
Abstract
Human hematopoietic stem/progenitor cells (HSPCs) can be mobilized into the circulation using granulocyte-colony stimulating factor (G-CSF), for graft collection in view of hematopoietic transplantation. This process has been related to bone marrow (BM) release of serine proteases and of the matrix metalloproteinase-9 (MMP-9). Yet, the role of these mediators in HSC egress from their niches remains questionable, because they are produced by nonstromal cells (mainly neutrophils and monocytes/macrophages) that are not a part of the niche. We show here that the G-CSF receptor (G-CSFR) is expressed by human BM mesenchymal stromal/stem cells (MSCs), and that G-CSF prestimulation of MSCs enhances the in vitro trans-stromal migration of CD34+ cells. Zymography analysis indicates that pro-MMP-2 (but not pro-MMP-9) is expressed in MSCs, and that G-CSF treatment increases its expression and induces its activation at the cell membrane. We further demonstrate that G-CSF-stimulated migration depends on G-CSFR expression and is mediated by a mechanism that involves MMPs. These results suggest a molecular model whereby G-CSF infusion may drive, by the direct action on MSCs, HSPC egress from BM niches via synthesis and activation of MMPs. In this model, MMP-2 instead of MMP-9 is implicated, which constitutes a major difference with mouse mobilization models.
Introduction
H
HSPC migration into blood occurs both during development and adult life. During development, HSCs are able to migrate through the circulation from one hematopoietic site to the next (i.e., from the embryonic aorta-gonad-mesonephros region to the fetal liver and finally to the adult BM) [13]. In adult mammals, HSCs are mainly located within the BM, but a small proportion of them has been shown to continually egress into peripheral blood before returning to the BM [14,15]. In addition, HSCs can be mobilized into the circulation using specific growth factors, such as the granulocyte-colony-stimulating factor (G-CSF), to collect a graft for hematopoietic rescue after a high-dose anticancer therapy [16]. Although hematopoietic grafts after G-CSF infusion have been collected from peripheral blood for at least 20 years and are today predominantly used for clinical transplantations, the mechanisms underlying HSC mobilization are not yet fully understood. HPC mobilization has been related to BM release of proteases, including neutrophil serine proteases (i.e., elastase and cathepsin G) and matrix metalloproteinase-9 (MMP-9), which can degrade the extracellular matrix. A local increase of proteolytic activity induces a disruption of the interaction between the microenvironment and HSCs, which involves predominantly the cytoadhesive molecule VCAM-1 and the β1 integrin VLA-4, the growth factor Stem Cell Factor (SCF) and its receptor c-Kit, and the chemokine Stromal cell-Derived Factor-1 (SDF-1/CXCL12) and its receptor CXCR4. Neutrophil elastase has been shown to degrade SDF-1 during G-CSF-induced mobilization, resulting in a dramatic decrease of the BM concentration of this chemokine [17]. Neutrophil elastase can also cleave the extracellular domain of CXCR4 [18] and VCAM-1 [19]. A role for MMP-9 in G-CSF-induced HSPC mobilization has also been shown in a mouse model [20] and suggested in humans [21]. However, neutrophils and monocytes/macrophages, which are the main source of elastase and MMP-9, respectively, are not a part of the HSC niche [12]. Since protease activity is much localized, it is probable that these 2 proteases produced by such cells do not contribute predominantly to HSC egress from the niche.
In this work we have investigated whether G-CSF could directly stimulate human BM MSCs to favor the migration of HSCs/HPCs. To this aim, we used an in vitro model, where CD34+ cells were able to migrate through a BM MSC layer in response to a SDF-1 gradient. Our results demonstrate that human BM MSCs express the G-CSF receptor (G-CSFR), and that G-CSF-stimulated migration of CD34+ cells strictly depends on G-CSFR expression and is mediated by MMP-2, but not MMP-9 or MMP-1. These findings suggest that G-CSF infusion may drive, by a direct action on MSCs, HSPC egress from BM niches via the synthesis and activation of MMP-2. Remarkably, in this molecular model, MMP-2 instead of MMP-9 is implicated, which constitutes a major difference with mouse mobilization models.
Materials and Methods
BM culture-amplified MSCs and sample preparation
BM cells were obtained by iliac crest aspiration from informed and consenting patients undergoing orthopedic surgery (Cardiovascular Surgery Department, Trousseau Hospital, Tours, France). Cells were centrifuged, seeded in flasks at a density of 5×104 per cm2 in an αMEM culture medium (Invitrogen) supplemented with 10% screened fetal calf serum, 20 μM L-glutamine (Invitrogen), 100 U/mL of penicillin G (Invitrogen), and incubated at 37°C in 95% humidified air and 5% CO2. Adherent culture-amplified MSCs were used at passage 2 (P2). When P2 layers became confluent, the supernatant was entirely replaced by a migration medium consisting in RPMI with 0.25% bovine serum albumin (Sigma-Aldrich) alone (control), or supplemented for 3 consecutive days with 15 ng/mL/day G-CSF (Neupogen®; Amgen) or 15 U/mL/day IL-1β (R&D Systems). For enzyme-linked immunosorbent assay (ELISA), supernatants were removed, filtered, and stored at −80°C, whereas cells were trypsinized and collected for RNA extraction and flow cytometry. For zymography and western blot assays, membrane fractions were isolated. Cells were homogenized in a Dounce homogenizer. After centrifugation at 500 g to remove nuclei and large-cell debris, supernatants were centrifuged at 100,000 g to pellet the membranes. Supernatants were then retained as the cytosolic fraction of the cell lysates.
