
Abstract
Ectopically, expression of defined factors could reprogram mammalian somatic cells into induced pluripotent stem cells (iPSCs), which initiates a new strategy to obtain pluripotent stem cell lines. Attempts have been made to generate buffalo pluripotent stem cells by culturing primary germ cells or inner cell mass, but the efficiency is extremely low. Here, we report a successful method to reprogram buffalo fetal fibroblasts (BFFs) into pluripotent stem cells [buffalo induced pluripotent stem cell (biPSCs)] by transduction of buffalo defined factors (Oct4, Sox2, Klf4, and c-Myc) using retroviral vectors. The established biPSCs displayed typical morphological characteristics of pluripotent stem cells, normal karyotype, positive staining of alkaline phosphatase, and expressed pluripotent markers including Oct4, Sox2, Nanog, Lin28, E-Cadherin, SSEA-1, SSEA-4, TRA-1-81, STAT3, and FOXD3. They could form embryoid bodies (EBs) in vitro and teratomas after injecting into the nude BALB/C mice, and 3 germ layers were identified in the EBs and teratomas. Methylation assay revealed that the promoters of Oct4 and Nanog were hypomethylated in biPSCs compared with BFFs and pre-biPSCs, while the promoters of Sox2 and E-Cadherin were hypomethylated in both BFFs and biPSCs. Further, inhibiting p53 expression by coexpression of SV40 large T antigen and buffalo defined factors in BFFs or treating BFFs with p53 inhibitor pifithrin-a (PFT) could increase the efficiency of biPSCs generation up to 3-fold, and nuclear transfer embryos reconstructed with biPSCs could develop to blastocysts. These results indicate that BFFs can be reprogrammed into biPSCs by buffalo defined factors, and the generation efficiency of biPSCs can be increased by inhibition of p53 expression. These efforts will provide a feasible approach for investigating buffalo stem cell signal pathways, establishing buffalo stem cell lines, and producing genetic modification buffaloes in the future.
Introduction
S
Buffalo is one of the most important domestic animals distributed in tropical and subtropical regions, which can provide milk, meat, and draft for agriculture. There are 2 kinds of buffalos classified as swamp and river types. The milk yield of Chinese swamp buffalos is lower than that of river type buffalos [22], and is in urgent need of improving their production traits and disease resistance ability by genetic manipulation technology [23]. Gene modification can be accomplished by transgenic cloning, but gene targeting in somatic cells of large animals is difficult and inefficient. Based on the embryonic stem cells (ESCs), gene targeting has succeeded in mouse [24], which has been considered to be the promising way to generate site-specific gene targeting in large animals [25]. To date, ESC lines have been established in mouse [26], human [27], monkey [28], and rat [29,30], but there is no bonafide large animal ESCs available for the precise gene modification. Although a lot of effort has been made to isolate and culture buffalo ESCs, the colony could not passage for a long time in vitro, and these cells were only defined as ES-like cells [31,32]. The main reason that limited the research of buffalo ESCs and other large animal ESCs may be due to the lack of proper knowledge on the signal pathways, pluripotent markers, and culture conditions [33].
Based on the traditional method, ESC lines derived from the buffalo blastocysts encountered a challenge. In the present study, we tried to reprogram buffalo fetal fibroblasts (BFFs) into pluripotent stem cells using the buffalo defined factors, including Oct4, Sox2, Klf4, and c-Myc (or OSKM). The pluripotent properties of buffalo induced pluripotent stem cells (biPSCs) and methylation pattern of the defined factors were examined. We also tried to improve the efficiency of biPSCs by coexpressoin of OSKM and SV40 large T antigen (T), which were involved in p53 pathway [34] and iPSC reprogramming [35,36]. Finally, the biPSCs were employed as donor cells for SCNT to verify its practical applications for the buffalo genetic manipulation.
Materials and Methods
Plasmid construction and retrovirus packaging
The pMX-based retrovirus vector and packaging cell Plat-GP cells were purchased from Cell Biolabs. The open reading frame of buffalo defined factors (Oct4, Sox2, Klf4, c-Myc, Nanog, and Lin28) were amplified from the buffalo in vitro fertilization (IVF) blastocysts (produced using the method previously reported by our lab [32]) by reverse transcriptase–polymerase chain reaction (RT-PCR). The SV40 large T antigen was amplified from 293-T cells and hTRET was amplified from pCI-neo-hTERT (Promega). Then these factors were cloned into the pMX vector respectively. Internal ribosome entry site-green fluorescent protein (IRES-GFP) cassette were inserted into the downstream of pMX-Oct4, Oct4, and GFP simultaneously expressed under the control of the same promoter. The sequences of primers and reaction conditions were listed in Supplementary Table S1 (Supplementary Data are available online at
Preparation of BFFs
Tissues from the skin of Chinese swamp buffalo fetuses (about 5 months) were enzymatically digested with 0.25% trypsin and 0.05% EDTA for 30 min followed by 0.2% collagenase for 45 min. The disaggregated cells were washed 3 times in Dulbecco's modified Eagle's medium (DMEM; Hyclone) supplemented with 10% fetal bovine serum (FBS; Hyclone) by centrifugation at 500 g for 5 min, and then placed in culture in a 60 mm tissue culture dish (Corning) under a humidified 5% CO2 in air atmosphere at 38.5°C. After 7–10 days of culture, confluent fetal fibroblast monolayers were obtained and then routinely passaged 3–6 times using an enzymatic solution (0.25% trypsin and 0.05% EDTA) for 3 min before using for transduction with pseudovirus.
