
Abstract
Geminin is a nuclear protein that performs the related functions of modulating cell cycle progression by binding Cdt1, and controlling differentiation by binding transcription factors. Since embryonic stem cells (ESC) and the epiblast share a similar gene expression profile and an attenuated cell cycle, ESC form an accessible and tractable model system to study lineage choice at gastrulation. We derived several ESC lines in which Geminin can be inducibly expressed, and employed short hairpin RNAs targeting Geminin. As in the embryo, a lack of Geminin protein resulted in DNA damage and cell death. In monolayer culture, in defined medium, Geminin supported neural differentiation; however, in three-dimensional culture, overexpression of Geminin promoted mesendodermal differentiation and epithelial-to-mesenchymal transition. In vitro, ESC overexpressing Geminin rapidly recolonized a wound, downregulated E-cadherin expression, and activated Wnt signaling. We suggest that Geminin may promote differentiation via binding Groucho/TLE proteins and upregulating canonical Wnt signaling.
Introduction
At gastrulation, the epiblast is allocated to 3 germ layers: endoderm, mesoderm, and ectoderm. This process involves an epithelial-to-mesenchymal transition (EMT), complex morphogenetic movements, and differential gene expression that culminate in lineage-fate decisions. Despite the importance of understanding cell fate choice in early development, progress has been limited by the early lethality of many gene-null models and the general inaccessibility of the mammalian embryo at this early stage. Since embryonic stem cells (ESC) are derived from the inner cell mass of the blastocyst, they retain the ability to generate all cells of the mouse embryo [5,6]. In addition, ESC and the epiblast have similar gene expression patterns and share an uniquely abbreviated cell cycle. Because of these similarities, differentiation of ESC offers a simple model of cell fate choice at gastrulation and an experimentally tractable system to examine the gene function in development.
One gene that has been shown to modulate both cell cycle progression and differentiation is Geminin, which was identified in 2 independent functional screens in Xenopus embryos: the first screen to identify genes that were degraded during mitosis [7], and the second to identify genes involved in neural induction [8]. Geminin functions as a cell cycle-licensing factor, permitting replication of the genome once and only once during mitosis [9,10]; Geminin-null mouse embryos die by E3.5 due to endoreduplication errors [11,12]. Late in the S phase, Geminin binds and inhibits the interaction of Cdt1 with DNA origins of replication, and must then be degraded during the M phase to allow replication.
Geminin overexpression was reported to expand the Xenopus neural plate by inhibiting BMP signaling, independent of an effect on cell division [8]. Expression of Geminin is restricted to the neural plate in the Xenopus embryos by the Tcf and Vent sites [13], indicating that BMP and Wnt pathway signaling cooperate to control Geminin expression during differentiation. Geminin has also been proposed to control the transition from pluripotency to differentiation in the Xenopus embryo by epigenetically repressing the transcriptional response to the Activin/Nodal, FGF, and BMP pathways in conjunction with polycomb proteins, thus promoting differentiation of the neural ectoderm and inhibiting non-neural lineages [14].
To determine how Geminin functions in lineage specification, we developed several ESC lines in which Geminin can be inducibly overexpressed, and employed short hairpin RNAs (shRNA) to target the native Geminin mRNA. Reduction of Geminin protein via targeted shRNA resulted in cell death due to DNA damage. In monolayer culture, in defined medium, Geminin overexpression supported differentiation of neural precursor cells and neurons. In embryoid bodies (EBs); however, overexpression of Geminin induced mesendodermal differentiation and expression of genes involved in EMT. Initiation of mesendodermal differentiation appears to result from Wnt pathway activation possibly by binding of Geminin to Groucho/Transducin-Like Enhancer of split (TLE) proteins in the nucleus that block Tcf/Lef target gene expression in the absence of activated β-catenin [15].