Trans-stromal migration assay
Trans-stromal migration assays were performed in Transwell inserts (Costar) with 5-μm-pore filters using hematopoietic cells, including megakaryocytic leukemia-derived MO7e cells (kindly provided by Dr Fawzia Louache, Institut Gustave Roussy) and human BM-derived CD34+ cells. Human BM CD34+ cells were collected from informed and consenting orthopedic surgery patients and enriched using a magnetic bead separation kit (MiniMACS®; Miltenyi Biotec), according to the manufacturer's instructions (mean purity of CD34+ cells within the selected cells was 90%). The filter upper face of Transwell membranes was coated with fibronectin (Sigma-Aldrich) and seeded with 104 P2 MSCs in the culture medium. Two days later, the culture medium was replaced with a migration medium, and cells were stimulated, or not, for 3 consecutive days with 15 ng/mL/day G-CSF (filgrastim, Amgen) or 15 U/mL/day IL-1β (R&D Systems). Twenty hours after the last stimulation, 50×104 MO7e or 2.2×104 CD34+ cells were loaded into the upper chamber in 100 μL of migration medium, and 600 μL of migration medium supplemented with 100 ng/mL SDF-1α (R&D Systems) was added to the bottom chamber. The Transwell plates were then incubated at 37°C, 5% CO2, for 4 h. The percentage of cell migration was the ratio (×100) between the cell number recovered in the lower compartment and the number of loaded cells. To investigate the role of MMPs in cell migration, the MMP inhibitor GM 6001 (2×10−5M; Calbiochem, Merck Chemicals Ltd.) or its negative control was added before migration. To identify the specific MMPs involved, additional blocking experiments were performed using anti-MMP-1 (clone 41-1E5; Oncogene Research Products), anti-MMP-2 (clone CA-4001; Chemicon) or anti-MMP-9 (clone GE-213; Chemicon) monoclonal antibodies (mAbs) in comparison to an isotypic negative control mAb (IgG1).
G-CSF receptor expression
Western blot
G-CSFR expression was explored in BM MSCs untreated or transfected with shRNA, and membranes were isolated as described above. A total of 5 μg protein was separated using 15% (w/v) acrylamide sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis and electrophoretically transferred onto polyvinylidene difluoride (PVDF) membranes. PVDF membranes were blocked with 5% dried milk, incubated with primary mouse anti-human G-CSFR (clone 38660; R&D Systems) mAb, and then with a secondary rabbit polyclonal IgG anti-mouse antibody (Ab) conjugated to horseradish peroxidase (Biorad). Bands were revealed by chemiluminescence using the ECLplus® Western blotting detection kit (GE Healthcare Europe GmbH) and quantified with a Chemismart 2000® Imager (Vilber Lourmat).
Flow cytometry
BM MSCs and normal peripheral blood neutrophils were incubated with primary mouse anti-human G-CSFR (clone 38660; R&D Systems) mAb and subsequently with an R-phycoerythrin-conjugated goat anti-mouse Ab after permeabilization with the BD Cytofix/Cytoperm® kit (BD Biosciences).
RNA-interference targeting the G-CSF receptor
Cells were transfected with 29-mer anti-human CSF3R (G-CSFR) shRNAs in the pRS plasmid (ref #TR313700; OriGene;
Protease expression in BM MSCs
Quantitative reverse transcriptase–polymerase chain reaction
We studied 12 protease and inhibitor genes potentially involved in cell migration. In addition, we evaluated a total of other 94 genes encoding proteins related to cell adhesion, extracellular matrix, cytoskeleton, growth factors, cytokines, and chemokines (see Supplementary Data). mRNA expression was assessed by TaqMan real-time reverse transcriptase–polymerase chain reaction (RT-PCR). P2 confluent layers of unstimulated and stimulated BM MSCs were lyzed directly in the culture dishes, and total RNA was isolated using the RNeasy Kit® (Qiagen;
Flow cytometry
Membrane and intracellular MMP-2 and MMP-9 proteins were detected by flow cytometry (FACSCalibur®; BD Biosciences). Cell suspensions (1×106 cells/mL) were preincubated at 37°C with 1 mL of GolgiPlug® (BD Biosciences), an inhibitor of protein transport, for 4 h. Then, cells were sequentially incubated with specific mouse anti-MMP-2 or anti-MMP-9 mAbs, biotinylated goat anti-mouse polyclonal IgG (BD Biosciences), and R-phycoerythrin-conjugated Streptavidin (Dako Cytomation). For intracellular expression, cells were first permeabilized with the BD Cytofix/Cytoperm® kit (BD Biosciences). MMP-2 and MMP-9 levels (active and latent forms) in MSC supernatants were measured by ELISA using the Quantikine® human MMP-2 and MMP-9 immunoassays (R&D Systems), according to the manufacturer's instructions. MMP-2 and MMP-9 gelatinase activities were measured by gelatin zymography in BM MSC membrane and cytosolic fractions, as previously described [23].
