
Abstract
Friedreich's ataxia (FA) is a multisystemic disorder characterized by progressive gait, ataxia, and cardiomyopathy. There are few treatments for this disease; thus, we analyzed in vitro the possible beneficial effect of adult stem cells in FA. To this end, human adipose stem cells from healthy individuals and periodontal ligament cells from FA patients were isolated and cultured. FA cells are especially vulnerable to oxidative stress; thus, they were submitted to this condition and cultured in adipose stem cell–conditioned medium. This resulted in increased cell survival and upregulation of oxidative-stress-related genes as well as frataxin, among other genes. A number of trophic factors were shown to be expressed by the adipose stem cells, especially brain-derived neurotrophic factor (BDNF), which was also identified in the conditioned medium. The culture of the ataxic cells under oxidative stress and in the presence of this trophic factor confirmed its protective effect. Thus, this work demonstrates that adipose stem cell–conditioned medium from healthy individuals is capable of changing the transcription levels of oxidative-stress-related genes in cells that are particularly susceptible to this condition, avoiding cellular degeneration. Also, this work shows how neurotrophic factors, particularly BDNF, are capable of increasing cell survival in response to oxidative stress, which occurs in many neurodegenerative diseases.
Highlights
1. Periodontal ligament cells from Friedreich's ataxia (FA) patients are susceptible to oxidative stress.
2. Adipose-tissue-derived stem cell–conditioned medium reduces active caspase-3 levels and increases cell survival.
3. Adipose stem cell–conditioned medium contains a number of trophic factors, brain-derived neurotrophic factor (BDNF) being the most significant.
4. BDNF is a vital trophic factor to protect FA cells from oxidative stress.
Introduction
A
Stem cells have been proven to be a potent tool for the treatment of many diseases, including neurodegenerative disorders. It has been shown in various animal studies that stem cells are capable of stopping degenerative processes or even regenerate lost tissue. This occurs due to several mechanisms: cell fusion [5 –8], trophic factor release [9 –11], immunomodulation [12], transdifferentiation [13,14], or activation of host progenitor cells [15,16].
In our lab, we have previously shown that bone marrow stem cells are capable of protecting degenerating neurons in demyelinating, motorneuron degenerating, and cerebellar ataxia animal models [14,17–19, respectively]. Previous works have proven that adipose stem cells have similar effects as bone marrow [20]. Thus, we decided to investigate the use of adult stem cells in a FA model. Specifically, we will analyze the possible neuroprotective effect stem cells may have on degenerating ataxic cells. To this end, we isolated and expanded periodontal ligament cells from FA patients, in order to study in vitro the beneficial effects adult stem cells may exert. Since FA cells are sensitive to oxidative stress, we submitted the cells to this condition and cocultured the cells in adipose stem cell–derived conditioned medium. The results of this work show how stem-cell-mediated trophic factor release may increase cell survival under oxidative stress conditions.
Experimental Procedures
Adipose-tissue-derived stem cell isolation and culture
Human adipose tissue was obtained by liposuction from healthy individuals, who previously signed a written consent, and immediately refrigerated. No more than 12 h after the extraction, the adipose tissue was processed. First, the tissue was washed extensively in phosphate-buffered saline (PBS; Invitrogen) supplemented with 1% penicillin/streptomycin (Invitrogen). Afterward, it was digested in 0.2 mg/mL animal-component-free collagenase-I (Worthington) for 45 min at 37°C, followed by mechanical dissociation. Then, the tissue was washed extensively to remove the collagenase and centrifuged at 1,500 rpm for 10 min. The resulting pellet was washed and centrifuged twice, and counted in a Neubauer chamber before placing in culture. The culture medium used was animal-origin-free mesenchymal stem cell medium (StemCells Technologies), and the cells were placed in a cell culture flask at a concentration of 1×106 cells/flask. The medium was changed 48 h after the initial plating and afterward every 3–4 days. After 7–10 days, when the culture was almost confluent, the cells were detached using TrypLE Select animal-origin-free stable trypsin replacement (Gibco) and replated at a concentration of 50,000 cells/flask. For the experiments, cells from passages 2–3 were used, which considered to be adipose-derived stem cells (Fig. 1A). To confirm this, immunocytochemical staining for CD34, CD44, CD90, and CD45 was performed (see Fig. 1A–D).

Characterization and effect of oxidative stress in adipose-derived stem cells of healthy individuals.
Periodontal-ligament-derived stem cell isolation and culture
Before extraction, the patients were informed of the procedures to be performed and signed a written consent. The teeth used were deciduous teeth in the process of falling out. The teeth, either after spontaneously falling or by extraction in a dental clinic, were immediately taken into the laboratory and processed. After extracting the periodontal ligament, the tissue was processed in a similar fashion as the adipose tissue (collagenase treatment, followed by mechanical dissociation and culture). The cells used were at passages 3–5. In the case where periodontal ligament cells from healthy individuals were used, this was taken from the biobank of our institution. This cell population is a mixture of different individuals so as to maintain patient's privacy. The procedure to extract the cells was the same; only in this case the teeth were taken from individuals of various ages.
