
Abstract
Heart failure is one of the leading causes of death worldwide. Myocardial cell transplantation emerges as a novel therapeutic strategy for heart failure, but this approach has been hampered by severe shortage of human cardiomyocytes. We have recently induced mouse embryonic stem cells to differentiate into embryoid bodies and eventually, cardiomyocytes. Here, we address recent advancements in cardiomyocyte differentiation from cardiac stem cells and pluripotent stem cells. We highlight the methodologies, using growth factors, endoderm-like cell cocultures, small molecules, and biomaterials, in directing the differentiation of pluripotent stem cells into cardiomyocytes. The characterization and identification of pluripotent stem cell-derived cardiomyocytes by morphological, phenotypic, and functional features are also discussed. Notably, increasing evidence demonstrates that cardiomyocytes may be generated from the stem cells of several tissues outside the cardiovascular system, including skeletal muscles, bone marrow, testes, placenta, amniotic fluid, and adipose tissues. We further address the potential applications of cardiomyocytes derived from various kinds of stem cells. The differentiation of stem cells into functional cardiomyocytes, especially from an extra-cardiac stem cell source, would circumvent the scarcity of heart donors and human cardiomyocytes, and, most importantly, it would offer an ideal and promising cardiomyocyte source for cell therapy and tissue engineering in treating heart failure.
Introduction
H
Currently, there are 2 major stem cell sources for generating CMs: cardiac stem cells (CSCs) and embryonic stem (ES) cells. Notably, increasing evidence has demonstrated that CMs may be generated from the stem cells of several tissues outside the cardiovascular system, including skeletal muscle, bone marrow, placenta, testes, amniotic fluid, and adipose tissues. We illustrate various kinds of stem cells for generating CMs for the treatment of heart diseases (Fig. 1). A number of factors have been demonstrated that promote the differentiation of stem cells and accelerate the integration of stem cells after transplantation. Biomaterials can be used to facilitate the delivery of proteins, genes, or small interfering RNA to stem cells [4]. In this article, we address the advancements in the generation of CMs from various kinds of stem cells, including cardiac and extra-cardiac tissue sources. The characterization, identification, and functional analyses of pluripotent stem cell-derived CMs are also discussed. In addition, we discuss the different strategies, including growth factors, endoderm-like cell cocultures, small molecules, and biomaterials, that are used in stem cell-derived CMs and highlight their potential applications in treating cardiovascular diseases.

Stem cell-derived CMs for potential treatment of heart diseases. CM, cardiomyocyte; ES cells, embryonic stem cells; iPS cells, induced pluripotent stem cells; MSCs, mesenchymal stem cells; SP, side population. Color images available online at
Differentiation of CSCs into Mature CMs
Mammalian cardiac development begins with the generation of the mesoderm early in gastrulation. The posterior part of the primitive streak contains CSCs that express both mesoderm posterior-1 and Flk1. The pericardial mesoderm cell has multipotency, as evidenced by the fact that it can produce the myocardial or epicardial lineage [5]. Subsequent development of the heart is determined by complex interactions between genetic programs and mechanical forces. Both CM and cardiac muscle cell differentiation count on the same set of transcription factors. Genetic programs for heart development are controlled by transcription factors that regulate the morphology, phenotypes, and fate decisions of cardiac cells [6]. However, the downstream target genes of these transcription factors that control the fate determinations of cardiac cells remain largely unknown about.
There are several types of CSCs and cardiovascular progenitor cells (CPCs) based on their phenotypes, and we compare the phenotypic and functional traits of various endogenous CSCs and CPCs (Table 1). CSCs expressing c-kit are first identified in the rat heart [7], and, importantly, c-kit-positive CPCs are capable of differentiating into CM-like cells bearing myocyte-specific structural proteins in the presence of a conditioned medium [8]. Although c-kit-positive CPCs are capable of differentiation in vitro [8], they cannot result in cardiac myogenesis in vivo in the adult heart after injury. Notably, CSCs have been isolated from human biopsy specimens, and they can differentiate into functional CMs after an injection into immune-compromised mice. It has been suggested that c-kit-positive CSCs have great therapeutic potential, and these cells have recently been used in a clinical phase 1 trial in patients with ischemic cardiomyopathy [9]. Significantly, autologous c-kit-positive CSC transplantation has been demonstrated to be effective in improving the function of patients with heart failure without producing adverse effects [9]. Nevertheless, this study is an initial trial with a few patients, and it awaits the detailed phase 2 trials.
AP, action potential; CMs, cardiomyocytes; CV, cardiovascular; ECs, endothelial cells; MEF, mouse embryo fibroblast; MHC, myosin heavy chain; MSCs, mesenchymal stem cells; NRCMs, neonatal cardiomyocytes; Sca1, stem cell antigen 1; SP, side population; VSMCs, vascular smooth muscle cells.