Immunofluorescence
BM MSCs were cultured on Lab-Tek® chamber slides (Nunc). Confluent layers were fixed and permeabilized with 2% formaldehyde in PBS. Slides were incubated with anti-human MMP-2 (clone VB-3, Oncogene) or MMP-14 (clone 114-6G61) mouse Ab for 1 h, followed by Alexa-488- or Alexa-594-conjugated goat anti-mouse Ab (Interchim). After 30 min, slides were mounted with Vectashield+DAPI® (Vector Laboratories Ltd.) and examined under a Leica DMR/HCS fluorescence microscope (Leica).
Statistical analysis
Paired values from different experimental groups were compared using the nonparametric Wilcoxon's test with a significance level set at P<0.05.
Results
The G-CSF receptor is expressed by BM MSCs
P2 adherent culture-amplified cells, systematically checked for the absence of contaminating hematopoietic cells (CD45+, CD34+, or CD14+ cells), have been extensively characterized at the plasma membrane level [24], showing a typical MSC phenotype that includes the CD73, CD90, and CD105 markers. Their multipotency toward the adipogenic, osteogenic, and chondrogenic and vascular smooth muscle lineages have also been previously reported [22], and the hematopoietic supportive capacity of the MSCs used for the assay was verified (Supplementary Fig. S1; Supplementary Data are available online at
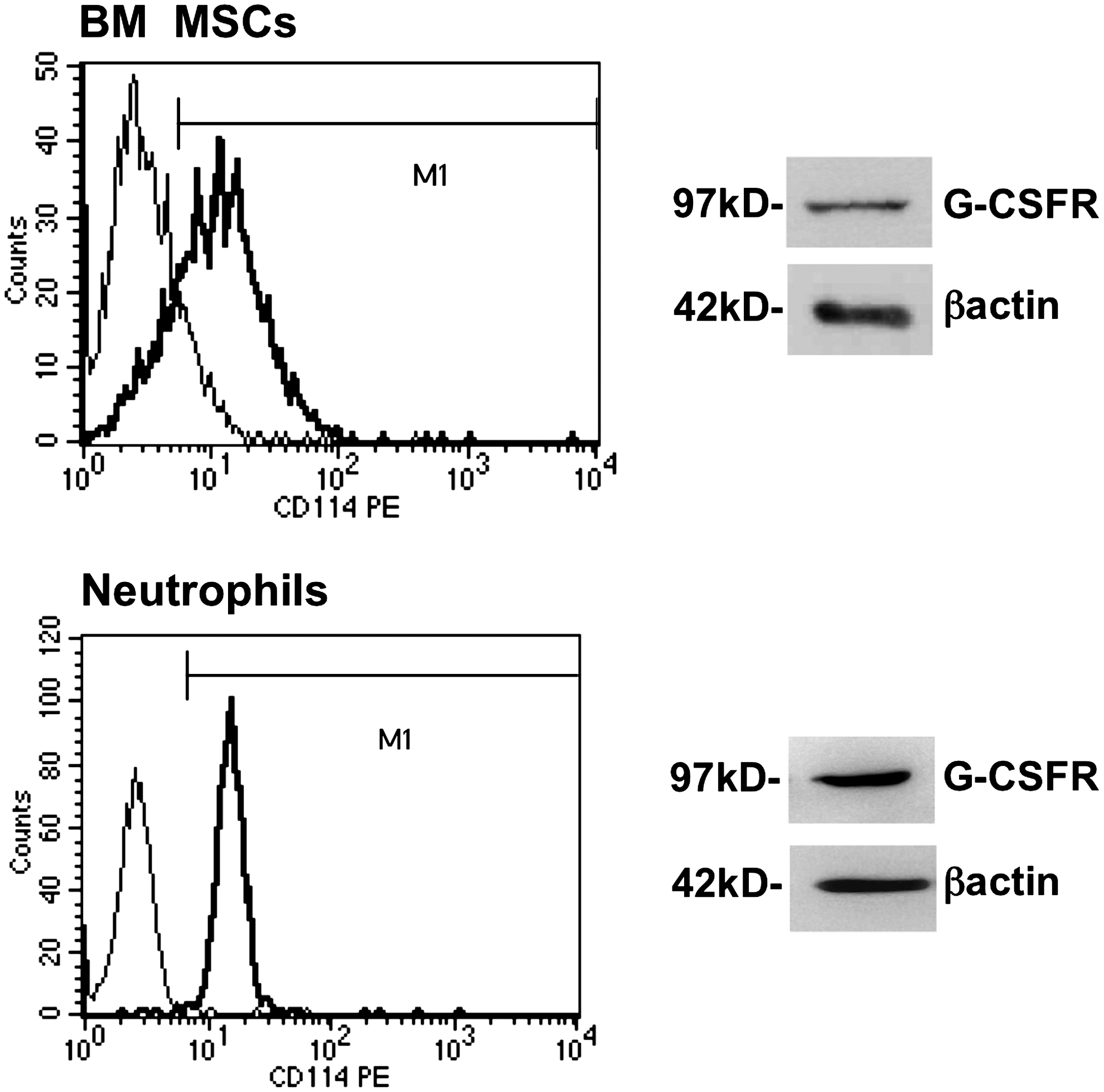
Granulocyte-colony-stimulating factor receptor (G-CSFR) expression in bone marrow mesenchymal stem/stromal cells (BM MSCs) assessed by flow cytometric and western blot analyses. Flow cytometry histograms show the level of expression of G-CSFR (thick line) compared to the isotypic negative control (thin line). Western blots were performed on BM MSC membrane fractions with an anti-human G-CSFR or anti-β Actin antibody (loading control). Human neutrophils were used as positive control for G-CSFR expression. Results are from one representative experiment (out of 3).