Materials and Methods
ES cell culture and differentiation
Undifferentiated mouse ESCs were maintained in 0.1% gelatin-coated tissue culture flasks in a complete medium composed of DMEM (Invitrogen), 10% fetal bovine serum (Atlanta Biologicals), 50 mM HEPES (Sigma), and 1 mM β-mercaptoethanol (Sigma) with 5 ng/mL LIF (Chemicon). Neural-permissive culture conditions were achieved by plating cells at low density (∼2.0×104 cells per cm2) in gelatin-coated 6- or 12-well plates in 20% Neural basal medium (Invitrogen), 80% Ham's F12 medium (Invitrogen) with N2 and B27 salts (Invitrogen), 1 mM retinoic acid (Sigma), and 0.05% knockout serum replacement (Invitrogen). To form EBs, cells were plated at 1.25×105 cells per mL in nonadherent Petri dishes in medium consisting of DMEM (Invitrogen), 10% fetal bovine serum (Atlanta Biologicals), 50 mM HEPES (Sigma), and 1 mM β-mercaptoethanol (Sigma) for 4 days, and then transferred to gelatin-coated 12-well dishes for an additional 2 days of adherent culture in the same medium. Cells were maintained at 37°C with 5% CO2.
To test their differentiation potential in the intact animal, control and induced transgenic ESC were injected over the flank of NOD SCID-gamma (NSG) mice (The Jackson Laboratory). Geminin expression was induced for 4 days before implantation, and control cells were maintained in culture for a similar period. Cells were trypsinized, pelleted, and 1×106 cells in a volume of 250 μL injected subcutaneously. After 3–6 weeks, when obvious tumors were present, animals were sacrificed, tumors photographed, and embedded without fixation in OCT. Frozen sections were cut and stained with hematoxylin and eosin or immunohistochemical localization of cell-type-specific antibodies (described in the following section) carried out to confirm the presence of multiple lineages in the teratoma.
Plasmid construction and transfection
Geminin shRNA plasmids were described previously [16]. The H1 promoter was used to drive expression of the shRNA, while a separate eGFP cassette driven by the CMV promoter was used to identify transfected cells and monitor the transfection efficiency (Fig. 7A). ESC were passaged 12 to 24 h before transfection and plated in gelatin-coated 6-well plates (2.5×104 cells per cm2) in complete medium with LIF. ES cells were transfected with Lipofectamine Plus (Invitrogen) as directed by the manufacturer and cultured overnight in serum-free conditions (OptiMem; Invitrogen), followed by EB medium before RNA extraction, western blotting, or immunohistochemistry (IHC), as described below.
Inducible expression of Geminin in ESC
We obtained the MGZRTcH2 ESC cell line and the corresponding exchange vector from Dr. Shinji Masui [17]. This cell line contains a tetracycline-regulated transactivator, a tetracycline-response element, followed by the minimal promoter of the human cytomegalovirus (hCMV*-1) immediate early gene, a hygromycin resistance cassette, directional loxP sites, and an IRES-Venus (yellow fluorescent protein) cassette knocked-in to the endogenous ROSA26 promoter. The corresponding exchange vector replaces the hygromycin resistance cassette with the desired cDNA in addition to adding a puromycin resistance cassette. When the exchange vector is correctly enzymatically recombined into the ROSA26 locus, the resulting clones are no longer hygromycin resistant, but are puromycin resistant—allowing both positive and negative selection, thereby ensuring the elimination of cells with random integration of the exchange vector (Fig. 1A).