Two other phenotypes of CSCs have been identified according to the expression of either stem cell antigen 1 (Sca1) or multidrug resistance gene 1 (MDR1). CSCs expressing Sca-1 in mice and rats lack the expression of hematopoietic stem cell (HSC) markers (CD45 and CD34) or hematopoietic transcription factors (Lmo2, Gata2, and Tal) [10]. Although Sca-1-positive CSCs express early cardiac markers (GATA4, Mef2, and Tef1), they are negative for Nkx2.5 or genes encoding cardiac sarcomeric proteins. In vitro, Sca-1-positive CSCs show sarcomeric structure and spontaneous beating when treated with oxytocin [11]. In vivo, an injection of Sca-1-positive CSCs into mice with ischemic injury has been shown to differentiate into CMs, as evidenced by the expression of sarcomeric α-actin, cTnI, and connexin 43 [12]. A novel subpopulation of Sca-1-positive CSCs has been identified in mice, and these cells have diverse differentiation potential for cardiac regeneration [13]. However, it needs to be determined whether Sca-1-positive CSCs exist in humans. Side-population cells expressing MDR1 in the adult heart are highly enriched for stem cell activity. MDR1-positive CSCs can differentiate into CMs showing spontaneous beating [14]. In vivo, these cells can migrate into damaged myocardium and differentiate into both CMs and endothelial cells after acute ischemic injury [14,15]. These studies suggest that MDR1-positive cells are intrinsic CSCs, and they participate in the regeneration of damaged hearts. However, the mechanisms by which MDR1-positive CSCs contribute to the functional repair of diseased hearts remain unclear.
The cardiosphere is regarded as an important three-dimensional (3D) model of partial cardiogenic differentiation in vitro. In contrast to monolayer cultures, 3D culture technologies mimic cardiac tissue-like morphologies and provide a suitable environment for the identity and integrity of heart structures. Therefore, cardiosphere-derived cells are committed to cardiac and angiogenic differentiation and recruit endogenous regeneration in the peri-infarct zone after transplantation [16]. Significantly, cardiosphere-derived cells have for the first time been used for autologous transplantation in clinical phase 1 trial in the patients with myocardial infarction [17]. The therapeutic regeneration of the heart is encouraging, although it needs further assessment of clinical phase 2 trials.
Increasing evidence suggests that the major cardiac cell lineages arise from a multipotent cell population expressing Flk-1, c-kit, and Isl-1 [18 –21] (Table 1). The expression of Isl-1 is regarded as the onset of myocyte commitment, as Isl1 is a transcriptional activator of the myocyte transcription factor in the Mef2 family. Thus, Isl-1-positive cells in culture would allow for the establishment of the model to unveil the early events of cardiogenesis. However, Isl-1-positive cells are rare in the adult heart, and the functional significance of these cells in cardiac homeostasis is largely unknown.
CM Differentiation from Pluripotent Stem Cells
ES cells are pluripotent stem cells derived from the inner cell mass of the blastocyst, and they are able to produce all cell types (about 260 kinds of cells) in the body. Pluripotent ES cells provide an ideal platform in identifying and characterizing early cell lineage commitment (eg, cardiovascular and hemangioblast lineages) and differentiation of stem cells, which appear hard to study in the embryo. ES cells can be induced by a variety of methods that undergo stepwise differentiation in the embryoid body (EB), mesoderm expressing Brachyury T, CPCs expressing GATA-4, Nkx2.5 and Isl-1, and mature CMs. We illustrate systemic and stepwise CM differentiation of ES cells and growth factors used for the differentiation process (Fig. 2). It has been demonstrated that Oct4 is essential for the differentiation of ES cells into CMs, as evidenced by the fact that the gene silencing of Oct-4 by small interfering RNA in ES cells suppresses their specification toward cardiac lineages [22]. Currently, CMs can be generated from pluripotent stem cells using various approaches, including spontaneous EB differentiation, coculture with endoderm-like (END-2) cells, specific growth factors, small molecules [23]; and the defined biomaterials. We address these methods in detail next. A direct reprogramming strategy has been developed that induces fibroblasts to transdifferentiate into CMs [24]. Significantly, the reprogramming is not involved in pluripotent steps (Fig. 2), and, thus, the generation of CMs by this method is much safer for their clinic application than that using conventional reprogramming toward pluripotency.