Immunocytochemical analysis
A standard immunocytochemical protocol was used. First, the cells were fixed in 4% paraformaldehyde, washed, and blocked with 10% goat serum, 5% bovine albumin, 0.025% triton, and PBS. Afterward, they were incubated overnight at room temperature with the primary antibody, diluted in blocking solution (10% goat serum, 5% bovine albumin, and PBS). The following primary antibodies were used: mouse anti-CD44 (1:200; BD Pharmingen), mouse anti-CD45 (1:200; AbD Serotec), mouse anti-CD34 (1:200; BD Pharmingen), mouse anti-CD90 (1:200; BD Pharmingen), and rabbit anti-cleaved caspase-3 (1:400; Cell Signaling Technology). The following day, the cells were incubated with the secondary antibodies, which were either Alexa Fluor antibodies (1:500; Molecular Probes) or biotinylated secondary antibodies (1:200; Vector Laboratories) followed by an incubation with streptavidin conjugated with Cy3 (1:500). DAPI (Molecular Probes) or Hoechst (Sigma-Aldrich) was used to stain nuclei. Histological samples were observed and micrographs were taken under a fluorescence microscope (Leica DMR; Leica Microsystems).
Hydrogen peroxide, brain-derived neurotrophic factor, and TrkB blocking treatment of cultured cells
Adipose stem cells from healthy individuals and periodontal ligament cells from healthy and FA patients were cultured in standard medium and supplemented with 0.1 mM hydrogen peroxide (Sigma-Aldrich) for 48 h. Hydrogen peroxide was added twice, once at the beginning of the experiment and 24 h afterward. To obtain the adipose stem cell–conditioned media, this was recovered 24 h after incubation under normal culture conditions. Also, conditioned medium from adipose stem cells exposed to hydrogen peroxide for 24 h was recovered. Both conditioned media were centrifuged at 1,500 rpm for 10 min and the resulting pellet was discarded. Also, periodontal ligament cells were cultured in the same hydrogen peroxide medium [labeled L(H) in the Figs. 3 and 4]. Depending on the stem-cell-conditioned media, 2 different experimental conditions were performed: periodontal ligament cells cultured in 0.1 mM hydrogen peroxide and in adipose stem cell–conditioned medium [L(H)+A in the Figs. 3 and 4], and periodontal ligament cells exposed to hydrogen peroxide and in conditioned medium from adipose stem cells that had previously been submitted to oxidative stress [L+A(H)].
For the cultures where brain-derived neurotrophic factor (BDNF) was added, this trophic factor was included along with the hydrogen peroxide for 2 days, at several concentrations (1–100 ng/mL; Sigma Aldrich), while in the cell cultures with the TrkB blocker (recombinant human TrkB Fc Chimera; R&D Systems), 100 ng/mL was used.
Real-time quantitative PCR
Total mRNA of the cells was isolated using the Trizol protocol (Invitrogen). Five micrograms of mRNA was reverse-transcribed, and ∼100 ng of cDNA was amplified by real-time PCR using Power SYBR Green Master mix (Applied Biosystems). All the samples were run in triplicate using the StepOne Plus Real-Time PCR system (Applied Biosystems) and analyzed with the StepOne Software. Analyses were carried out using the delta C(T) method and calculated relative to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (forward: AGCCACATCGCTCAGACAC, reverse: GCCCAATACGACCAAATCC). The following primers were used, taken from the PrimerBank webpage (
Western blot analysis
The lysis buffer and process used were similar to that of Palomo et al. [21]. Cell lysates and culture media were separated on 15% sodium dodecyl sulfate–polyacrylamide gels, and probed for mouse anti-frataxin (1:750; Santa Cruz Biotechnology) and rabbit anti-BDNF (1:500; Santa Cruz Biotechnology), respectively. Secondary antibodies were visualized by chemiluminescence (ECL; Amersham). For protein quantification we used Quantity One software (BioRad).
Statistical analysis
Statistical significance between control and experimental groups was calculated with Sigmaplot v11.0 software, using the paired t-test.
Results
Adipose stem cells exposed to hydrogen peroxide increase the expression levels of genes implicated in oxidative stress and iron metabolism
Adipose-tissue-derived stem cells were isolated, cultured, and characterized as seen in Figure 1. These cells, as in bone marrow mesenchymal stem cells, express the surface markers CD44 and CD90, and do not express CD34 and CD45 (Fig. 1A–D). Also, periodontal ligament cells from FA patients were isolated and cultured. These cells express several stem cell and neural markers, due to their neural crest origin [22].