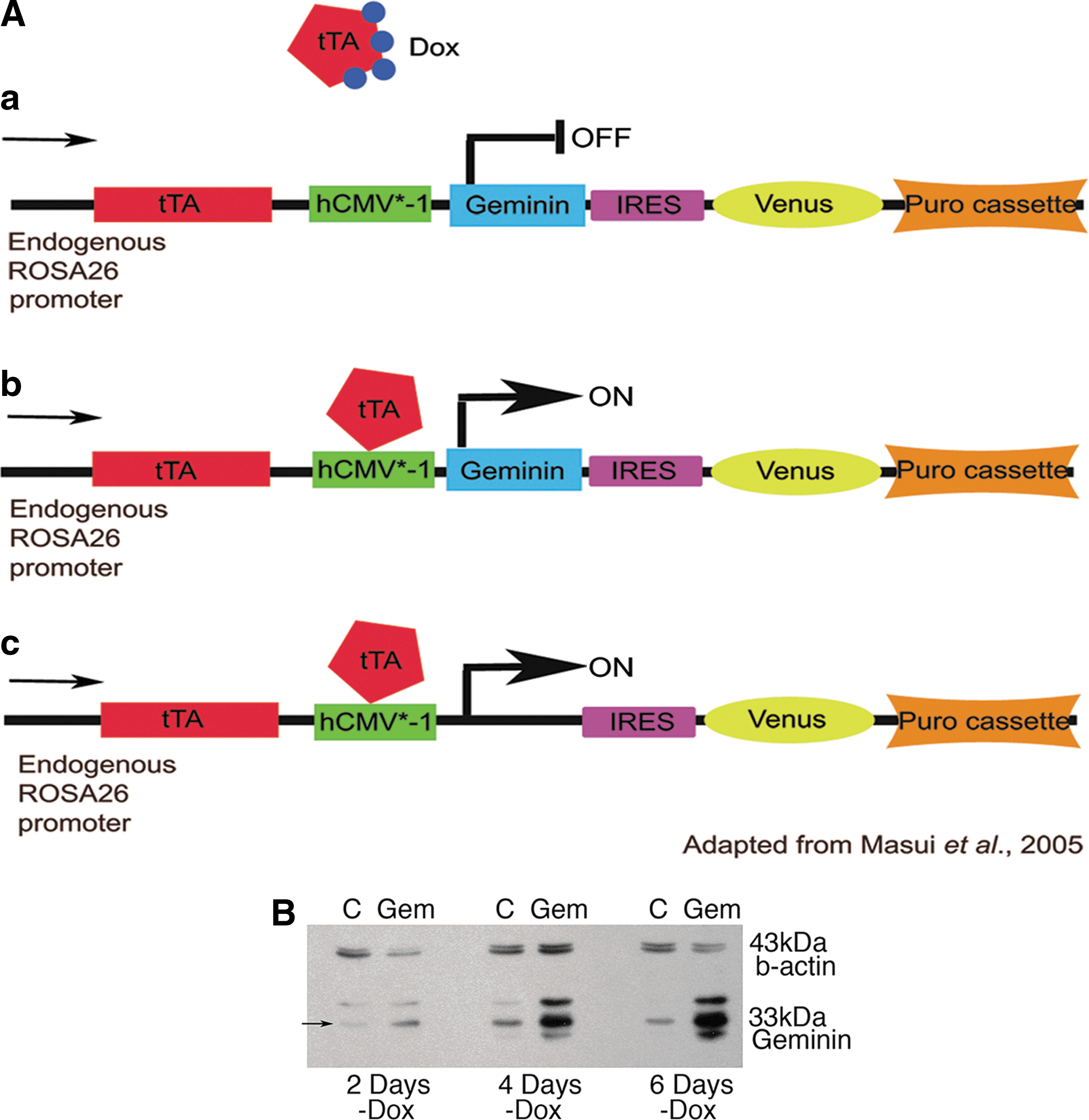
Geminin can be inducibly expressed in embryonic stem cells (ESC).
A polymerase chain reaction (PCR)-cloned and sequence-verified Geminin cDNA was inserted into one exchange vector. A control exchange vector with no cDNA was also generated to develop control cell lines that are puromycin resistant and express the Venus protein upon doxycycline (Dox) withdrawal. Each exchange vector was transfected into the MGZRTcH2 cell line with a Cre recombinase vector, followed by selection with 2 mg/mL puromycin (Sigma). Clones were expanded in complete medium with 1 mg/mL Dox (Sigma) to inhibit transgene expression and 2 mg/mL puromycin to maintain selection, and several selected to determine their differentiation capability. Clones were checked for random integration with 100 mg/mL hygromycin (Sigma). Overexpression of Geminin was verified by western blot and quantitative PCR. During all differentiation experiments, selection was maintained with 4 mg/mL puromycin, and Geminin expression was induced by Dox withdrawal (tet-off).