Retroviral transduction and biPSC culture
The BFFs that had been passaged 3–6 times were infected twice with pseudovirus. At 48 h after transduction, the BFFs were infected with pseudovirus that were trypsinized and plated onto the mouse embryonic fibroblasts (MEF; Cyagen) that had been treated with 10 μg/mL mitomycin C (Sigma) for 3 h at 1×104 cells per 60 mm dish in the high glucose DMEM supplemented with penicillin/streptomycin (Gibco) and 10% FBS. At 24 h after seeding on MEF, the medium was changed with ES medium [high glucose DMEM supplemented with penicillin/streptomycin, 20% ESC FBS, 2 mM
Alkaline phosphatase staining and immunofluorescence assay
To detect alkaline phosphatase (AP) activity, biPSCs were fixed with 4% paraformaldehyde for 30 min at room temperature, washed 3 times with phosphate-buffered solution (PBS), and stained with NBT/BCIP solution (Amresco) for 15–30 min at room temperature. Then, the stained biPSCs were examined under the inverted microscope (Nikon). To examine the expression of pluripotency markers, biPSCs were fixed as the AP staining protocol. For permeabilization, cells were treated with 0.1% Triton X-100 in PBS for 20 min, and then incubated in blocking solution [1% bovine serum albumin (BSA) in PBS] for 30 min. The primary antibodies of anti-Oct4 (1:250; Santa Cruz), anti-Sox2 (1:250; Santa Cruz), anti-Nanog (1:400; Santa Cruz), anti-SSEA1 (1:300; Santa Cruz), anti-SSEA4 (1:300; Santa Cruz), anti-Tra-1-81 (1:250; Santa Cruz), and anti-E-Cadherin (1:250; Santa Cruz) were diluted in blocking solution and incubated with cells overnight at 4°C. On the next day, cells were washed 3 times with PBS (5 min each), and incubated with secondary antibodies at room temperature for 1 h in the dark. Cells were washed 3 times with PBS, and costained with Hoechst (2 μg/mL, Sigma) at room temperature for 10 min. Cell fluorescence was examined using fluorescence microscope (Nikon).
RT-PCR and quantitative real-time PCR
The first-strand cDNA was extracted from BFFs, biPSCs, embryoid body (EB), inner cell mass (ICM) of buffalo IVF blastocysts, and EG-like cells (prepared according to the method reported by our lab [37]) using Cells-to-cDNA™ II Kit (Applied Biosystems). RT-PCR was applied to detect the expression of endogenous pluripotency markers and exogenous genes in biPSCs. Quantitative real-time PCR was carried out using TaqMan primers and probe specific to candidate genes on the ABI PRISM 7500 Real Time System (Applied Biosystems). The Quantitative PCR (QPCR) mixture (20 μL) contained 10 μL 2×Premix Ex Taq (Takara), 0.4 μL 50×Rox reference dye, 0.1 μM of each primer and probe, and 60 ng cDNA. The thermal cycling conditions were 50°C for 2 min, 95°C for 10 min, followed by 40 amplification cycles (95°C for 15 s, 60°C for 1 min). The relative expression level of target genes was calculated by 2−ΔΔCt method relative to the Histone. The details of primers, probes, and reaction conditions for RT-PCR and quantitative real-time PCR are listed in Supplementary Table S1.
Bisulfite genomic sequencing
The CpG islands involved in the promoter fragment of buffalo Oct4, Nanog, Sox2, and E-Cadherin were cloned and sequenced. The DNA of biPSCs was purified using the QIAamp DNA Mini Kit (Qiagen), and treated by bisulfite using Methylamp™ DNA Modification Kit according to the manufacturer's instruction. The methylation primers were designed according to the online software (
Karyotype analysis
The biPSCs that had grown up to 60%–70% confluence were arrested in metaphase by exposing them to 0.2 μg/mL demecolcine for 3 h in a humidified atmosphere of 5% CO2 in air at 37°C, and then resuspended in 0.8% sodium citrate for 30 min at 37°C. After centrifugation at 1,200 rpm for 3 min, cells were fixed with 4 mL acetic methanol (1:3) solution for 20 min at 37°C. Thereafter, chromosome spreads were prepared by dropping 20 μL cells suspension onto cold slides and incubated at 75°C for 3 h. At last, the slides were stained with 10% Giemsa solution for 15 min and analyzed under an Olympus microscope.