The effect of hydrogen peroxide on the expression of several genes in adipose-tissue-derived stem cells of healthy individuals was analyzed (Fig. 1E). The genes studied were superoxide dismutases (SOD1, SOD2, and SOD3), hydrogen peroxide neutralizing genes (Catalase and GPX-1), NADPH oxidases (NOX1, NOX2, NOX3, NOX4, and NOX5), dual oxidases (DUOX1 and DUOX2), as well as genes implicated in iron metabolism (frataxin, transferrin, H-ferritin, and L-ferritin). As it can be seen in the histogram, the adipose stem cells exposed to hydrogen peroxide expressed increased transcription levels of several oxidative-stress-related genes, including SOD1, catalase, and several of the NOX and DUOX2. Also, almost all the iron-related genes, except for L-ferritin, were upregulated. As the cause for the degeneration in FA is due to low levels of frataxin and subsequent iron accumulation, the possible upregulation of these genes in FA patients is of utmost importance. Overall, these results indicate a possible compensatory response of the cells in order to increase cell survival due to the oxidative stress conditions, increasing overall the metabolism of the cell.
Comparative gene expression between periodontal ligament cells isolated from healthy and FA individuals
Periodontal ligament cells were extracted from both healthy and FA individuals, and the basal expression levels of the genes commented in the previous section were analyzed under normal culture conditions (Fig. 2A). As a result, FA cells expressed significantly lower levels of frataxin (see Fig. 2B for western blot analysis), as well as the majority of the genes analyzed. On the other hand, 2 genes, NOX3 and DUOX1, presented higher levels of expression. Previous studies in yeast and human cells have shown that antioxidant enzymes are affected due to the frataxin defect, and are not capable of responding to oxidative stress conditions, corroborating our results [23,24, respectively].
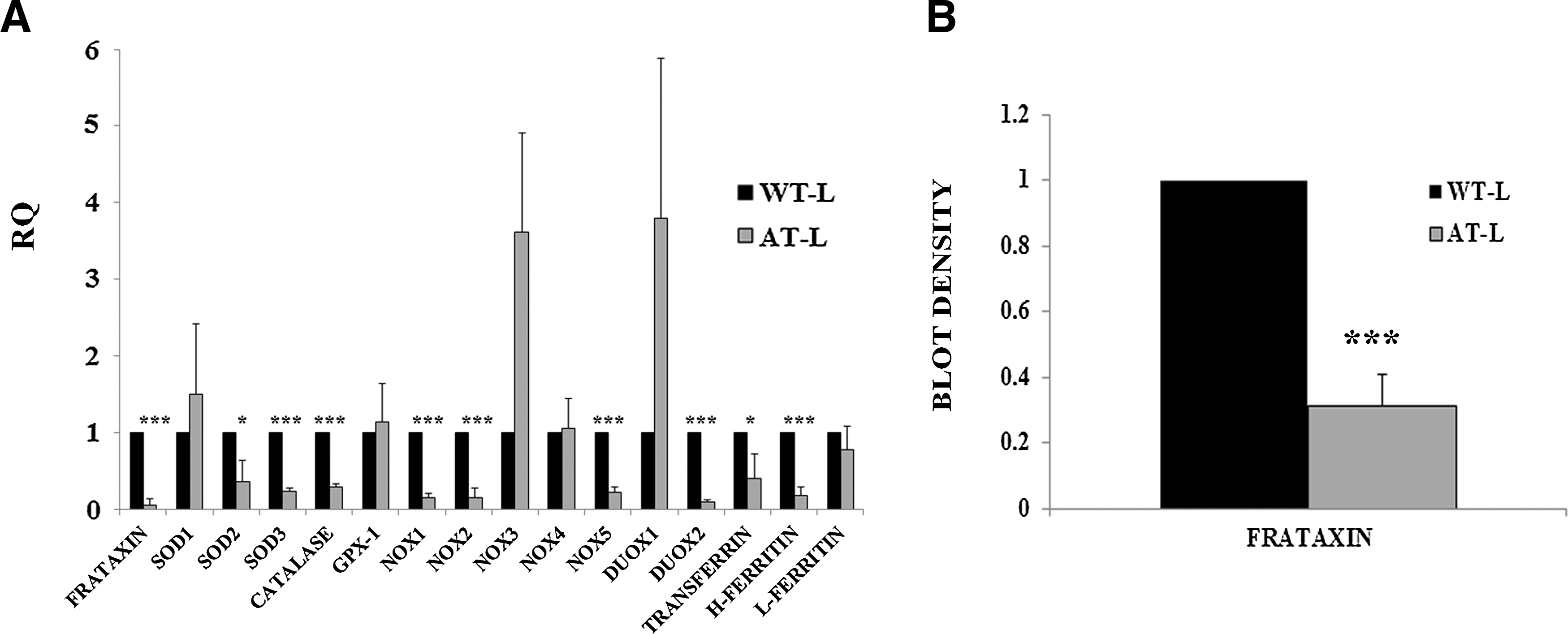
Comparative analysis of periodontal ligament cells isolated from healthy and Friedreich's ataxia (FA) patients under normal culture conditions.