Cell cycle analysis
Cells were grown in EB medium for 6 days. On day 4, cells were plated at 2×105 cells/well in 6-well plates in triplicate in EB medium. After 48 and 96 h, cells were trypsinized and counted using a Coulter Counter (BD). For cell cycle analysis, samples were fixed by adding cold absolute ethanol and kept at −20°C until analysis. Fixed cells were resuspended in PBS, and DNAse-free RNase A (Sigma) was added to a 20 mg/mL final concentration. Samples were incubated at 37°C for 30 min, and DNA was labeled with 0.1 mg/mL propidium iodide (Sigma). Cells were sorted in a FACSCalibur using Cell Quest Pro MAC 9.0 and Mod Fit LT Mac 3.1 SP3 for cell cycle analysis.
Immunohistochemistry
ES cultures were fixed with 2% paraformaldehyde (PFA) for 15 min at room temperature and washed twice with PBS. Fixed cells were blocked in 10% donkey serum for 30 min, followed by an overnight incubation at 4°C with primary antibody (rabbit anti-Sox3: Mike Klymkowski, University of Colorado, 1:1,000; goat anti-Oct3/4: Santa Cruz, 1:500; rabbit anti-Foxa2: Upstate Biotechnology, 1:500; mouse anti-E-cadherin: Cell Signaling, 1:500; goat anti-Brachyury: Santa Cruz, 1:500; anti-activated β-catenin: Millipore; 1:250; or rabbit anti-phospho-histone H2A.X antibody: Cell Signaling, 1:500). Cells were then washed with PBS and incubated with secondary antibodies (1:400) conjugated to Cy3 or FITC (Jackson Immunoresearch) for 2 h at room temperature. Nuclei were visualized with Hoechst 33258 (Sigma), and images were obtained using an Leica DM inverted fluorescence microscope and an Olympus DP70 camera with associated software.
Quantitative RT-PCR
RNA was harvested with Trizol (Invitrogen) following the manufacturer's protocol, and genomic DNA was removed by DNAse digestion (Sigma). Complete DNA digestion was confirmed by semiquantitative PCR using primers for β-actin before reverse transcription. About 0.5 to 1.0 mg RNA sample was used for reverse transcription with Verso RT (Thermo Scientific) using random nonamers (Invitrogen) following the manufacturer's protocols. For quantitative PCR, cDNAs were diluted 1:2, and 1 μL was used per reaction with the ABgene SYBR green master mix. All primer pairs were rigorously screened to eliminate the primer dimer, and reaction conditions were optimized, producing reaction efficiencies between 90% and 110%. Quantitative PCR results were calculated, and statistical analysis performed with REST2008 software [18]. Primer sequences and detailed reaction conditions are available upon request.
Western blotting
ESC were lysed in an RIPA buffer (50 mM Tris–HCL, pH7.4, 1% NP-40, 0.25% Na-deoxycholate, and 150 mM NaCl2) with Complete protease inhibitor (Roche), cell debris was pelleted by centrifugation, and the protein-containing supernatant was removed and analyzed using the Pierce Protein Assay to determine total protein concentration. Ten to 30 mg of protein was loaded onto 10% or 12% polyacrylamide SDS PAGE gels and then transferred to PVDF membranes. Membranes were blocked with 5% milk powder/TBST (Tris-buffered saline and 0.2% Tween 20) and incubated with rabbit anti-Geminin (1:2,000; Santa Cruz), anti-activated β-catenin (1:2,000; Millipore), goat anti-Tcf3 (1:500; Santa Cruz), goat anti-panTLE (1:500; Santa Cruz), mouse anti-E-Cadherin (1:20,000; Cell Signaling), mouse anti-N-Cadherin (1:5,000; Life Technologies), and/or mouse anti-β-actin (1:10,000; Sigma) primary antibodies in 5% milk powder/TBST overnight at 4°C. Membranes were washed in TBST and incubated for 1 h at room temperature in a horseradish peroxidase-conjugated secondary antibody (1:1,000–1:10,000; Jackson Immunoresearch). Membranes were developed using a Pierce Supersignal West Pico chemiluminescent substrate. Western blotting was done on 3 Geminin transgenic cell lines and was repeated at least 3 times each.