Preparation of EB
The biPSCs were digested with 0.25% trypsin and seeded on nonadherent culture plates in the differentiation medium (high glucose DMEM supplemented with 20% FBS, 2 mM
Teratoma formation
To examine the pluripotency of biPSCs in vivo, about millions of trypsinized biPSCs were hypodermically injected into the 4-to-6-week-old BALB/C nude mice. On 6–10 weeks after injection, mice were sacrificed, and teratomas were explanted. The teratoma was fixed with Bouin's solution and embedded in paraffin. Sections were stained with hematoxiline/eosin for histological examination. The DNA was extracted from the remaining teratoma by QIAamp DNA Mini Kit (Qiagen), and amplified to identify the origin of these teratomas by PCR. The sequences of primers specific to buffalo and mouse are listed in Supplementary Table S1.
Reconstruction of nuclear transfer embryos using biPSCs
Reconstruction of nuclear transfer embryos using biPSCs was performed according to the method reported by Shi et al. [22]. In brief, trypsinized biPSCs were transferred into the perivitelline space of enucleated oocytes with a 25-μm micropipette. The couplet was transferred to a droplet of 100 μL of fusion medium (0.28 M mannitol, 0.1 mM CaCl2, 0.1 mM MgSO4, 5 mM HEPES, and 0.1% BSA) overlaid with mineral oil and then placed on the micromanipulator with 2 platinum needle electrodes (0.2 mm apart). The fusion was induced by the application of an alternating current pulse of 2 V for 1 s and then 3 direct current pulses of 1 kV/cm for 15 μs with an ECM2001 Electrocell Manipulator (BTX, Inc.). Couplets were then washed in embryo culture medium (TCM-199 supplemented with 3% fetal calf serum, CM) and incubated in this medium for 30 min at 38.5°C. The fusion of couplets was checked at 200× magnification under an inverted microscope. Three hours after the fusion, the activation of fused embryos was induced by exposure to 5 μM ionomycin in CM for 5 min and subsequent incubation in 2 mM 6-dimethylaminopurine for 3 h at 38.5°C and 5% CO2 in air. After activation, reconstructed embryos were placed in coculture with granulosa cell monolayers in a 30 μL droplet of CM overlaid with mineral oil under a humidified atmosphere of 5% CO2 in air at 38.5°C. The granulosa cell monolayers were established at 48–72 h before introduction of embryos. After introduction of embryos, half of the medium was replaced with fresh medium every 24 h. After 2 days of coculture, cleavage of reconstructed embryos was checked, and the number of developed blastocysts was recorded within 8 days of coculture.
Results
Generation of biPSCs using buffalo defined factors
To generate iPSCs from buffalo somatic cells, 6 transcription factors frequently used in preparing iPSCs were cloned from buffalo IVF blastocysts by RT-PCR, including Oct4, Sox2, Klf4, c-MYC, Nanog, and Lin28 (or OSKMNL, NCBI accession No.: Oct4 JN991003, Sox2 JN986576, Klf4 JN986574, c-Myc JN986573, Nanog JN991004, Lin28, and JN986575). Multiple sequences alignment showed that these genes were highly conserved between buffalo and other species. Then, these 6 buffalo transcription factors were cloned into pMX-based retrovirus vectors respectively. To test the transduction efficiency and the silencing of exogenous genes, GFP-containing retrovirus vector was also constructed and named as pMX-Oct4-IRES-GFP. The result of BFFs infected twice (12 h interval) by retrovirus showed that the infective efficiency of ultracentrifugal retrovirus were higher than fresh retrovirus supernatant (Fig. 1B). Thus, ultracentrifugal retrovirus was employed in the following investigation.