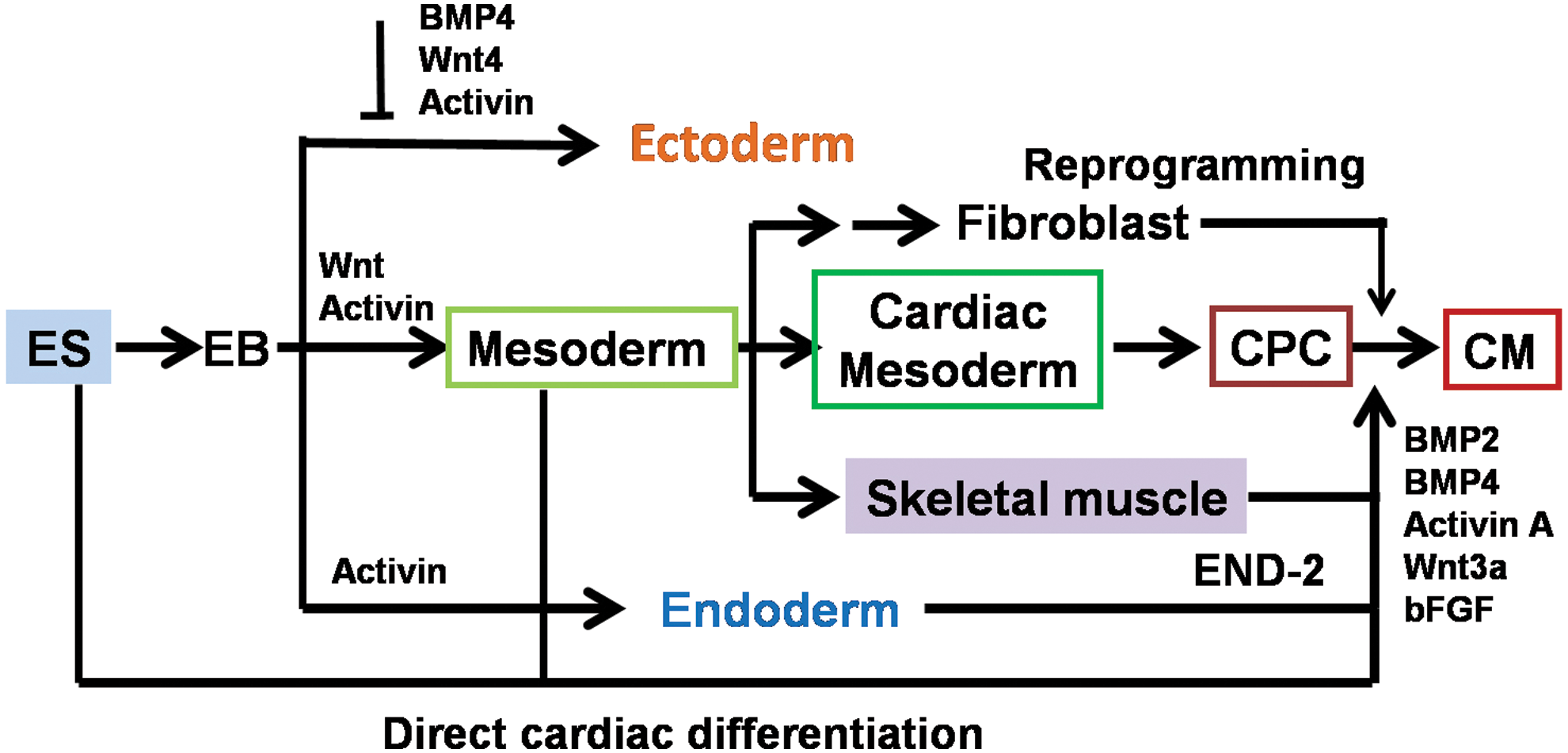
Systemic and stepwise CM differentiation systems from pluripotent stem cells. CPC, cardiovascular progenitor cells; EB, embryoid body; END-2, endoderm-like cells. Color images available online at
Cardiac differentiation from EBs
Human ES cells can differentiate spontaneously into contracting CM-like cells when cultured in suspension and form 3D aggregates called EBs. In a similar fashion to mouse models, human ES cells also begin to differentiate into EBs once they are removed from the mouse embryo fibroblast (MEF) feeder layer. Cardiac differentiation is one of the first cell commitments from human EBs, as evidenced by the expression of numerous markers for mesoderm and early cardiac cell lineages [25]. Human ES cells can form EBs after they are dissociated into small clumps and grow in suspension for several days [26,27]. After plating, rhythmically contracting areas appear in the EBs [26,27]. The EB-based strategy is regarded as the basic approach for initiating CM differentiation from ES cells. Nevertheless, this approach is limited due to a number of factors, including the relatively low yield of CMs and heterogeneous cell populations, and notably not all human ES cell lines have been shown to be capable of generating cardiac cell lineages.
It is possible that CM differentiation from EBs will satisfy the criteria for producing ideally transplantable cells, if the yield and purity of CMs can be significantly improved. As noted, the addition of growth factors and/or small molecules can greatly enhance EBs to a more directed differentiation down the cardiac lineage [28 –32]. Crucial additions include transforming growth factor beta (TGF-β) family members (eg, bone morphogenetic protein and activin), members of the Wnt family, and fibroblast growth factor (FGF) family, all of which are major regulators in cardiac development [33]. The Wnt/β-catenin signaling pathway activator Wnt3A can induce mesendoderm formation [25]. Additionally, vascular endothelial growth factor (VEGF), a p38 mitogen-activated protein kinase (MAPK) inhibitor, stem cell factor (SCF), the Wnt signaling antagonist Dkk1, and ascorbic acid are important for cardiac differentiation from ES cells [19,34,35]. Sulfonyl-hydrazone [36], a glycogen synthase kinase 3 (GSK3) inhibitor, 6-bromoindirubin-3-oxime (BIO) [36], and dorsomorphin [37] have also been identified as possible candidates for improving cardiac differentiation. Notably, the p38 MAPK pathway plays different roles in directing human and mouse ES cells to differentiate into CMs. In human ES cells, p38 MAPK negatively regulates the generation of human ES-derived CMs [35], whereas it promotes the formation of CMs from mouse ES cells [38,39].