Cell migration assay
To determine if cells overexpressing Geminin would more rapidly recolonize a wound, 3 lines of Geminin transgenic ESC were plated in triplicate at 5×105 cells per well in gelatin coated 6-well plates in EB medium±Dox. After 5 days of transgene induction, a cell scraper was used to remove a strip of cells from the middle of each dish, and the medium changed immediately. On 5 sequential days of culture, 5 photomicrographs (at different levels of the scrape) were taken from each of 3 replicate cultures±Dox, with 3 biological replicates (n=45 photomicrographs per timepoint) for each of the 3 lines. The distance between the cut edges was measured on each day and averaged, and significant differences were determined by matched-pair t-test.
TOP-/FOPFlash assay
Control and Geminin ESC were grown in adherent culture in EB medium for 48 h to induce transgene expression, in the presence and absence of the GSK3β inhibitor Chiron99021 (10 μM) as a positive control. Cells were transfected with TOPFlash or FOPFlash plasmids (Millipore) using Lipofectamine Plus (Invitrogen) according to the manufacturer's instructions. A Renilla plasmid (pRL-CMV; Promega) was cotransfected as an internal vector control. After 24 h, cells were lysed with passive lysis buffer and subjected to the Dual-luciferase assay (Promega) according to the manufacturer's directions. Samples were read with a Veritas microplate luminometer (Turner Biosystems). Data are expressed as the ratio of TOPFlash relative to Renilla over FOPFlash relative to Renilla, and significance was assessed using the Wilcoxon signed-rank test.
Nuclear β-catenin assay
To further interrogate the status of Wnt-signaling pathway members in Geminin-overexpressing ES cells, we determined the percentage of cells in which β-catenin was present in the nuclei. Control and 2 Geminin transgenic cell lines (G12 and G17) were grown without Dox to induce transgene expression, with Dox (negative control), or cells gown with Dox and the GSK3-β inhibitor Chiron99021 (10 μM) to stimulate the Wnt pathway (positive control). After 72h, cells were fixed in 2% PFA and stained with anti-β-catenin antibodies and with the Hoechst dye to label nuclear DNA. Quantification was carried out by counting the number of β-catenin+ nuclei divided by the total number of (Hoechst+) nuclei in 10 fields from 4 replicate cultures with 3 biological replicates (120 fields per group). Data are expressed as means of the 3 biological replicates±SD, and statistically significant differences between the treatment groups were determined using the Wilcoxon signed-rank test, followed by matched-pair tests.
Coimmunoprecipitation
Control and Geminin ESC were grown in EB medium for 4 days to induce transgene expression. Cells were scraped from the tissue culture dishes and lysed in Blenis lysis buffer (10 mM KPO4, 1 mM EDTA, pH7.0, 5 mM EGTA, pH7.2, 10 mM MgCl2, 50 mM β-glycerol phosphate, and 0.3% Chaps) with Complete protease inhibitor (Roche). Protein samples were divided in half, and 2 mL of goat anti-TLE (Santa Cruz) or rabbit anti-Geminin (Santa Cruz) antibodies were added to an aliquot of each sample set and incubated overnight at 4°C with end-over-end agitation. Preblocked Protein A or Protein G Sepharose beads (GE) were added to samples with antibody as well as to samples with input protein alone as a negative control and incubated for 1 h at 4°C with end-over-end agitation. Samples were centrifuged, and the supernatant was saved for a loading control. Sepharose beads were washed 3 times with a lysis buffer at 4°C. The loading buffer was added to the Sepharose beads, and the samples were boiled for 5 min to release the antibody/antigen complexes from the beads before analysis by western blot as described above.