G-CSF stimulation of human BM MSCs enhances trans-stromal migration of immature hematopoietic cells
To model the interactions between hematopoietic cells and BM stromal MSCs, an in vitro trans-stromal migration assay was set up in which human immature hematopoietic cells (i.e., BM CD34+ or MO7e cells) migrated through a confluent BM MSC layer grown on fibronectin-coated Transwell filters in response to an SDF-1 gradient. Using this assay, we investigated whether G-CSF (compared to IL-1β, a classical cytokine inducer of mesenchymal cells) could exert a direct effect on BM MSCs to favor the migration of hematopoietic cells through the stromal layer. To this aim, the BM MSC layers were stimulated, or not, with G-CSF or IL-1β for 3 days before performing the trans-stromal migration assay. The percentage of cells that migrated through G-CSF-stimulated MSCs was significantly higher than that when using unstimulated MSC layers for both MO7e, a G-CSF-irresponsive megakaryoblastic cell line [25] (24%±0.9% vs. 9%±0.6%; P<0.001; Fig. 2A), and BM CD34+ cells (38.9%±2.8% vs. 20.2%±2.1%; P<0.01; Fig. 2B). In contrast, IL-1β pretreatment did not have any effect on trans-stromal migration of MO7e cells (Fig. 2A). We verified that, in the absence of a stromal layer, G-CSF did not affect the migration of MO7e cells through filters devoid of any cell (data not shown), which is in agreement with the fact that these cells are insensitive to G-CSF [25].
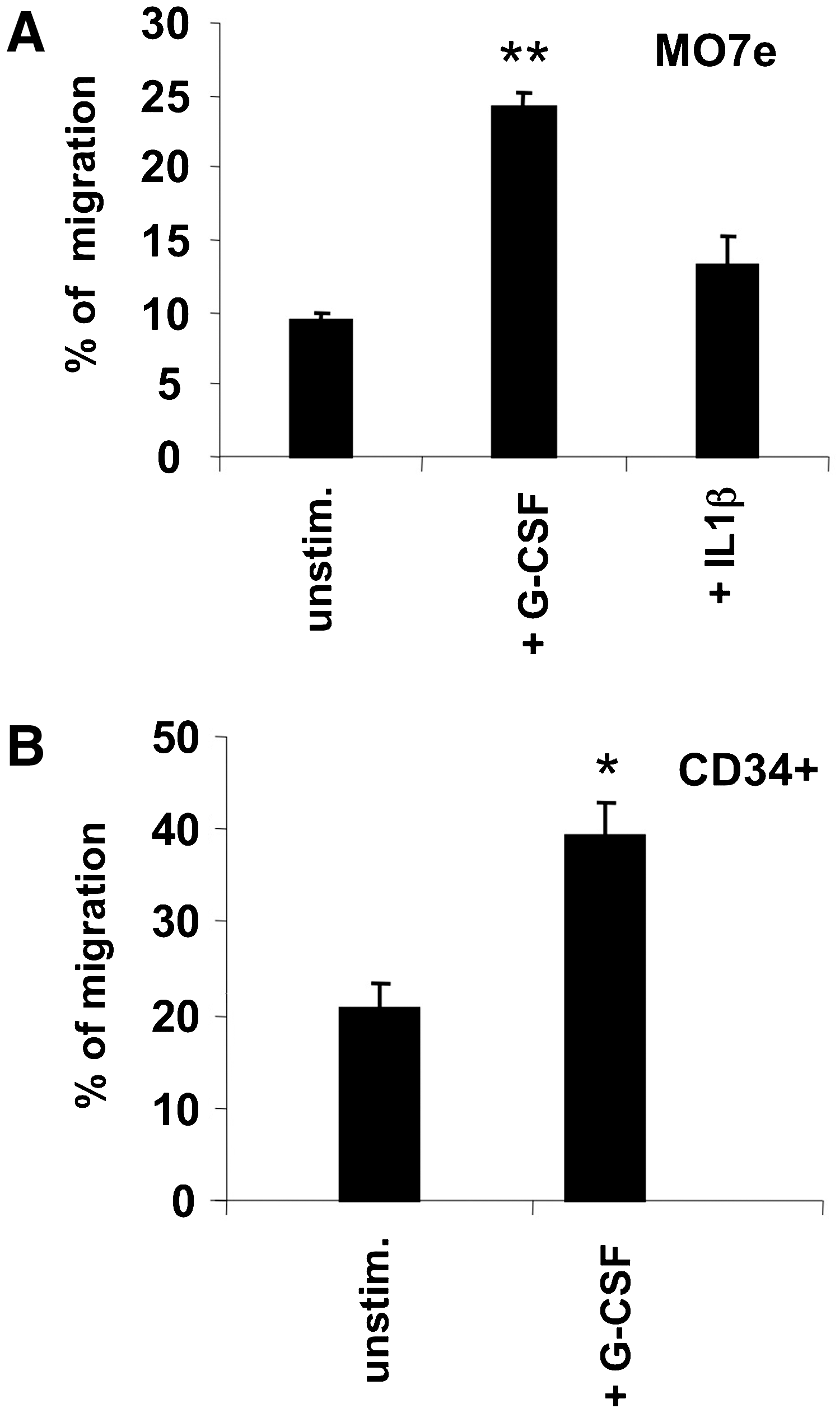
Effect of G-CSF on in vitro trans-stromal migration of hematopoietic cells. Columns represent the average percentage (and bars SEM) of migration through unstimulated, G-CSF-, or IL-1β-stimulated MSCs.