Cardiac differentiation in END-2 cell coculture
The cardiac inductive signals from the END-2, which simulates embryonic endoderm, have been used to induce CM differentiation [40,41]. The method is based on the coculture of pluripotent stem cells with visceral END-2 lines, which promote the formation of beating clusters of cells with characteristics of CMs. A serum- and insulin-free medium conditioned by the END-2 has also been shown to induce the effective differentiation of EBs from human ES cells into CMs [42]. Most of the cells derived from pluripotent stem cells share the phenotypic features and electrophysiology of human fetal CMs [41]. These studies suggest that the factors secreted from END-2 are important components for the microenvironment or niche of stem cell differentiation into CMs. However, the efficiency of cardiac differentiation using the END-2 coculture approach is relatively low when compared with growth factor-induction methods [28 –32]. This END-2 coculture method may be improved in the presence of ascorbic acid, because it has been shown that ascorbic acid can enhance the expression of late-stage markers of cardiogenesis [43]. Further identification of inducers for the END-2 coculture would promote more efficient differentiation of pluripotent stem cells into CMs. A protocol has been developed for promoting maturation by culturing the CMs derived from the coculture with END-2 cells in the 3D system [44]. Interestingly, this approach enables a long-term culture of mature CMs with electrophysiological properties for up to 1 year.
CM differentiation with growth factors
Cardiac differentiation from stem cells is controlled by complex networks, and no single growth factor has been identified that induces the effective differentiation of stem cells into CMs. We illustrate the growth factors and their signaling pathways regulating the cardiomyogenesis in Fig. 3. The establishment of the cardiovascular system counts on the members of the TGF-β superfamily, Wnt, Notch, and FGF pathways. Thus, CM differentiation from stem cells can be induced by the addition of specific growth factors. BMP2 and Wnt3 direct the differentiation of pluripotent stem cells toward cardiac lineages in vitro, and the differentiation can be strengthened by coculturing them on MEF secreting bFGF [18]. Bone morphogenetic protein (BMP) has been shown to enhance cardiac differentiation [45]. Members of the TGF-β1 superfamily play crucial roles in many biological processes, including cardiovascular development, as evidenced by the fact that activin A and BMP4 induce the cardiac differentiation of stem cells [45]. Notably, the combination of activin A and BMP4 improves CM differentiation from both human ES cells and the induced pluripotent stem (iPS) cells [45,46], and combinatorial activin receptor-like kinase and bFGF signals promote the cardiac differentiation of human ES cells under feeder- and serum-free culture conditions [47]. Unlike BMP and activin signaling pathways, the Wnt signaling pathway plays different roles in cardiac differentiation in a stage-dependent manner. Wnt signaling activation is essential for the mesoderm formation of ES cells, but the Wnt pathway has to be inhibited for cardiac specification [48,49]. Significantly, cardiac specification by producing 40%∼50% CMs can be induced from ES cells with the Wnt inhibitor Dkk1 [19]. Wnt signaling activates the Notch pathway in early stages of cardiac differentiation [50]. However, the mechanisms of interaction between Wnt and Notch pathways are largely elusive. It has been suggested that some of these signaling pathways act via autonomous pathways in directing the differentiation of CMs, whereas others work through paracrine pathways to regulate cardiac differentiation from ES cells indirectly. Ongoing improvements are being made to use the combinations of different growth factors for the maximal differentiation of CMs.
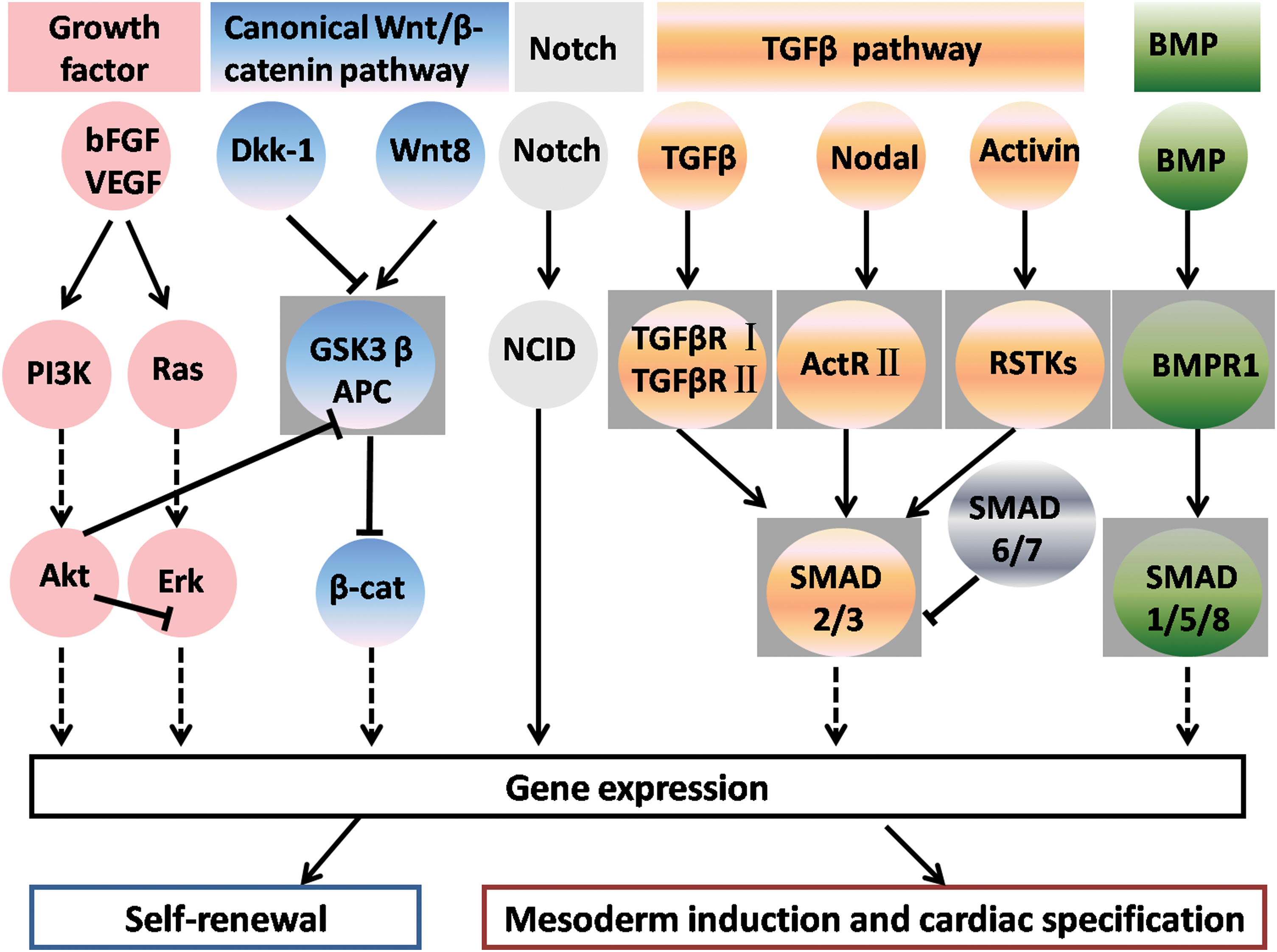
Crosstalk between intracellular signaling molecules and pathways used in regulating self-renewal, mesoderm induction, and cardiac specification. BMP, bone morphogenetic protein; FGF, fibroblast growth factor; TGF-β, transforming growth factor beta; VEGF, vascular endothelial growth factor. Color images available online at