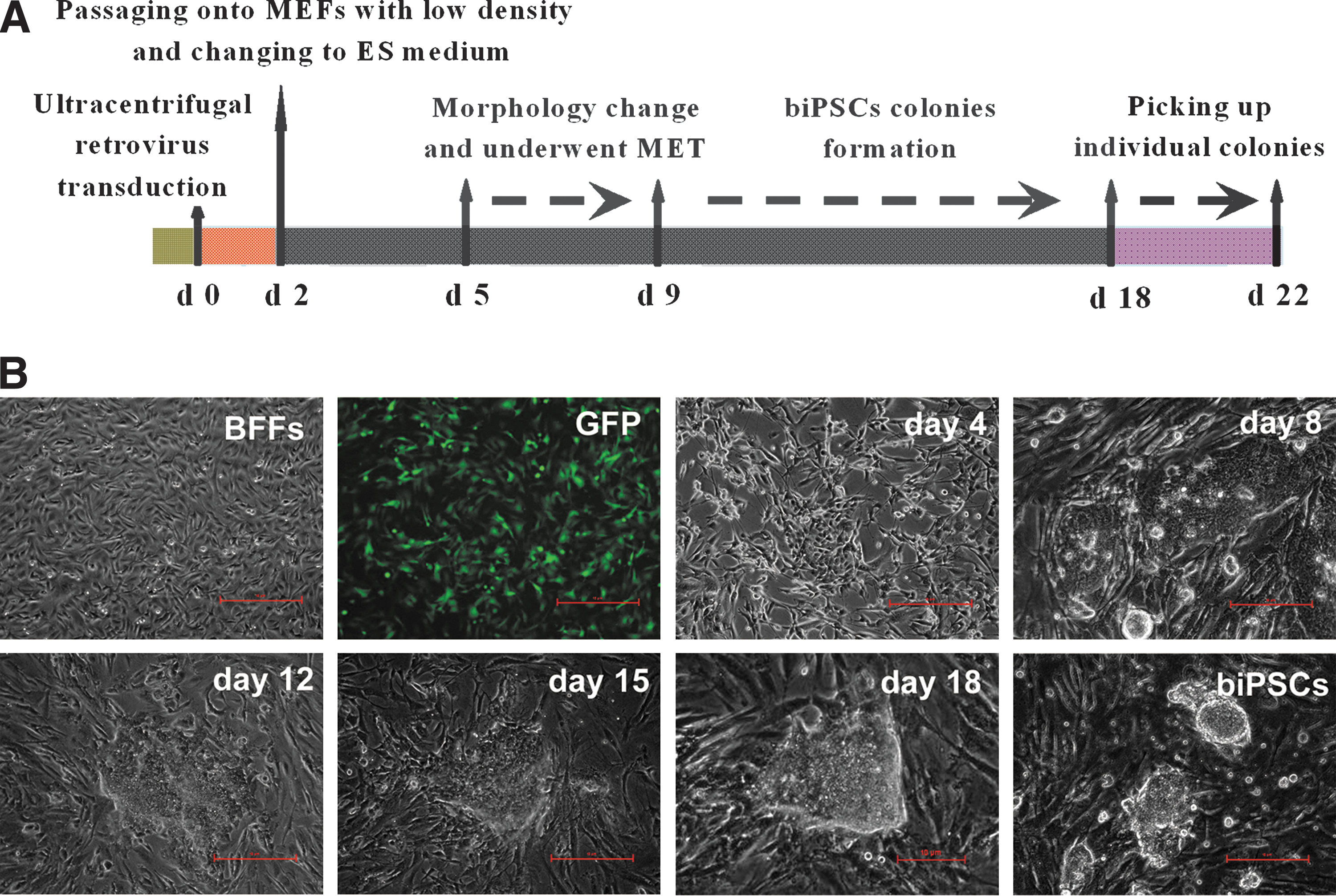
Buffalo induced pluripotent stem cells (biPSCs) generated by retrovirus-based buffalo defined factors.
Two cocktails of retroviruses containing defined factors OSKM or OSKMNL were used to transduce BFFs respectively (Fig. 1A). The infected cells were trypsinized and plated onto MEF feeder at 1×104 cells per 60 mm dish in DMEM supplemented with 10% FBS at 48 h after transduction. At 24 h after seeding on MEF, the medium was changed with ES medium. At 4–5 days after transduction, the infected cells began to change in morphology and proliferated rapidly. Then, the reprogramming fibroblasts transited from mesenchymal to epithelial-like cells, which was known as MET [38,39]. The MET of transduced BFFs was completed around day 8, and the biPSC colonies were observed at 11–14 days after transduction. Around 17–20 days, large round colonies formed with clear boundary to the feeder cells (Fig. 1B). The colonies were picked and cultured in the 96-well plate for further expansion.
When AP staining was applied to identify the positive biPSC colonies, 21 positive colonies induced with OSKM combination and 38 positive colonies induced with OSKMNL combination were obtained. Although the expression of Klf4 and c-Myc were detected in BFFs by RT-PCR and immunofluorescence (data not shown), no biPSCs were obtained when Klf4 and c-Myc were withdrawn from the 2 cocktails (Fig. 2A). More biPSCs (2-fold higher) were generated with combination of 6 defined factors OSKMNL in comparison with OSKM, but none of the colonies generated with the 2 combinations passaged more than 6 generations in the ES medium containing bFGF and LIF. Considering less transgenes, OSKM combination was chosen for the further generation of biPSCs.