Adipose-tissue-derived stem cell–conditioned medium increases cell survival and upregulates the expression levels of genes implicated in oxidative stress and iron metabolism, in cultured cells isolated from FA patients
The periodontal ligament cells from FA patients were submitted to oxidative stress, as in the case of the adipose stem cells. Cell death was calculated both using Hoechst (Fig. 3A–D) and active caspase-3 staining (Fig. 3E–H). The following culture conditions were performed: periodontal ligament cells under normal culture conditions (Fig. 3A), periodontal ligament cells exposed to hydrogen peroxide (Fig. 3B), periodontal ligament cells exposed to hydrogen peroxide and in adipose stem cell–conditioned medium (Fig. 3C), and periodontal ligament cells exposed to hydrogen peroxide and in conditioned medium from adipose stem cells previously submitted to oxidative stress (Fig. 3D). In the case of Hoechst staining, 20.71%±4.56% of periodontal ligament cells exposed to hydrogen peroxide presented pyknotic nuclei indicating that the cells were undergoing cell death processes (Fig. 3E), as opposed to 1.76%±0.74% and 0.44%±0.58% of the periodontal ligament cells cultured under 1 of the 2 adipose stem cell–conditioned medium (Fig. 3F, G; n=3). As for active caspase-3 staining, 9.55%±2.55% of the periodontal ligament cells exposed to hydrogen peroxide expressed active caspase-3 (Fig. 3E). Meanwhile, in the cultures with adipose stem cell–conditioned medium (Fig. 3F–G), only 1.40%±0.38% (in the case of FA periodontal ligament cells under stem-cell-conditioned medium) and 1.69%±0.25% (in the case of FA cells under conditioned medium from stem cells that have been previously submitted to oxidative stress) were active caspase-3 positive (n=3).
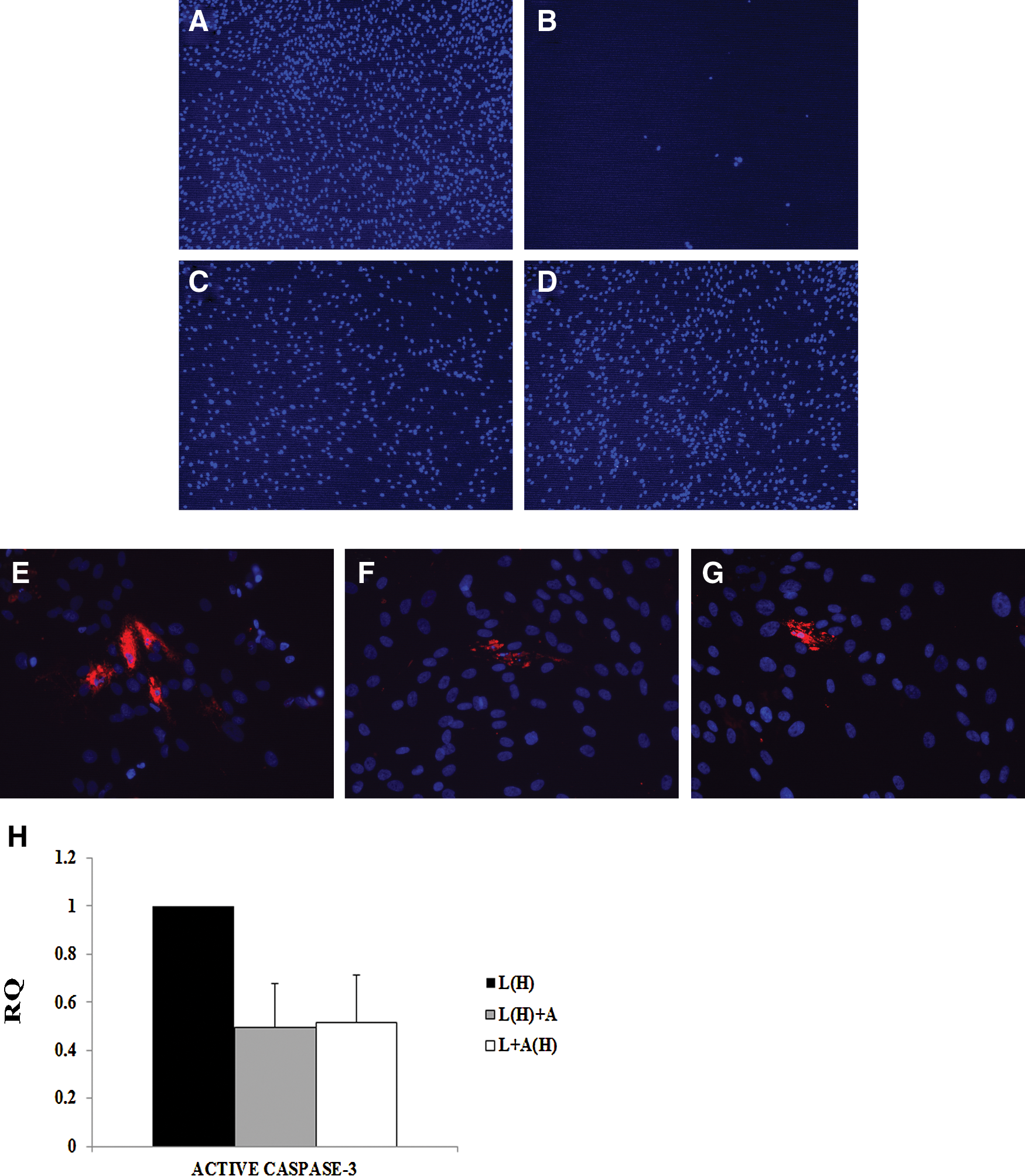
Caspase-3 is activated in response to oxidative stress.