Results
Geminin can be inducibly expressed in ESC
To explore the effects of increasing levels of Geminin protein in ESC, we created cell lines that express Geminin when Dox is removed from the culture medium (Tet-off; Fig. 1A). Twenty-four Geminin and 24 control (puromycin-resistant cells that inducibly express only Venus yellow fluorescent protein) ESC lines were cloned and expanded for further study. Accurate targeting was verified by testing for both hygromycin sensitivity and puromycin resistance, as well as verification that Geminin was overexpressed only in the absence of Dox. Several lines were selected for additional study based on Dox-regulated Geminin protein overexpression and/or Venus protein expression. Geminin protein levels were increased 2 days after Dox withdrawal, and by 4 days, Geminin expression was significantly elevated compared to control ESC and remained overexpressed at 6 days (Fig. 1B). These data indicate that the Geminin protein can be reliably overexpressed in response to Dox withdrawal.
Geminin promotes neural precursor and neuronal differentiation in monolayer culture
To examine their neural differentiation potential, cells were grown in defined, neural-permissive medium at low density for 6 days of transgene induction. In all 3 lines tested, compared with the same uninduced cell lines, Geminin overexpression promoted differentiation of both Sox3+ neural precursors and Tuj1-positive primitive neurons (Supplementary Fig. S1; Supplementary Data are available online at
Geminin promotes EMT and mesendodermal differentiation in EBs
To examine the role of the Geminin protein in ESC differentiation, we also employed a three-dimensional (3D) differentiation assay. Cells were plated in nonadherent Petri dishes in the absence of LIF and Dox, but with 10% serum, promoting the formation of EBs and inducing differentiation. Because some tetracycline derivatives, including Dox, inhibit matrix metalloproteinases [19], potentially reducing EMT in control cells grown in the presence of Dox, we compared 3 Geminin-inducible lines and a control line that lacked the inserted Geminin cDNA both cultured without Dox.
Most Geminin-overexpressing EBs attached to the nonadherent Petri dish 3 to 4 days after Dox withdrawal, and the cells rapidly migrated from the EB (Fig. 2). However, the vast majority of control EBs remained intact and floating for the entire 6-day assay. Therefore, to force the attachment of control EBs and promote differentiation, cells were grown as floating EBs for 4 days and then replated onto adherent tissue culture dishes for an additional 2 days before fixation for IHC or harvesting of RNA. Geminin EBs expressed strikingly less E-cadherin protein compared with control EBs (Fig. 2A) and downregulated E-cadherin at the level of transcription in quantitative PCR (Fig. 3B). We also examined E-cadherin and N-cadherin expression by western blot in 3 cell lines cultured as EBs with and without Dox for 2, 4, and 6 days (Fig. 5A), which identified a reduction in E-cadherin protein levels after 4 and 6 days of transgene induction and a concomitant upregulation of N-cadherin protein and mRNA 6 days after transgene induction.