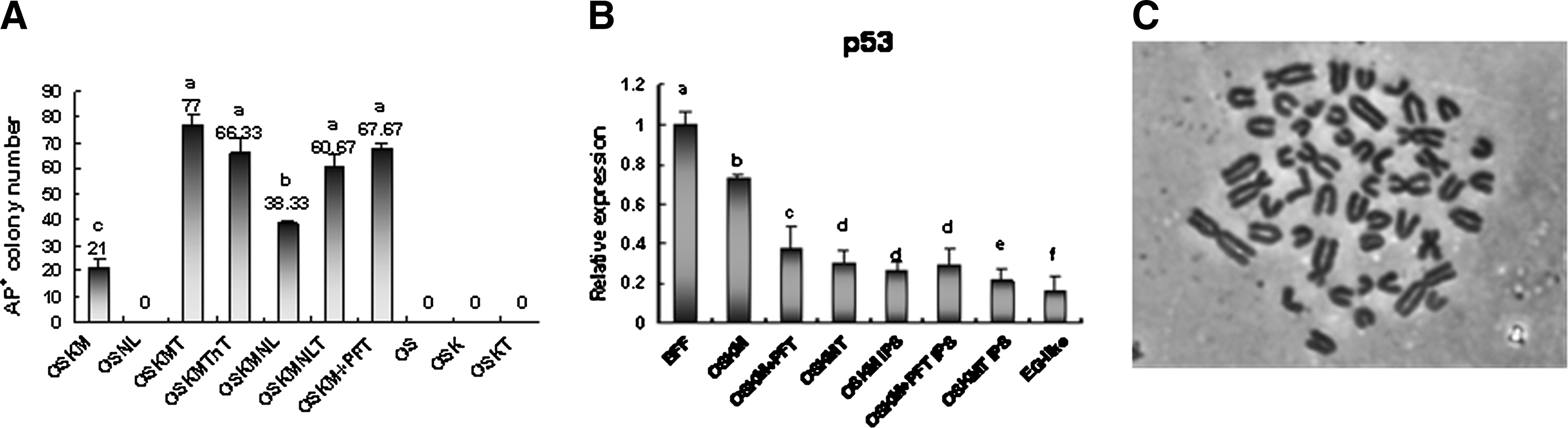
biPSCs generated with different combinations.
Increasing the efficiency of biPSCs by inhibiting p53 expression with SV40 large T antigen
The SV40 large T antigen has been reported to improve the efficiency of iPSC generation [35,36], especially for the establishment of livestock iPSC lines (such as sheep and hircine) [10,11]. Some reports indicate that reducing the expression of p53 can improve the efficiency of iPSC generation [40 –44], and the SV40 large T antigen may increase iPSC generation probably by inhibiting the expression of p53 [10]. To investigate the effect of SV40 large T antigen on buffalo somatic cell reprogramming and biPSC generation, retrovirus vector coexpressing SV40 large T antigen and OSKM were employed to generate biPSCs. When SV40 large T antigen was added to the OSKM, 77 AP positive colonies were generated from 1×104 BFFs, which was around 3-fold higher than OSKM only (Fig. 2A). More importantly, the biPSC colonies could be expanded more than 10 passages and exhibited normal karyotype (Fig. 2C). We named these limited biPSC lines as biPS1, biPS2, biPS3, and biPS4.
To examine the effect of p53 inhibitor, pifithrin-a (PFT) on the biPSC reprogramming, BFFs infected with OSKM were treated with PFT (20 μM) for 1 week after transduction. When BFFs were treated with PFT, 62 AP-positive colonies were generated from 1×104 BFFs, which was around 2.5-fold higher than the untreated group (Supplementary Fig. S1). However, the biPSCs derived from BFFs treated with PFT could not expand more than 6 passages in the PFT supplementing medium, indicating that PFT can promote biPSC reprogramming, but has little effect on long-term proliferation.
QPCR was applied to test the p53 relative expression level during the process of biPSC generation in different combinations. The expression level of p53 in BFFs was higher than biPSC colonies and EG-like cells. On day 6 after transduction with OSKM, OSKM+PFT, and OSKMT, the p53 expression of BFFs decreased, and the decrease extent in BFFs transduced with OSKMT and OSKM+PFT was higher than the BFFs transduced with OSKM (Fig. 2B).