Small molecules for CM differentiation
Small molecules play essential roles in the development of ES cells and, thus, they could become powerful tools for stem cell research and regenerative medicine. Since the function of small molecules in stem cell biology has been well understood, they can be used for directing specific differentiation from ES cells and applications of cell therapy and tissue development in the treatment of human disease. Significantly, a number of commonly used small molecules have been identified in directing the cardiac differentiation of stem cells and we list them in Table 2.
BMP, bone morphogenetic protein; DMSO, dimethyl sulfoxide; EB, embryoid body; FBS, fetal bovine serum; IWR-1, inhibitor of Wnt response 1; IWP-1, inhibitor of Wnt production 1; ND, not determined.
Small molecules that selectively block signaling pathways, including BMP and Wnt signaling, may be valuable for directing the differentiation of ES cells into CMs. As an example, dorsomorphin [37], a selective inhibitor of BMP signaling, has been used to promote cardiomyogenesis from mouse ES cells. The inhibition of the Wnt signaling by small molecules inhibitor of Wnt response 1 (IWR-1) and inhibitor of Wnt production 1 (IWP-1) enhances cardiac differentiation [51]. Recently, inhibitor XAV939 for the Wnt/beta-catenin signaling pathway has been identified as directing cardiomyogenesis in mouse ES cells [52]. These studies illustrate that small-molecule inhibitors for BMP and Wnt signaling pathways are valuable probes for exploring their potential in cardiomyogenesis from stem cells.
Other small molecules, including ascorbic acid [53], isoxazolylserines [54], and dimethyl sulfoxide (DMSO) [43], have also been identified to stimulate the generation of CMs from pluripotent stem cells. The roles of these small molecules are measured by the upregulation of late-stage markers of cardiomyogenesis. Nevertheless, it remains to be determined whether small molecules work directly or indirectly in cardiomyogenesis during ES cell differentiation. Recently, 2 other small molecules, namely Ghrelin and Icariin, have been shown to enhance the generation of CMs from ES cells [55,56]. Ghrelin, a 28-amino-acid peptide, is involved in promoting the differentiation of mesoderm-derived stem cells. An increase in the percentage of beating cells in the EBs is observed from day 6 to 18 of differentiation by Ghrelin [55]. Icariin has been demonstrated as promoting the differentiation of CMs with functional beating from murine ES cells in vitro [56]. In addition, other compounds are discovered as likely affecting specific steps of the cardiogenic program. For instance, a group of chemically related compounds, for example, cardiogenol A-D [57], promote cardiac differentiation at an early stage of ES cells, and microRNAs also play important roles in the cardiac differentiation of ES cells [58]. Although small molecules can be used to enhance cardiac differentiation, CMs with high purity and maturation cannot be currently obtained from ES cells. Thus, optimal methods are required to produce more homogenous and mature CMs rather than cell mixtures.
Biomaterials as an instructive extracellular microenvironment for CM differentiation
Biomaterials have been increasingly used in cell culture and tissue engineering. For stem-cell fate determinations, biomaterials serve as scaffolds for cell self-renewal and differentiation, and they direct the 3D reestablishment and tissue engineering. Biomaterials include naturally derived biomaterials, synthetic and biosynthetic biomaterials, nano-biomaterials, and peptide-immobilized substrates, as we summarize and classify them in Table 3. The methods may include 3D-engineered scaffolds, 3D hydrogel scaffolds, and biodegradable polymer scaffolds. We have used poly(2-acrylamido-2-methyl-propane sulfonic acid sodium salt) (PNaAMPs) and poly(sodium p-styrene sulfonate) (PNaSS) hydrogels that induce mouse ES cells to differentiate into EBs and, eventually, CMs [59]. Our study indicates that PNaSS and PNaAMPS hydrogels facilitate the spreading, migration, and differentiation of EBs into 3 germ layers with a high efficiency of CM differentiation.