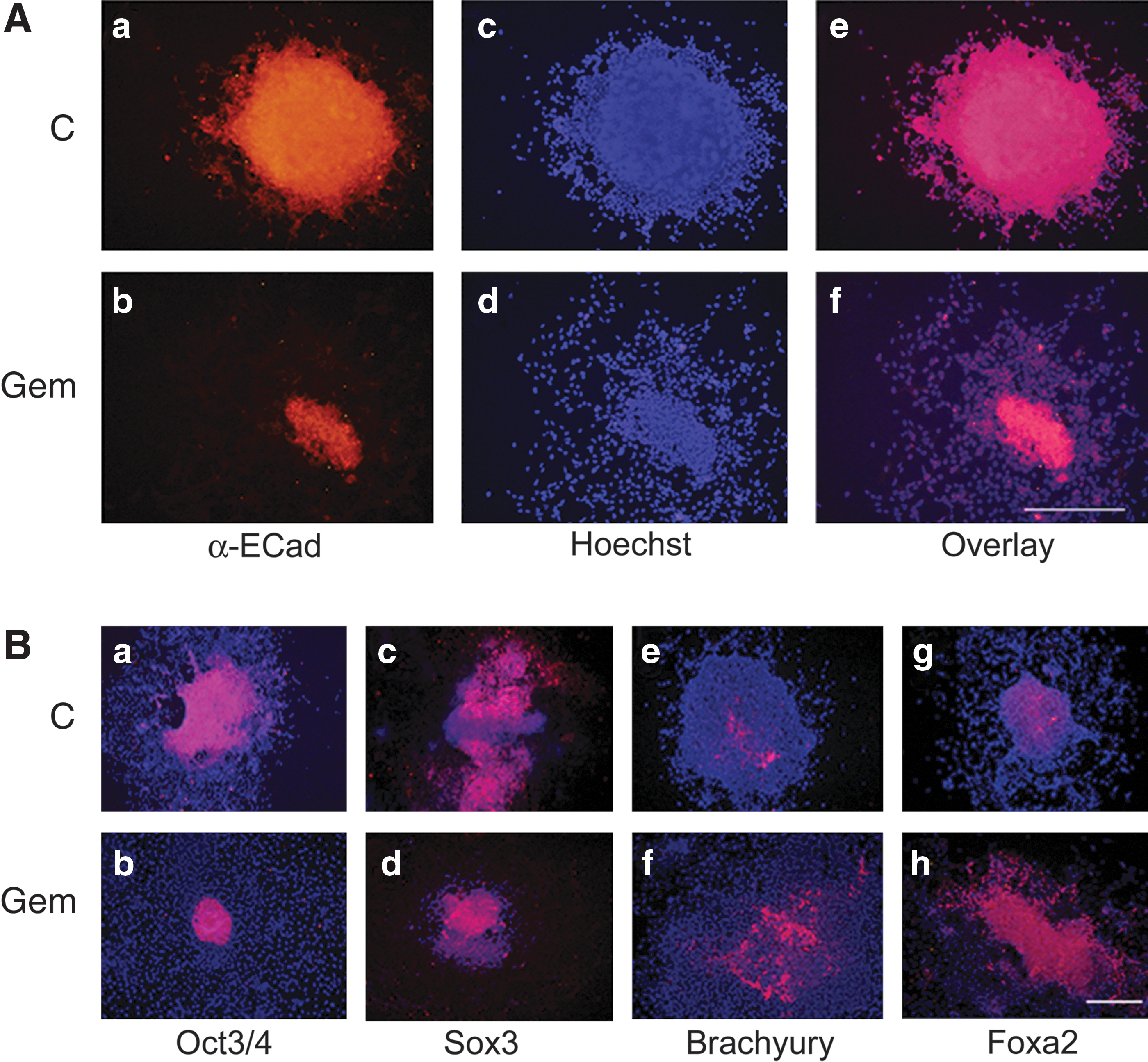
In three-dimensional culture, Geminin promotes epithelial-to-mesenchymal transition (EMT) and mesendodermal differentiation.