ShRNA-induced downregulation of the G-CSF receptor in BM MSCs inhibits the stimulating effect of G-CSF on CD34+ cell trans-stromal migration
shRNA interference was used to downregulate GCSFR to demonstrate the functional role of G-CSFR expressed in BM MSCs during migration of HPCs. First, downregulation of G-CSFR expression was verified by western blotting after transfection of specific shRNAs against human GCSFR in BM MSCs (Fig. 3A). The transfection procedure did not affect trans-stromal migration, since the percentage of migrating cells did not differ between assays carried out using untransfected MSCs and MSCs transfected with the negative controls, including empty shRNA-cloning vector (shRNA1) and noneffective shRNA (shRNA2), with (black columns) or without (white columns) G-CSF prestimulation (Fig. 3B). Conversely, trans-stromal migration of BM CD34+ cells strongly decreased when using G-CSF-stimulated MSCs transfected with the 4 G-CSFR-specific shRNAs (shRNA3 to 6) (Fig. 3C).

Downregulation of the G-CSF receptor expression in BM MSCs by shRNA interference and effect on G-CSF-stimulated trans-stromal migration of CD34+ cells.
MMP-2 (but not MMP-9) is expressed in human BM MSCs and is activated by G-CSF
To clarify the role of MSC-derived molecules in HSPC migration, we first evaluated potential changes of protease mRNA expression induced by G-CSF in the human BM MSCs we used for the transmigration assay. The expression of 12 protease genes that included those encoding MMPs, their inhibitors TIMP-1 and TIMP-2, CD26/dipeptidylpeptidase IV, and urokinase plasminogen activator receptor (PLAUR) was analyzed by real-time RT-PCR in BM MSCs stimulated or not by G-CSF. The protease genes that displayed the highest expression level were those for MMP-2 and MMP-14/MT1-MMP with the inhibitors TIMP-1 and TIMP-2, whereas those for CD26 and PLAUR were expressed at a lower level and that for MMP-9 was virtually undetectable (Fig. 4A). No clear G-CSF-induced change in protease expression was observed at the mRNA level. Of particular interest, the gelatinase expression profile in mouse MSCs appeared totally at the opposite to that seen in humans, since in mouse MSCs, only MMP-9 was expressed without any expression of MMP-2 (Supplementary Fig. S2).

Protease expression assessed in BM MSCs stimulated or not by G-CSF.
In addition, we investigated whether other genes encoding proteins involved in cell migration or hematopoiesis regulation were modified by G-CSF treatment of human MSCs (Supplementary Fig. S3). A total of 94 genes cell involved in cell adhesion, including extracellular matrix and cytoskeleton components (Fig. S3A), growth factors and cytokines (Fig. S3B), and chemokines (Fig. S3C), were studied. Again, no clear G-CSF-induced change was observed, except for EGF and CCL5 (RANTES) gene expression that was upregulated (mean yields of 3.03 and 3.61, respectively). Noteworthy was the high expression of the CXCL12 (SDF-1) gene by BM MSCs, which was not influenced by G-CSF-treatment.
Considering the crucial role of the gelatinases MMP-2 and MMP-9 in HSPC mobilization [20,26,27] and despite the lack of influence of G-CSF on their mRNA levels, we assessed MMP-2 and MMP-9 expression at the protein level. In MSC supernatants, only MMP-2, and not MMP-9, was detected by ELISA (Fig. 4B), but the MMP-2 expression level was not modified by G-CSF treatment. MMP-2 was also detected by flow cytometry, weakly at the MSC cell membrane and more strongly intracellularly, with again no clear change of expression after G-CSF treatment (Fig. 4C). In agreement with the ELISA results, MMP-9 was not detected either at the membrane or intracellularly. MMP-2 expression was also confirmed by immunofluorescence (Fig. 4D) in BM MSCs together with that of MMP-14/MT1-MMP, which is the major MMP-2 activator [28].
The antigenic detection of MMPs does not necessarily reflect their functionality, since MMPs are secreted as inactive (latent) zymogens (i.e., pro-MMPs) that have to be activated by cleavage to reveal their proteolytic activity. Therefore, we assessed the gelatinolytic activity of both MMP-2 and MMP-9 by zymography in membrane and cytosolic fractions of unstimulated and G-CSF-stimulated BM MSCs. Pro-MMP-2 (72 kD) was detected in both membrane (Fig. 5A) and cytosolic (Fig. 5B) fractions of unstimulated MSCs, and its expression increased in both fractions after G-CSF-treatment. Active MMP-2 (62 kD) was exclusively detected in membrane fractions and increased upon G-CSF stimulation (in 2 of 3 experiments) (Fig. 5A). The absence of MMP-9 expression in BM MSCs was confirmed by zymography, since neither the latent (92 kD) nor the active (83 kD) form could be detected in any of the cell fractions (Fig. 5C).