Active caspase-3 was also analyzed by quantitative PCR (Fig. 3H), corroborating the immunocytochemistry results, indicating that stem-cell-conditioned medium increases the cell survival of FA cells exposed to hydrogen peroxide. In the Fig. 3H, L(H)+A indicates periodontal ligament cells exposed to hydrogen peroxide and in adipose stem cell–conditioned medium, whereas L+A(H) is periodontal ligament cells exposed to hydrogen peroxide and under conditioned medium from adipose stem cells previously exposed to oxidative stress. Also, L(H) indicates periodontal ligament cells exposed to hydrogen peroxide.
Also, quantitative expression of oxidative stress and iron-metabolism-related genes were analyzed and compared in periodontal ligament cells under normal culture conditions and in the presence of hydrogen peroxide (Fig. 4A). As a result, it was observed that the expression levels of the genes analyzed in periodontal ligament cells from FA patients under oxidative stress either did not change or were even decreased. Only NOX1 and NOX2 were upregulated. This was opposed to what was observed in the adipose stem cells from healthy individuals (see Fig. 1E, almost all the genes were upregulated). However, when the periodontal ligament cells exposed to hydrogen peroxide were cultured under stem-cell-conditioned medium, there was an overall upregulation of the genes analyzed (Fig. 4B). Of the 2 adipose stem cell–conditioned media used, the one taken from stem cells that had previously been exposed to hydrogen peroxide [L+A(H) in the Fig. 4C] presented the highest amount of frataxin upregulation, increasing its expression ∼4 times. Also, SOD2, GPX-1, NOX5, DUOX2, transferrin, H-ferritin, and L-ferritin were increased. Increased frataxin expression at the protein level was corroborated by western blot analysis (Fig. 4C, D).
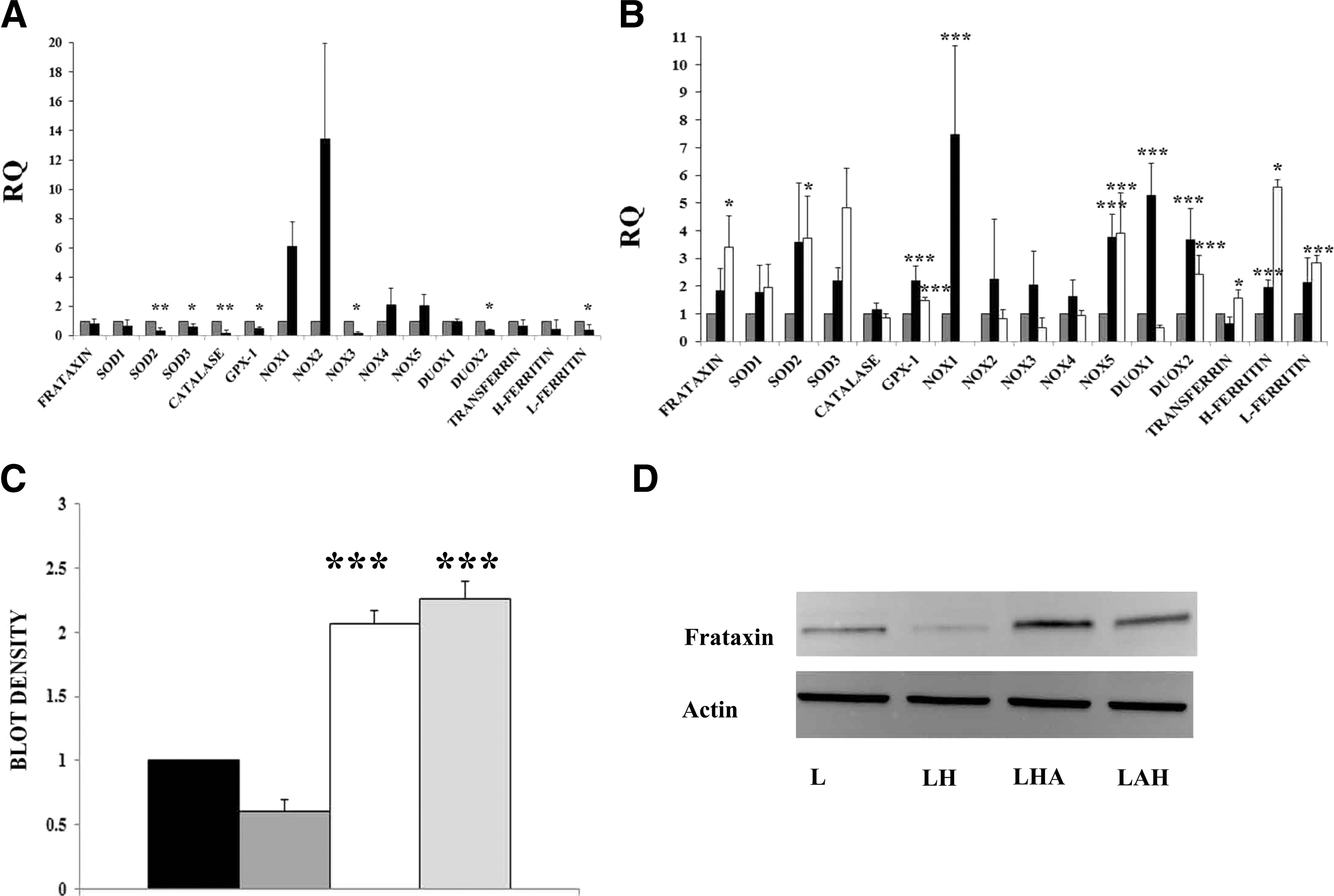
Gene and protein expression in response to oxidative stress in FA cells cultured with adipose-tissue-derived stem cell–conditioned medium.
Adipose-tissue-derived stem cells secrete several trophic factors in response to hydrogen peroxide
