cells were identified by GFP expression driven by the lentivirus vector (GFP+ cells). For the current study, we infected the mouse hepatocellular carcinoma cell line Hepa1-6 and the mouse embryonic liver cell line BNL C1.2 with lentiviruses carrying NS shRNA (#1 or #2) or a scrambled control shRNA and then stained the cells with an anti-NS antibody. Both shRNAs, but not the scrambled control, efficiently reduced expression of NS protein in the hepatic cell lines (Fig. 6A, data not shown). NS knockdown (GFP+) cells in the cell lines had dramatically reduced colony-forming capacity (Fig. 6B, data not shown). We also found that NS downregulation significantly decreased the proportion of Ki-67-positive cells among transfected (GFP+) cells in the hepatic cell line (Fig. 6C) and in the hepatic colonies derived from freshly isolated fetal liver precursor cells (Fig. 6D). Thus, NS downregulation inhibits proliferation of hepatic precursor cells.

Inhibition of proliferation of hepatic precursor cells following NS downregulation in vitro.
NS plays an essential role in hepatocyte proliferation in response to liver injury in vivo
The observation that NS is upregulated in hepatocytes after partial hepatectomy suggested that NS is essential for hepatocyte proliferation. To examine the effect of loss NS function in hepatocytes in vivo, we introduced the shRNA plasmids into liver cells by hydrodynamic injection of plasmid DNA via the tail vein [20]. Partial hepatectomy was performed 3 days later and we found that hepatocytes were successfully transfected with shRNA plasmids by detection of GFP expression. Three days after partial hepatectomy, we found that NS knockdown in the hepatocytes significantly suppressed expression of the Ki-67 antigen (Fig. 7), indicating that NS is essential for hepatocyte proliferation in response to liver injury in vivo.

Inhibition of hepatocyte proliferation following NS downregulation in vivo. NS shRNA plasmids were introduced into liver cells by hydrodynamic injection of the plasmid DNA via the tail vein, followed by partial hepatectomy 3 days after injection. Liver tissue specimens were prepared 3 days later, and sections were stained with an anti-GFP antibody (green), anti-Ki-67 antibody (red), and DAPI (blue). Ki-67 expression was evaluated in more than 60 GFP+ cells for each sample.
Discussion
In this study, we examined the expression and function of NS in developing and injured liver, and we evaluated the capacity of hepatic NS-expressing cells to form colonies by using an NS-GFP system. As previously reported [19], DDC treatment induced the emergence of ductal cells that express both cholangiocellular and hepatocytic markers, called “oval cells,” in periportal regions. Several studies have identified markers of oval cells, including Ep-CAM and CD133 [19,24,25]. NS-GFP was not specific for oval cells, since most hepatocytes and nonparenchymal cells also expressed GFP. Therefore, NS-GFP expression alone cannot be used to purify hepatic stem/precursor cells. However, since a distinct subpopulation of cells expressing high GFP levels (GFPhigh) showed higher clonogenic potential, combining this system with evaluation of other stem cell markers could enable efficient enrichment of a stem/precursor cell population. In addition, NS may be particularly important for the development of liver, since the expression level of NS-GFP in developing liver cells appeared to be higher than that in adult non-parenchymal cells. NS may therefore play a critical role in expansion of the hepatic stem/precursor cells during liver development.
A previous study demonstrated that NS is required for rRNA processing [9,26], suggesting that NS expression regulates protein synthesis. Enhanced protein synthesis requires activation of ribosomal biogenesis. NS belongs to the class of nucleolar GTPases that includes yeast Nug1, which exports pre-60S ribosomal subunits out of the nucleolus [27]. In Caenorhabditis elegans, nst-1 mutants exhibit reduced rRNA levels, suggesting a critical role of NS in ribosome biogenesis [9]. NS knockdown apparently delays processing of 32S pre-rRNA into 28S rRNA and is accompanied by a substantial decrease in protein synthesis and in the levels of rRNAs and some mRNAs [26]. Because protein synthesis is required for cell growth and proliferation, NS expression in both hepatic precursor cells and hepatocytes may be important for tissue regeneration. On the other hand, protein synthesis appears to be enhanced in resting hepatocytes for reasons unrelated to regeneration. Mature hepatocytes exhibit high levels of protein synthesis to maintain serum protein levels, and protein translation actively occurs even in non-dividing hepatocytes. Thus, NS expression may be controlled by several different signals.
One possible regulator of NS is Myc, which is upregulated by partial hepatectomy [28]. When Myc is overexpressed in mouse hepatocytes in vivo using recombinant adenovirus, hepatocytes enlarge in the absence of significant cell proliferation, an event associated with upregulation of large- and small-subunit ribosomal and nucleolar genes [29]. In addition, a recent study identified the NS gene as a direct transcriptional target of the Myc oncoprotein [22]. Therefore, NS may function to increase cell mass in response to Myc activation following partial hepatectomy. It has also been reported that, constitutive activation of Myc generates hepatocellular carcinoma, whereas Myc inactivation promotes differentiation of tumor cells into hepatocytes and biliary cells, which form bile duct structures [30], suggesting that Myc maintains cells in an undifferentiated status. NS may have a similar function in the case of hepatic malignancy. In addition, Myc may control post-translational regulation of NS protein. A recent study revealed that NS is a target of reactive oxygen species (ROS) [31]. In transformed hematopoietic cells, Myc activation leads to high ROS levels, resulting in impaired NS protein degradation. Therefore, Myc activation may stabilize NS protein in regenerating liver. Because GFP would not be stabilized in the same way as the NS protein, these findings suggest that NS protein levels may not be precisely correlated with NS-GFP levels. Nonetheless, both NS protein levels and NS-GFP expression are consistently upregulated by partial hepatectomy. These findings suggest that overall NS expression is likely regulated by both transcriptional and protein stability.
In conclusion, we have demonstrated that NS is essential for proliferation of both hepatic precursor cells and hepatocytes. Understanding the mechanisms regulating NS expression and function may contribute to development of methodologies useful for enhancing liver regeneration in pathological states.
Footnotes
Acknowledgments
We thank Drs. Akihide Kamiya, Hiromitsu Nakauchi, Tetsuhiro Chiba, Atsushi Iwama, and Atsushi Miyajima for providing information and technical advice for establishing assay systems for hepatic colony formation and isolation of liver cells and Dr. Kenichi Harada for insightful suggestions for histological analyses. A.H. was supported by a Grant-in-Aid for Scientific Research on Innovative Areas and the Project for Development of Innovative Research on Cancer Therapeutics (P-DIRECT) from the Ministry of Education, Culture, Sports, Science and Technology, Japan, and by a grant from the Japan Science and Technology Agency (JST), CREST.
Author Disclosure Statement
No competing financial interests exist.