Expression of pluripotent markers and silence of transgenes in biPSCs
The generated biPSC colonies showed round and tightly packed morphology, high ratio of nucleus to cytoplasm, and prominent nucleoli, which were similar to the human ESCs but not mouse and rat (Supplementary Fig. S2). Meanwhile, biPSCs exhibited a high AP activity by AP staining and expressed ESC-related markers (Oct4, Sox2, Nanog, SSEA-1, SSEA-4, TRA-1-81, and E-Cadherin) by immunofluorescence (Fig. 3A). RT-PCR revealed that biPSCs expressed buffalo endogenous pluripotency-related genes, including Oct4, Sox2, Nanog, STAT3, GP130, FOXD3, E-Cadherin, bFGF2, and p53 (Fig. 3B). The expression level of endogenous Oct4, Sox2, and Nanog in biPS1, biPS2, and biPS4 were similar to the ICM and embryonic germ stem cells (EG-like cells) by QPCR (Fig. 3C).
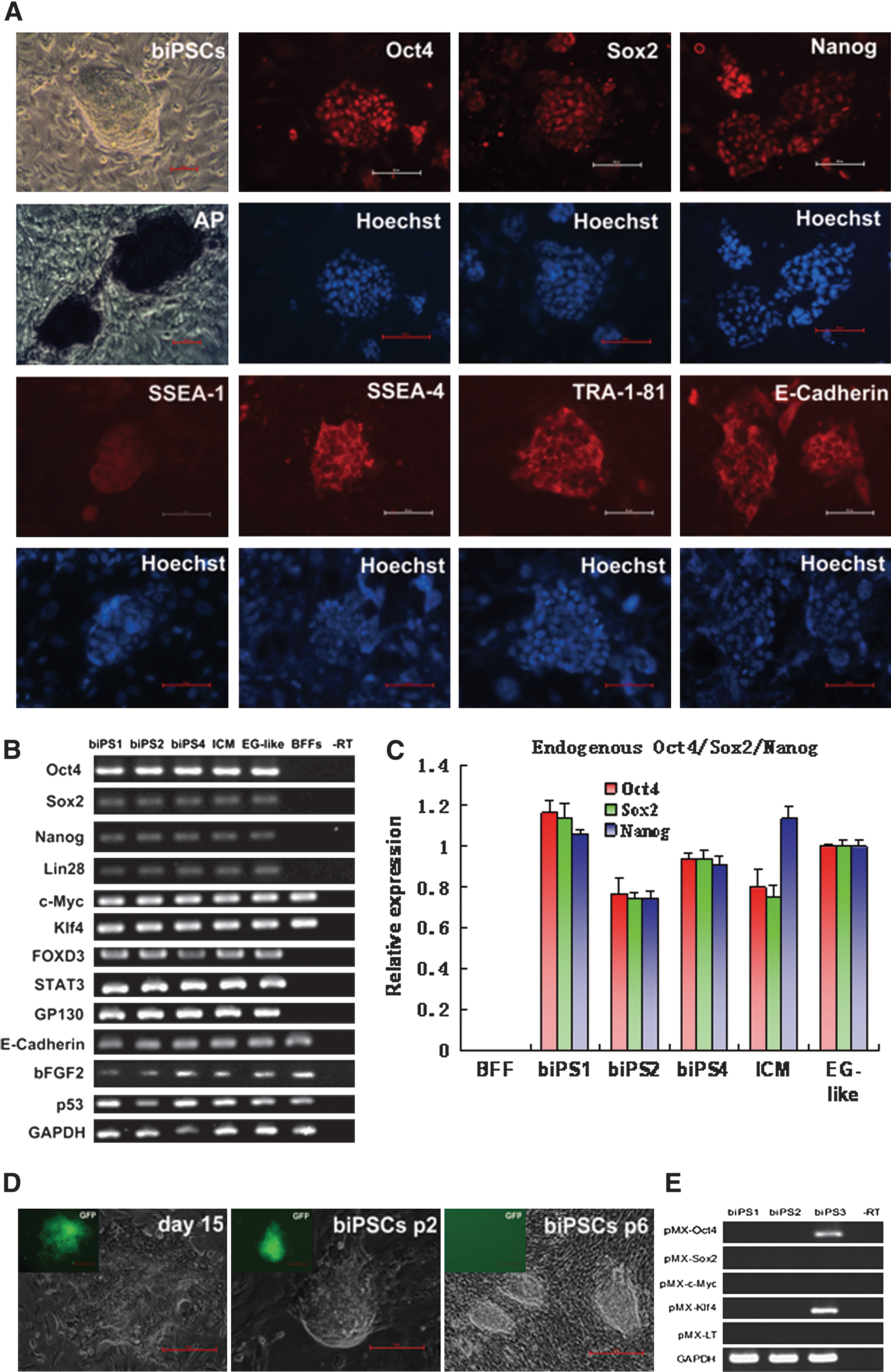
The expression of genes related to pluripotency and silencing of exogenous transgenes in biPSCs.