As the adipose stem cell–conditioned medium, especially from cells that had previously been under oxidative stress conditions, was sufficient to increase oxidative-stress-related genes as well as frataxin, we decided to analyze the conditioned medium for trophic factors that may induce this upregulation. A total of 10 trophic factors were analyzed by quantitative PCR in the adipose stem cells (Fig. 5A). The result was that there was a significant increase in BDNF, VEGF, and NGF when comparing adipose stem cell–conditioned medium exposed to hydrogen peroxide [A(H) in the figure] compared with standard culture conditions (A in the Fig. 5A). Specifically, BDNF was the trophic factor that increased the most, with a 5-fold increase, compared with VEGF and NGF, which presented a more modest increase (<2-fold).
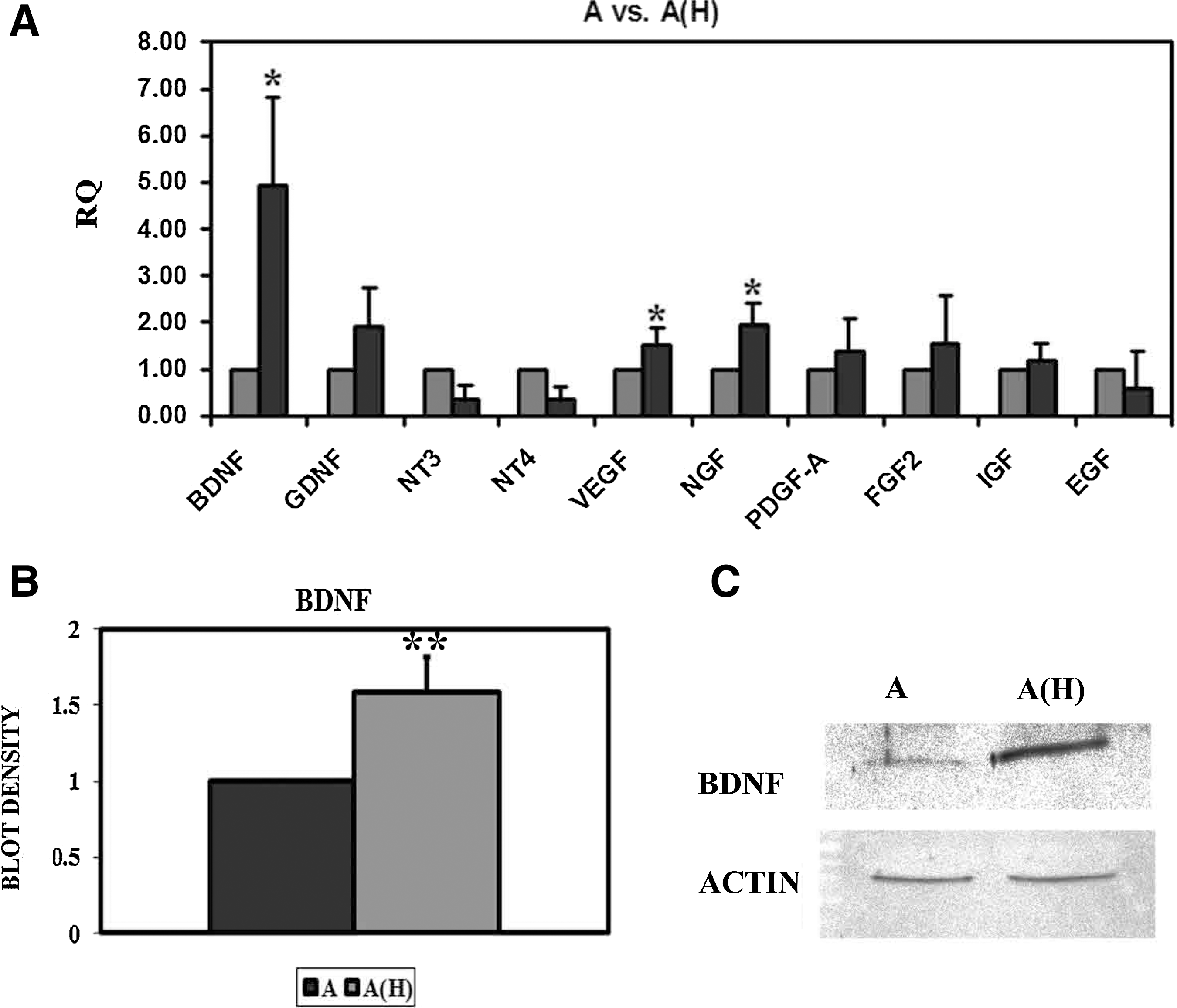
Trophic factors released by adipose-tissue-derived stem cells.
Further, western blot analysis demonstrated that BDNF was secreted in higher quantities in the conditioned medium from stem cells under oxidative stress (Fig. 5B, C). Interestingly, the strongest band to appear in the gel was at 28 kDa, corresponding either to the truncated BDNF form [25] or BDNF-BDNF dimer [26]. Either way, both forms are biologically functional and are capable of binding to the BDNF receptors.
As periodontal ligament cells express surface receptors where BDNF may bind to [27,28], it is plausible to consider that this trophic factor may be an important factor responsible for the increase in cell survival observed, as well as the upregulation of the genes analyzed.
BDNF increases the expression of SODs, NOXs, frataxin, and iron-metabolism-related molecules in response to hydrogen peroxide
To confirm whether BDNF alone is capable of protecting FA periodontal ligament cells from oxidative stress, we cocultured for 2 days the cells in different concentrations of the trophic factor (1–100 ng/mL). There was a dose-dependent response to BDNF, being the most significant improvement when the trophic factor was used at 100 ng/mL. Active caspase-3 was analyzed by quantitative PCR, resulting in less expression of this gene in cells cultured with BDNF (Fig. 6A). As in the other culture conditions, oxidative stress and