3D, three-dimensional; FN, fibroectin; G-CSF, granulocyte colony-stimulating factor; HA, hyaluronic acid; LM, laminin; hESCs, human embryonic stem cells; mESCs, murine embryonic stem cells; PNaAMPs, poly(2-acrylamido-2-methyl-propane sulfonic acid sodium salt); PNaSS, poly(sodium p-styrene sulfonate).
Characterization and identification of pluripotent stem cell-derived CMs
Pluripotent stem cell-derived CMs can be characterized and identified by morphological, phenotypic, and functional features. In morphology, CMs derived from pluripotent stem cells can beat spontaneously and, thus, they are easily detected and identified [41,60]. Beating CMs are round in shape at an early stage, and they become more elongated in shape and accumulate in strands at a later stage. Under electron microscopy, CMs contain myofibrils that are first randomly oriented and then in various manners distributed throughout the cellular cytoplasm. CMs derived from ES cells share certain similarities with adult CMs at ultrastructural levels, that is, both of them have sarcomeres with A, I, and Z bands, gap junctions, desmosomes, and mitochondria [60 –62].
Phenotypically, numerous markers of mature CMs are expressed by pluripotent stem cell-derived CMs [41,60,63 –67]. Cardiac-specific transcription factors, including but not limited to GATA-4, Nkx2.5, Isl-1, Tbx-5, Tbx-20, and Mef2c, are expressed in the pluripotent stem cell-derived CMs [41,60,63 –67]. In addition, CMs derived from stem cells are positive for α-actinin, cardiac troponins T and I, sarcomere myosin heavy chain, atrial- and ventricular myosin light chains, desmin and tropomyosin, as well as for gap junction proteins [841,60,63 –67]. Pluripotent stem cell-derived CMs also express cardiac and muscle-specific proteins, creatine kinase-MB, and myoglobin [63]. Thus, CMs derived from stem cells share biochemical phenotypes with mature CMs.
More importantly, pluripotent stem cell-derived CMs have to be functionally determined. Similar to mature CMs, an increase of intracellular calcium triggers the contraction of CMs derived from pluripotent stem cells [41,68]. Significantly, CMs obtained from pluripotent stem cells can be cultured for a long term, while maintaining their spontaneous contractile ability that can be regulated by isoproterenol and carbachol [60,63,67]. In addition, CMs from pluripotent stem cells possess nodal, atrial, and ventricular types of action potential properties [41,69]. However, CMs derived from pluripotent stem cells show negative force-frequency relations, which is in contrast to mature CMs [70]. Collectively, pluripotent stem cell-derived CMs possess immature phenotypic features and appear to have an immature CM function. The phenotypic characteristics of iPS cell-derived CMs are comparable to those of CMs obtained from human ES cells [46]. Nevertheless, iPS cell-derived CMs may not be used for clinical application due to safety issues involving viral vectors. Since CMs obtained from ES cells become mature over time, novel approaches are required to maintain these cells for a longer time in vitro. Despite the differences of function between pluripotent stem cell-derived CMs and mature CMs, the in vitro CMs generated from stem cells have a functional significance. In addition, the in vitro generation of CMs generated from pluripotent stem cells would offer a useful approach for unveiling the molecular mechanisms of cardiomyogenesis.
CM Differentiation from other Stem Cell Populations
Adult stem cells reside in various adult tissues, and they can self-renew to maintain the pool of stem cells and differentiate into mature cells with particular functions. One of the advantages of using adult stem cells is that there are no ethical issues compared with ES cells, and, importantly, several adult stem cells also have the multipotency or pluripotency to differentiate into various cells which can be used for regenerative medicine. Various types of adult stem cells are shown to differentiate toward cardiac cell lineages. We summarize the advancements in cardiac and extra-cardiac tissue stem cell populations, originating from skeletal muscle, bone marrow, testes, placenta, and adipose tissue, with the potential ability to differentiate into CMs (Table 4 and Fig. 1).
FGF, fibroblast growth factor; SM, smooth muscle; SCF, stem cell factor; TGF-β, transforming growth factor beta; VEGF, vascular endothelial growth factor.
Skeletal muscle stem cells
The skeletal muscle stem cell (myoblast) is first considered for CSC therapy. Myoblasts can proliferate easily in vitro, and they show enhanced resistance under ischemic conditions [71]. Significantly, the transplantation of myoblasts into rodent hearts results in an improved ventricular function and reduces remodeling [72]. However, limitations in the therapeutic potential of skeletal myoblasts for stem cell therapy do exist. First, skeletal myoblasts do not differentiate into cardiac myocytes in vivo but rather develop into myotubes [72]. Additional limitations include the inability of those cells to synchronize with the surviving myocardium, thus posing the risk of arrhythmia and inducing embolic damage because of obstruction to the microvasculature after their administration [1]. Further preclinical and clinical studies are ongoing and may reveal exciting results. In animals, it has been reported that the overexpression of the connexin 43 gap junction protein in skeletal myoblasts can enhance their electrophysiological coupling with cardiac myocytes, suggesting that the risk of inducing arrhythmias can be circumvented [73].