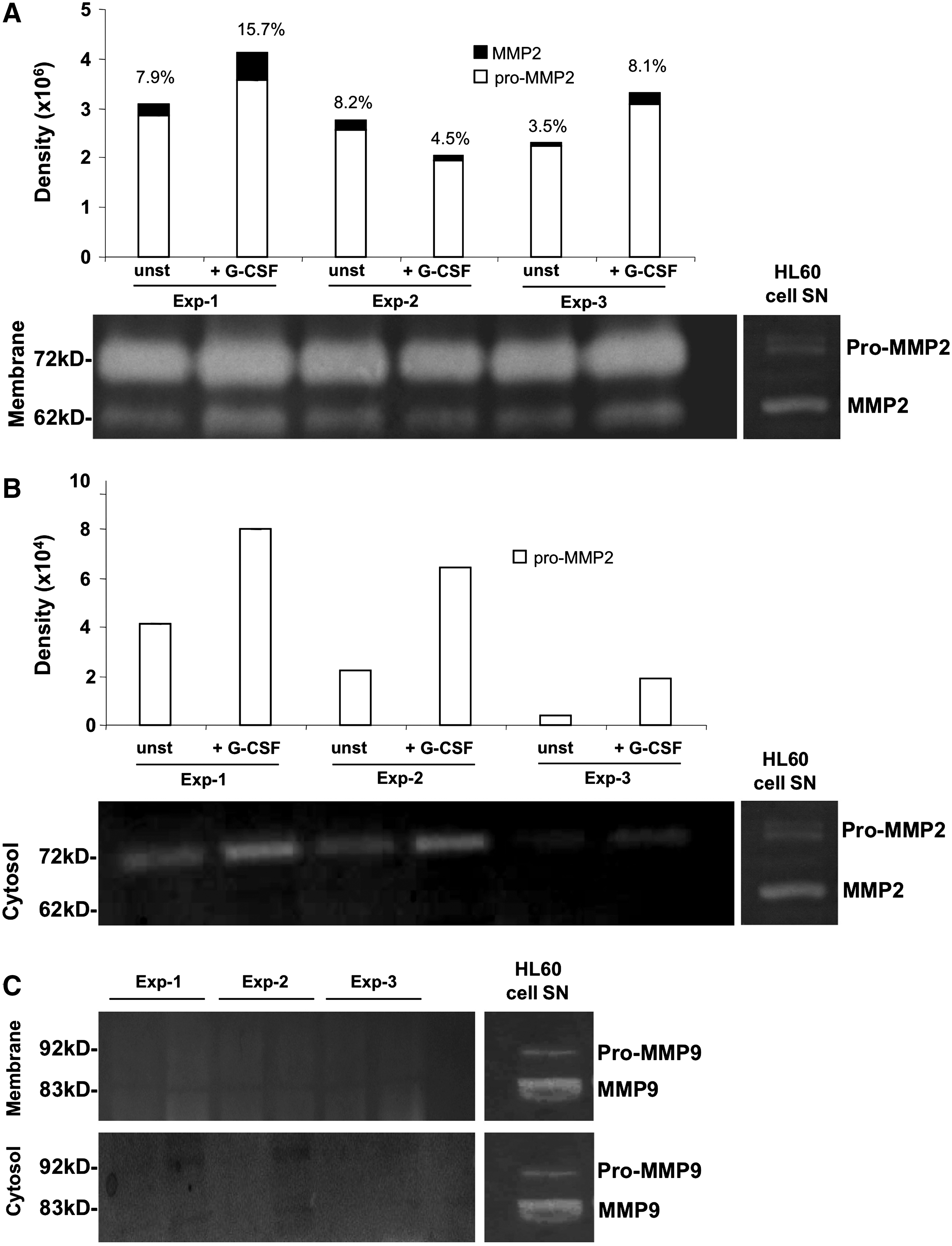
Effect of G-CSF on MMP-2 expression in BM MSCs assessed by zymography. Membrane and cytosolic fractions were isolated from G-CSF-stimulated or unstimulated (unst) BM MSCs to assess the gelatinolytic activity by zymography. The figure shows zymograms of membrane
MMP-2 (but not MMP-9 or MMP-1) is involved in the G-CSF stimulation of trans-stromal migration of hematopoietic cells
We hypothesized that G-CSF stimulation of HSPC trans-stromal migration could be linked to the local production of MMPs observed in BM MSCs. To test this hypothesis, trans-stromal migration assays with BM CD34+ or MO7e cells were carried out in the presence/absence of MMP inhibitors. First, the broad spectrum MMP inhibitor GM 6001, which exerts an inhibitory effect on MMP-1, 2, 3, 8, and 9, and a negative control molecule provided by the manufacturer were used. GM 6001 decreased significantly (P<0.01) the percentage of MO7e cells that migrated through the G-CSF-stimulated MSC layer from 26.2%±1.8% to 16.6%±0.9%, which represents an inhibition of 36% (Fig. 6A). In contrast, no inhibitory effect was seen with unstimulated or IL-1β-stimulated MSCs. To identify which MMPs were involved in this G-CSF-enhanced migration, trans-stromal migration assays were performed in the presence of mAbs that specifically neutralize the activity of MMP-1, MMP-2, and MMP-9 (each at the optimal blocking concentration of 5 μg/mL). The migration of both MO7e (Fig. 6B) and CD34+ cells (Fig. 6C) was significantly inhibited only by the anti-MMP-2 mAb, whereas no inhibition was observed with anti-MMP-9, anti-MMP-1, or irrelevant anti-IgG1 (negative control) Ab. Specifically, the percentage of migrated cells decreased from 23.0%±1.4% to 9.7%±0.9% (58% inhibition; P<0.001) for MO7e cells, and from 35.4%±2.2% to 21.3%±3.0% (40% inhibition; P<0.01) for CD34+ cells.