iron-metabolism-related genes were analyzed, several of these being significantly upregulated. These results were similar to those seen with the adipose stem cell–conditioned medium (see Fig. 4B). Thus, BDNF seems to be capable of protecting FA cells from oxidative stress and may be at least partially responsible for the increase in cell survival, as well as upregulation of the genes analyzed.

Gene expression analysis of periodontal ligament cells cultured in 100 ng/mL BDNF, and with recombinant TrkB antibody.
To confirm the protective effect of BDNF under oxidative stress, the receptor for BDNF, TrkB, was blocked using a recombinant human antibody. Periodontal ligament cells cultured with this antibody were not affected (Fig. 6C). However, when the periodontal ligament cells were cultured with the antibody and exposed to hydrogen peroxide, there was massive cell death, even under stem-cell-conditioned medium (Fig. 6D–F, respectively). This was clearly visible in the cultures, where numerous cells died and detached from the culture dishes. Active caspase-3 and Hoechst staining was performed and quantified in the various cultures. As a result, 2.75%±0.47% of the periodontal ligament cells under normal cell culture and with the TrkB blocker presented pyknotic nuclei, and 2.60%±1.48% were active caspase-3 positive. In the other culture conditions, not only was there a very significant loss of cell density, but also significantly higher percentages of pyknotic nuclei (7.73%±1.16% in the case of periodontal ligament cells exposed to hydrogen peroxide, 10.32%±4.23% in the case of periodontal ligament cells exposed to hydrogen peroxide and under stem-cell-conditioned medium, and 10.31%±1.87% in the cells exposed to hydrogen peroxide and under conditioned medium from adipose stem cells that have previously been submitted to oxidative stress, n=3). This was corroborated by active caspase-3 immunostaining (2.99%±0.53% of the periodontal ligament cells exposed to hydrogen peroxide, 1.35%±0.90% of the cells under stem-cell-conditioned medium, and 4.65%±3.10% of the cells under conditioned medium from stem cells that had previously been submitted to oxidative stress, n=3).