The expression of GFP was observed in some biPSC colonies in the early formation stage. However, the GFP was silenced after 4–6 passages, indicating that the exogenous genes had been silenced (Fig. 3D). To detect the silencing of exogenous genes precisely, primers specific for retrovirus vector and buffalo defined factors were designed, and no exogenous genes expression was detected in biPS1 and biPS2 after passage 10. But some exogenous transgenes continuously expressed in some colonies, such as Oct4 and Klf4 in biPS3 (Fig. 3E).
Epigenetic status of biPSCs
To detect the relation between expression pattern and the promoter methylation status of defined factors, the CpG islands involved in the fragment of buffalo of Oct4, Nanog, Sox2, and E-Cadherin were cloned and sequenced. The methylation status of those fragments was evaluated using bisulfate genomic sequencing. The promoter region of Oct4 remained hypermethylated both in the parental BFFs and early passage biPSCs (partially reprogrammed biPSCs or pre-biPSCs). The promoter region of Oct4 and Nanog were hypomethylated in passages 8 biPSCs, which was consistent with the activation of endogenous Oct4 and Nanog. In contrast, the methylation status of Sox2 and E-Cadherin promoter region were hypomethylated in BFFs, pre-biPSC, and biPSC colonies (Fig. 4).

The methylation status in the promoter regions of Oct4, Sox2, E-Cadherin, and Nanog by bisulfate sequencing. The open circles indicate unmethylated CpGs and closed circles indicate methylated CpGs.
Differentiation ability of biPSCs in vitro and in vivo
To confirm the differentiation potential of biPSCs, the differentiation ability of biPSCs was examined in vitro and in vivo. EBs were formed in vitro after suspension culturing of biPSCs in the differentiation medium for 7–14 days (Fig. 5A). RT-PCR revealed that biPSCs were able to differentiate into all 3 germ layers, as evidenced by the high expression of AFP, GATA4 (endoderm), ACTA2 (mesoderm), and TUBB3 (ectoderm) marker genes (Fig. 5B). When iPS1 and iPS2 were hypodermically injected into the BALB/C nude mice, typical teratomas were formed in the recipient mice on 6–10 weeks after injection (Fig. 5C). The specific PCR amplification confirmed that these teratomas were from buffalo but not mice (Fig. 5D). No teratomas were observed in 3 negative control nude mice injected with BFFs. The teratomas contained various tissues of the 3 germ layers, including epidermis, neural tissues (ectoderm), muscle, cartilage (mesoderm), gut-like epithelium, and respiratory epithelium (endoderm) as revealed by histological examination (Fig. 5E).

Differentiation of biPSCs in vitro and in vivo.
Discussion
In the present study, buffalo somatic cells were successfully reprogrammed into iPSCs with buffalo original defined factors Oct4, Sox2, Klf4, and c-Myc. The biPSCs displayed similar typical characteristics of the ESCs, such as the specific morphology, AP activity, and expression of pluripotent marker genes, and had the capability to differentiate into 3 germ layers both in vitro and in vivo, and develop to blastocysts after nuclear transfer (Supplementary Fig. S3). These efforts will provide a feasible approach for investigating buffalo stem cell signal pathways, establishing buffalo stem cell lines, and producing genetic modification buffaloes in future.
It is generally accepted that there are some technical keys to reprogram somatic cells into iPSCs successfully for each animal species [9]. For buffalo, the choice of vector skeleton and retrovirus packaging system seem to be critical for biPSC generation. Initially, we attempted to reprogram BFFs with MSCV-based retrovirus system, which was used to generate human iPSCs successfully [45]. However, biPSCs could not be generated with MSCV packaging retrovirus.Then, we tried pMX retrovirus system, which has been proved to be efficient in mouse [1], human [2], and pig [46] iPSC generation. It was found that the buffalo fibroblasts were difficult to be infected by retrovirus. This may be due to the fact that the cell surface receptor of buffalo fibroblasts cannot efficiently bind to the virus envelope protein VSV-G in the infection condition of low titer pseudovirus [47]. Another reason may be due to the low infection efficiency of retrovirus in nondividing cells as observed in other reports [48]. To overcome the above difficulties, we increased the titer of the pseudovirus through ultracentrifugation, and took a twice transduction strategy to infect the BFFs. Around 18 days after transduction, the biPSC colonies appeared. In addition, to delete the negative effect of contact inhibition on the generation of biPSCs, the density of infected cells was examined and plating of infected cells onto MEF feeder at 1×104 cells per 60 mm dish was found to be optimal for generation of biPSCs. This was consistent with the phenomena that the moderate proliferation cells (such as liver and stomach cells) reprogrammed more efficiently than fibroblasts in rabbit [20] and mouse [9,49].