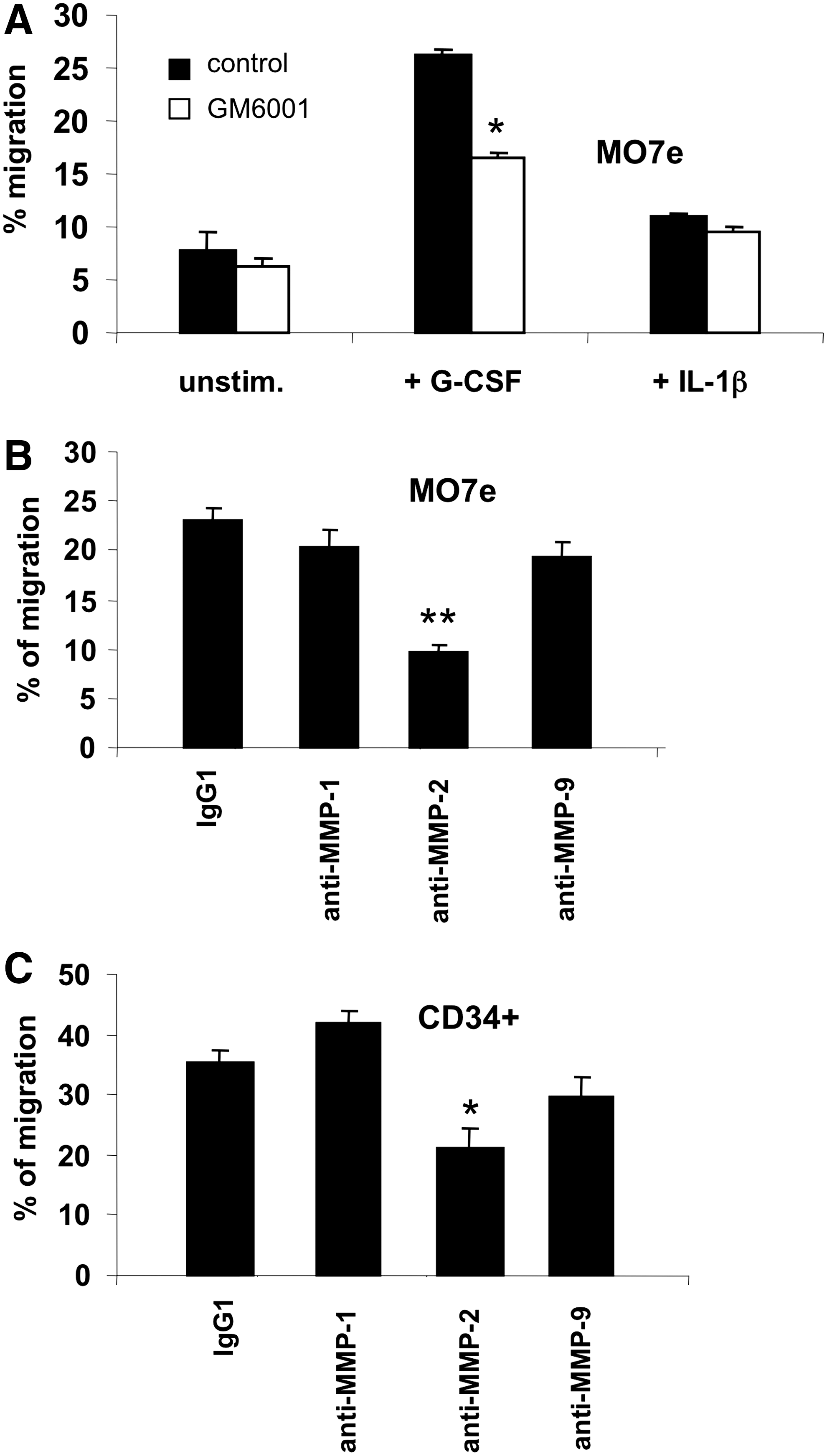
Role of MMP-2 on the G-CSF-stimulated trans-stromal migration of hematopoietic cells.
Discussion
In the present study, we modeled the mobilization of HSPCs using an in vitro migration assay that allowed us to demonstrate a direct role of G-CSF on the migration of HSPCs through BM MSCs. G-CSF activity was dependent on G-CSFR expression in BM MSCs and was mediated by an MMP-2-dependent mechanism.