Active caspase-3 gene expression was also analyzed by quantitative PCR (Fig. 6G, left histogram). As a result, all treatments presented higher active caspase-3 expression compared with control cultures, coinciding with the immunohistochemical analysis. Further, the gene expression analysis by quantitative PCR indicated no significant difference in the expression of frataxin, SOD1, SOD2, catalase, NOX3, DUOX2, or L-ferritin. However, the expression levels of the other genes (SOD3, GPX-1, NOX2, NOX4, NOX5, DUOX1, transferrin, and H-ferritin) were increased in the periodontal ligament cells exposed to hydrogen peroxide and under conditioned medium of stem cells submitted to oxidative stress, despite the TrkB blocker. Thus, blocking the BDNF receptor decreased the effectiveness of the adipose stem cell–conditioned medium, increasing cell death, although other factors in the conditioned medium were capable of upregulating oxidative-stress-related genes.
Discussion
The work presented here shows for the first time that adipose-tissue-derived stem cells are capable of protecting FA cells submitted to oxidative stress. This is due to the secretion of trophic factors in response to the damage, mainly BDNF. In response to this trophic factor, the ataxic cells increase frataxin expression ∼4 times, while increasing the expression of many genes implicated in hydrogen peroxide removal, as well as other factors implicated in iron metabolism (ferritins and transferrin). Recently, an article has been published showing that bone marrow mesenchymal stem cell–conditioned medium from healthy patients had similar effects on fibroblasts isolated from FA patients as those observed in this work [29]. Our work elucidates on the possible mechanisms by which the conditioned medium induces the increase in cell survival as well as frataxin upregulation. Particularly, it seems that the release of certain neurotrophic factors seems to be the main cause for the observed effect.
It has been previously shown by other authors that adipose stem cells, similar to bone marrow cells, are capable of expressing and secreting many trophic factors, including BDNF [30] and NGF [31]. On the other hand, periodontal ligament cells present surface receptors where BDNF may bind to, such as LNGFR (low-affinity NGF receptor, also known as p75) [27] and TrkB [28]. This trophic factor is known to be implicated in processes of cell survival, differentiation, and proliferation, among other functions [32,33]. Further, it has been shown that inoculating BDNF transgene into an ataxic mouse model improved the behavior test in ataxia cells, corroborating that this trophic factor is an important factor for neuroprotection in these disorders [34]. In our lab, we saw that bone marrow mesenchymal stem cells are capable of protecting Purkinje cells in a cerebellar ataxia model by expressing BDNF and to a lesser degree NT3 and GDNF, resulting in improved behavior results [19].
Interestingly, our western blot analysis of the conditioned media showed that the most abundant form of BDNF being secreted is a 28 kDa isoform. The mature form of BDNF is 14 kDa, while the pro-BDNF (in the endoplasmic reticulum) weighs 32 kDa. The 28 kDa may either be a BDNF dimer, which is also secreted and is fully functional [26], or it may be a truncated, immature form of BDNF, which is also biologically active [25]. Either way, this isoform is completely capable of binding to its ligands, mainly TrkB, thus increasing cell survival.
With these results, it would indicate that BDNF administration could help protect the neurons and cardiomyocytes in FA patients. Indeed, BDNF administration was initially performed in ALS patients, but failed since the trophic factor is not capable of surpassing the blood-brain barrier without a proper vehicle [35]. Intrathecal administration of BDNF, on the other hand, seemed a feasible option, but the clinical trials showed no improvement with respect to placebo controls [36 –39]. Also, it must be noted that BDNF administration presents a number of secondary effects that may debilitate the patient, such as hypersensitivity to pain [40]. Thus, although it seems that the use of this trophic factor alone may help ameliorate the progression of the disease, previous results in other diseases, such as ALS, indicate the contrary. On the other hand, stem cell transplantation may act as a biological pump that administers not only BDNF but also other trophic factors in the correct dosage without the side effects commented previously.
Thus, our work demonstrates that adipose-tissue-derived stem cells of healthy individuals are capable of protecting FA cells from oxidative stress and induce the expression of genes implicated in iron metabolism, including and most importantly frataxin.
Footnotes
Acknowledgments
The authors appreciate the help of F. Rodriguez, M. Ródenas, O. Bahamonde, A. Estirado, and A. Torregrosa for their technical assistance. This work has been financed by EUCOMMTOOLS, Science and Innovation Ministry (MICINN BFU-2008-00588, CONSOLIDER CSD2007-00023), Valencian government (PROMETEO/2009/028), Cell Therapy Network-Carlos III Health Institute (RD06/0010/0023), Alicia Koplowitz Foundation, Granada Foundation of Friedreich's Ataxia, 5P- Syndrome Foundation, and Diógenes Foundation/Elche (CATEDRA ELA).
Author Disclosure Statement
The authors state that there is no potential conflict of interest.