The silencing of exogenous transgenes and activation of endogenous pluripotent factors are hallmarks of full reprogramming of iPSCs. The continuous expression of exogenous factors would inhibit the differentiation of iPSCs [8]. Ideally, the exogenous genes should not integrate into the genome of the target cells or be excised once reprogramming has been completed. As reported in mouse, the expression of endogenous Nanog [50] and E-Cadherin [38,51,52] was necessary for full reprogramming of iPSCs. In the present study, the exogenous transcripts were silenced when the biPSCs were expanded for more than 6 passages in some colonies, and the expression of endogenous pluripotent factors were activated, including Nanog and E-Cadherin. Moreover, demethylation of the promoter in the pluripotency related genes was required for full reprogramming of iPSCs [53]. In mice, the reprogramming process was multiple-course with methylation status changes at the pluripotency-related loci, which were hypermethylated in somatic cells and partially reprogrammed cells (pre-iPSCs), contrary to the fully reprogrammed cells in which the pluripotency-related loci were hypomethylated [43,54]. In the present study, the promoter region of Oct4 and Nanog were hypomethylated in biPSCs, whereas hypermethylated in BFFs and pre-biPSCs. However, the methylation status of Sox2 and E-Cadherin promoter exhibited no difference among BFFs, pre-biPSCs, and biPSCs. These results were consistent with the mouse iPSCs [52,54]. Thus, hypomethylation of the promoter of endogenous pluripotent genes is necessary for full reprogramming of biPSCs.
Tumor suppressor p53 is not only involved in tumor pathway, but also in reprogramming of iPSCs and regulation of the mouse ESC and human ESC differentiation [55,56]. Inhibition of p53 expression could inhibit p53-mediated apoptosis, and improve the efficiency of somatic cell reprogramming [40]. The SV40 large T antigen can bind to p53 and inhibit its function [34]. It had been reported that SV40 large T antigen combined with other iPSCs-related factors could improve the reprogramming of human iPSCs [36]. Thus, some researchers speculated that SV40 large T antigen may regulate certain signaling pathways through inhibiting p53, such as the bFGF-PI3K-AKT pathway [57], or remodel the chromatin, which are beneficial for the reprogramming of iPSCs [10]. In the present study, SV40 large T antigen was found to decrease the expression of p53 and increase the efficiency of biPSC reprogramming. Alternatively, the employment of p53 inhibitor PFT to inhibit the expression of p53 in the early stage of reprogramming could promote the reprogramming of biPSCs, but did not increase the ability of biPSCs in self-renewal and proliferation. Thus, the action of SV40 large T antigen in promoting the biPSC reprogramming and expansion of passages may be due to its other roles besides the p53 pathway, such as immortalization.
In this study, we also found that these defined factors related to iPSCs indicate high functional conservation between buffalo and other different species. It has been reported that mouse and human defined transcription factors can reprogram the somatic cells of pig [7,8,19,46], sheep [10,58], and equine [12] into iPSCs. In fact, the buffalo factors could reprogram mouse fetal fibroblasts into iPSCs (data not shown). Considering the problem of heterologous gene contamination in the future genetic improvement, buffalo-derived defined factors were employed to generation biPSCs in the present study.
One important standard to judge the animal pluripotent stem cells is whether the cells have the capability to differentiate into defined cell types in vitro or in vivo. The pluripotency of biPSCs obtained in the present study was demonstrated by EB and teratoma assays, indicating that their reprogramming was completed. However, the biPSCs obtained were only maintained for around 10 passages. The limited passage of biPSCs obtained may be due to the suboptimal culture condition employed in the present study. In fact, most of buffalo ESC-like cells derived from blastocysts produced by IVF did not survive beyond 10 passages [31,32,59]. Recently, Sharma et al. [60] reported that buffalo ESC could be maintained for more than 85 passages under the culture condition of Knockout DMEM™, 15% Knockout serum replacement™, 5 ng/mL FGF-2, and 1,000 IU/mL mLIF. Thus, more works should be done to optimize culture condition for maintaining the self-renewal and proliferation of biPSCs in vitro for a long time in the future.
Footnotes
Acknowledgments
We are grateful to Dr Xibang Zheng (Guangxi University) for gifting vectors and Dr Shaorong Gao (National Institute of Biological Sciences, Beijing) for his technical assistance. This work was supported by the China High Technology Development Program (2011AA100607), China Transgenic Project (2011ZX08007-003), China Natural Science Foundation (30960251), and Guangxi Science Foundation (2010GXNSFF013002).
Author Disclosure Statement
No potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.