First, we investigated whether G-CSFR/CD114 was expressed in BM MSCs. Much recently G-CSFR has been reported to be expressed in mouse BM MSCs, the proliferation of which is increased in response to G-CSF [29]. In this work, we report, for the first time to the best of our knowledge, that G-CSFR is expressed in human BM MSCs. We detected this receptor by means of flow cytometry and western blotting. In western blot experiments, the receptor was clearly evidenced only in plasma and the internal membrane fraction, since the protein was too diluted to be detectable in total cell extracts. The G-CSFR detected was apparently not different from that present in peripheral blood neutrophils, both proteins having the same molecular weight of 97 kD.
Secondly, we showed that pretreatment of BM MSC layers with G-CSF increased the trans-stromal migration of both MO7e cells, a G-CSF-irresponsive megakaryoblastic cell line [25], and normal CD34+ cells. This effect on MSCs appeared to be specific for G-CSF, since IL-1β, a classical cytokine and cell adhesion molecule inducer in mesenchymal cells, did not exhibit such effect.
Thirdly, using a shRNA interference strategy to target G-CSFR, we demonstrated that trans-stromal migration of CD34+ cells after G-CSF-stimulation was strictly dependent on the expression of G-CSFR by BM MSCs. Taken together, our findings suggest that infused G-CSF acts directly on BM MSCs to enhance the egress of HSPCs from the niche, although an additional indirect mechanism cannot be excluded after BM MSCs had been in contact with CD34+ cells. The fact that BM MSCs directly respond to G-CSF via its receptor may explain some poorly understood data such as the effect of G-CSF on human stromal cells to enhance CD34+ cell adhesion through VCAM-1 increase under flow conditions [30] and to reduce BM osteoblast density and activity [31]. A recent report has shown that the in vivo administration of G-CSF in mice induces the downregulation of SDF-1, SCF, angiopoietins, and VCAM-1 transcripts in nestin+ cells that constitute a subset of perivascular stromal cells intimately associated to HSCs [12]. Our data on human MSCs are consistent with the latter study that showed a decrease of VCAM1 and ANGPT2 expression after G-CSF treatment (Supplementary Fig. S3A, B), although not reaching significance. Otherwise, the G-CSF-induced upregulation of EGF and CCL5 genes we observed (Supplementary Fig. S3B, C) should have no (or little) impact on the HSPC mobilization process.
Moreover, we investigated whether metalloproteases were implicated in the migration of HSCs/HPCs through G-CSF treated BM MSCs. Several reports have suggested the involvement of MMP-9 (Gelatinase B) in HSPC mobilization [21,26]. Although such role of MMP-9 has been demonstrated in a mouse model [20], the implication of this MMP remains controversial [32]. The present study excludes such role in humans, since MMP-9 expression in BM MSCs was marginal to nil in terms of transcripts (by RT-PCR), secreted proteins (by ELISA), and cell-associated proteins (by flow cytometry and immunofluorescence). In contrast, MMP-2 and MMP-14/MT1-MMP were detected using several methods, including quantitative RT-PCR and immunofluorescence. Moreover, using the very sensitive zymography functional assay, we found that the G-CSF treatment increased not only the latent MMP-2 (pro-MMP-2) in the cytosol but also the activated MMP-2 at the BM MSC membrane. MMP-2 (gelatinase A) shares with MMP-9 similar biological activities, including not only the degradation of denatured collagens (gelatins) and of native collagens IV and V but also the inactivation of the chemokine SDF-1/CXCL12 [33]. All these enzymatic activities of MMPs play a role in the mobilization of HSPCs from the niche. Finally, we demonstrate that the enhancing effect of G-CSF on trans-stromal migration of hematopoietic cells is largely dependent on MMPs, using the chemical inhibitor GM6001, and more specifically on MMP-2 (and not MMP-1 or MMP-9), using blocking antibodies. Thus, MMP-2 could be a crucial (but not exclusive) mediator involved in G-CSF-induced mobilization of HSCs out of the stem cell niche. The mechanism by which G-CSF induces the activation of MMP-2 at the BM MSC membrane remains to be clarified. A possible role of MT1-MMP (also shown to be expressed at the membrane) can be evoked, since this membrane-anchored MMP is known to be the major MMP-2 activator in the presence of TIMP-2 [28] and to play a crucial role in BM MSC migration [34 –36]. Interestingly, a recent report indicates that G-CSF treatment of human CD34+ cells induces expression and activation of MT1-MMP and mediates pro-MMP-2 activation and CD34+ cell migration across a reconstituted basement membrane [27]. Another very recent study suggests a contribution of BM osteoblasts in the mobilizing process of HSPCs by degrading CXCL12 through the secretion of B, K, and L cathepsins [37].
Collectively, our results show that G-CSF directly stimulates BM MSCs to induce MMP-2 synthesis and its activation at the membrane. MMP-2 might be necessary to remove the most primitive HSCs from BM niches, whereas serine proteases and MMP-9, locally produced by neutrophils or monocytes/macrophages, might rather be involved in the trans-endothelial migration of HSPCs and their egress into the blood stream. The molecular model of HSPC mobilization from the niche described here applies to humans, but not to mice, in particular because of the role of MMP-2 instead of MMP-9.
Footnotes
Acknowledgments
The authors are grateful to Elfi Ducrocq and Marie-Christine Bernard for their expert technical assistance. This work was supported by grants from the European Integrated Projects FIRST (contract N° 503436) and Genostem (contract N° 503161), the French Cancéropôle Grand-Ouest, the French Council of Centre Region and the International Rotary Clubs of Blois and Tours-Balzac.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.