Cardiac differentiation of bone marrow-derived stem cell populations
The bone marrow contains various stem cells, such as HSCs and mesenchymal stem cells (MSCs), with putative cardiac differentiation potential. Bone marrow-derived stem cells have the capacity to differentiate into CMs, opening the possibilities for the use of these cells in stem cell therapy [74]. Bone-marrow HSCs can integrate into the myocardium and improve cardiac function via indirect proangiogenic or other paracrine effects [75]. Studies on the improvement of the ventricular function of HSCs may eventually lead to the design and effective clinical trials in patients. MSCs are multipotent cells that reside in the stromal compartment of the bone marrow, and, notably, these cells could differentiate into beating CMs in vitro, igniting a lot of interest for their applications in cardiac regenerative therapy. Bone-marrow MSCs are suggested as improving the function of infarcted myocardium mainly by promoting angiogenesis or cell survival [76,77]. However, the nature of the mobilizing, migration, and homing signals for bone-marrow MSCs and the mechanism of differentiation and incorporation of these cells into the target tissues need to be explored further. Numerous studies have shown that 5-azacytidine (5-AZA) is an effective reagent for inducing the differentiation of bone-marrow MSCs into CMs [78 –80]. Nevertheless, 5-AZA also induces significant cell apoptosis. To find better inducers in terms of higher efficiency and better suitability for cardiovascular regenerative medicine, a number of growth factors, including TGF-β1, IGF1, dynorphin B, insulin, oxytocin, bFGF, and cardiotropin, have been investigated [81 –84]. Among them, TGF-β1 has proved to be the best cardiac inducer of MSCs toward cardiomyogenesis [83,84]. Clinical trials using MSCs have been performed for patients with acute myocardial infarction (Table 5), probably due to the benefits of stem cell therapy, including improvement of regional wall motion and viability in infarct area, or favorable paracrine effects on the injured myocardium.
CDC, cardiosphere-derived cells; CSCs, cardiac stem cells; CPCs, cardiovasular progenitor cells; EF, ejection fraction; EPC, endothelial progenitor cell; G-CSF, granulocyte-colony-stimulating factor; IC, intracoronary; LVEDV, left ventricular end-diastolic volume; LVEF, left ventricular ejection fraction; MI, myocardial infarction; MNC, mononuclear cells; NA, not applicable; NC, nucleated cells; PCI, percutaneous coronary intervention; SC, subcutaneous; TIA, transient ischemic attack; VT, ventricular tachycardia.
Other stem cell populations from testes, placenta, amniotic fluid, and adipose tissue
Spermatogonial stem cells (SSCs) are a subpopulation of type A spermatogonia in the testis. SSCs were previously regarded as unipotent stem cells, as they were considered as differentiating only into sperm. However, this concept has recently been modified. Notably, a number of recent studies have demonstrated that SSCs from both mouse and human testes can convert in vitro to become ES-like cells that are able to differentiate into various cell lineages of 3 embryonic germ layers [85 –91]. We have revealed the essential roles of Ras/ERK and Smad signaling pathways in regulating the self-renewal and differentiation of mouse SSCs [92,93], and we have recently isolated and cultured human SSCs [94]. Notably, SSCs have important implications in regenerative medicine due to their remarkable plasticity. In mice, cardiac and endothelial cells can be produced from SSCs [88,95]. Diverse cell lineages, such as vascular, heart, and functional CMs, could also be obtained from mouse and human SSCs [85 –91]. These studies provide new promising therapeutic prospects for SSCs to provide new sources of CMs in treating human heart diseases without involving ethical issues and immune rejection concerns.
Human fetal placenta MSCs and chorionic plate cells have been shown to spontaneously beat 3 days after cocultivation with murine fetal CMs [96]. Placenta-derived human fetal MSCs, thus, may be useful for patients who cannot provide bone marrow cells but who need stem cell-based cardiac therapy. Multipotent cells from amniotic fluid have been identified as amniotic fluid stem (AFS) cells [97]. CMs can be obtained from AFS cells when cocultured with rat neonatal cells. AFS cells have cardioprotective roles when xenotransplantated to a rat model of myocardial infarction [98]. Therefore, AFS cells might provide another novel source for the cell therapy of ischemic heart disease. Human adipose tissue-derived stem cells (ASCs) can assume CM activities, including action potential and spontaneous rhythmic beating [99], indicating that ASCs provide a new cell source of CMs. In addition, 2 other cell populations, including stromal vascular fraction cells [100] and adipose-derived stromal cells [101], have been identified. The latter could be induced into CMs in vitro by TGF-β1. Therefore, adipose tissues may represent another source of generating CMs from stem cells.
Transdifferentiation of stem cells and cardiac regeneration
Transdifferentiation, by definition, is a committed stem cell's crossing lineage boundaries and differentiating into cells belonging to another lineage. Transdifferentiation in vivo is still a matter of great debate. It has been reported that a Lin-c-kit+ fraction of bone marrow stem cells can generate 68% of the myocardium comprising myocytes after myocardial infarction in mice [102]. These findings have been challenged by other investigators demonstrating that Lin-c-kit+ stem cells do not differentiate into CMs after an injection into the infarcted heart [103]. Nevertheless, bone marrow-derived stem cells can be reprogrammed to cells expressing cardiac-specific genes in vitro, and endothelial stem cells can transdifferentiate in vitro into cells with cardiomyogenic phenotypes when cocultured with CMs [1]. The differences of stem cell plasticity in cardiac regeneration between in vivo and in vitro are currently unclear, but cell fusion may be involved in the transdifferentiation of stem cells in vitro. Cell fusion is the phenomenon by which stem cells fuse with other cells and the resultant hybrid cells usually assume the more undifferentiated phenotype but possess some characteristics of both cell types. Bone marrow stem cells could fuse with ES cells and adopt the recipient phenotype when cultured with ES cells [104]. Bone marrow cells fuse with CMs, and the resultant hybrid cells are virtually indistinguishable from unfused CMs even at the ultrastructural level [105]. The frequency of fusion is comparable to that of chimeric CMs in animal and human transplantation studies, highlighting the possibility that cell fusion could be a principal mechanism for the transdifferentiation of bone marrow stem cells into CMs. It has also been suggested that circulating cells fuse with CMs at the infarct border zone [106]. However, the fusion of CMs and bone marrow stem cells is uncommon, and the extent to which fusion in vivo and in vitro can reprogram somatic cell nuclei to produce functional CMs requires further investigations.
Potential Applications of CMs Derived from Stem Cells
Human ES cell-derived CMs have been shown to successfully engraft and electromechanically integrate when injected into injured hearts of immunosuppressed animals [60,107]. The transplanted CMs behave as a biological pacemaker and electrically excite the rest of the ventricle. These studies have generated enthusiasm and provided important proof-of-concept data for the generation of biological pacemakers. Cardiac differentiation and the beating cells can also become a useful tool for developmental biology and for uncovering the pathophysiology of human cardiac diseases. It is likely that human ES cell-derived CMs would be applicable for drug discovery and regenerative medicine [108]. Although CMs derived from human ES cells hold great potential for treating heart disease, a number of issues need to be solved so that satisfactory clinical applications can be expected. First, it is required to optimize methods that induce the differentiation of CSCs, human ES cells, and other stem cells into homogeneous, mature, and functional CMs. Second, it is necessary to use proper and sufficient markers to identify the original stem cells and cardiac lineages. We suggest that 2 and more markers should be employed for the identification of any individual cells from stem cells to mature CMs. Third, it is essential to uncover the molecular mechanisms regulating the generation of CMs from various kinds of stem cells, thus leading to obtaining sufficient CMs for basic research and clinical applications.
Clinical trials of stem cell-based cardiac therapy have been currently carried out on 2 different sets of patients, one with acute myocardial infarction and another with chronic left ventricular dysfunction. CSC therapy trials differ from each other due to a number of variables, including the clinical scenario of patients included in the trial, types of stem cells injected, mode of delivery, and predefined end points. We have summarized the salient features of the stem cell clinical trials in acute myocardial infarction to date (Table 5). The interpretation of these trials seems to be hard, because most of them have been either uncontrolled or with a few controls and not randomized. Moreover, differences in trial designs pertaining to the types of stem cell used, patients' status, timing of delivery of stem cells, and methods of measuring outcome contribute to the efficacy of stem cell therapy. Nevertheless, these early trials offer promising therapeutics for heart failure.
Conclusions and Future Perspectives
Much progress has been made to generate CMs from various kinds of stem cells, which provide great promise for their clinical applications. Although clinical trials have been performed in several types of stem cells in acute myocardial infarction, the CMs derived from stem cells are not ready for the clinical use, and numerous basic studies on improving cardiac differentiation from various stem cells are still required. The low efficiency of cardiac differentiation limits the wide applications of stem cell-derived CMs. Therefore, effective methods for generating sufficient and homogenous CMs are prerequisite for their eventually clinical applications. A number of differentiation inducers and factors have to be further investigated so as to provide an appropriate microenvironment or niche for the commitment and differentiation of stem cells into CMs. The integration of small-molecule inducers and stem cell biology to develop stable and efficient cardiac regeneration is required in the near future. Although clinical trials of stem cell therapy in acute myocardial infarction have shown promising results, the molecular mechanisms regulating these modulations are rather unclear. Future studies in stem cells and advancements in safe delivery of defined factors for directing cardiac differentiation are essential for uncovering the mechanisms of cardiomyogenesis and regenerative therapies. In addition, the cell sources for CMs are expected to be derived from extra-cardiac tissues. In summary, we have addressed the advancements made in the differentiation of cardiac and extra-CSC into CMs. Although several questions remain to be answered before the stem cell-derived CMs can be successfully used in clinical therapies, a number of exciting approaches of generating these cells have been developed and clinical trials are under way. The clinical and scientific community are working together to translate stem cell-derived CMs into effective cell therapy for heart diseases.
Footnotes
Acknowledgments
This work was supported by the grants from the National Science Foundation of China (31171422 to ZH and 81130038 to WQG) and the Shanghai Pujiang Program (11PJ1406400 to ZH), and partly by the National Basic Research Program of China (2012CB966800 to WQG) and the Leading Academic Discipline Project of the Shanghai Municipal Education Committee (J50208).
Author Disclosure Statement
No competing financial interests exist.