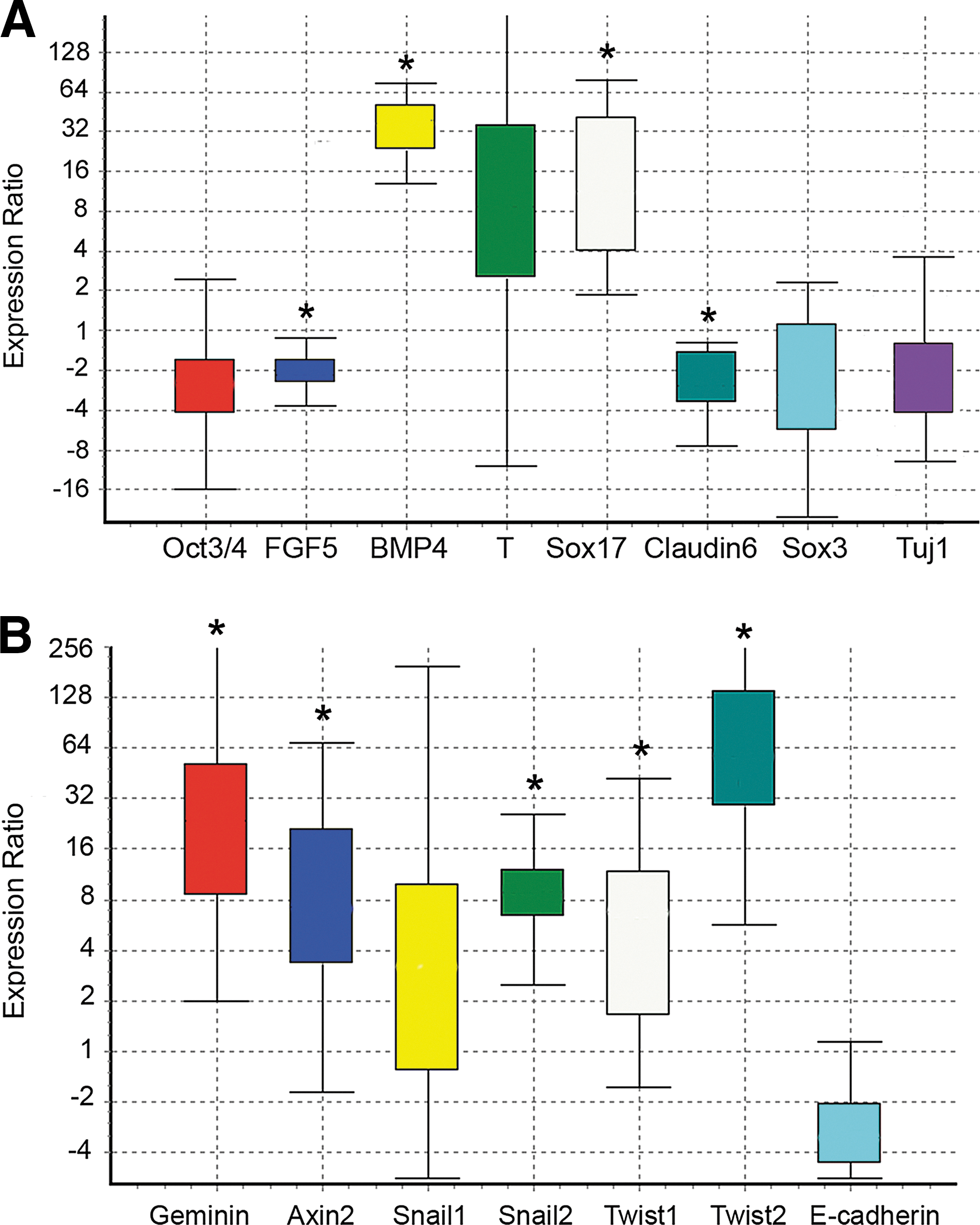
Analysis of gene expression in Geminin versus control EBs. Quantitative PCR of mRNAs from EBs grown for 6 days as described in Fig. 2A. Box-and-whisker plots were produced with REST software [18]. The top and bottom whiskers indicate the range, while the box indicates the upper- and lower-quartile values. Gene expression was calculated relative to β-actin and set to 1 in the control ESC line. Asterisks indicate statistically significant differences (Geminin vs. control, P≤0.05) calculated by the REST software (n=3 replicates).
When the expression of lineage-restricted markers was examined by IHC (Fig. 2B), compared with controls, Geminin overexpression decreased the expression of the pluripotent stem cell marker Oct3/4 and of the neural precursor gene Sox3, while the mesodermal marker Brachyury and the endoderm-restricted protein Foxa2 increased (Fig. 2B). We also examined the expression of lineage-restricted markers (Fig. 3A) and genes involved in EMT (Fig. 3B) in quantitative PCR. Geminin overexpression decreased Oct3/4 and the epiblast marker FGF5, indicating that the Geminin-overexpressing cells differentiated more rapidly than control ESC. Markers of the mesendodermal lineage, BMP4, Brachyury, and Sox17, were significantly increased, whereas the epidermal ectoderm marker Claudin6, Sox3, and the neuronal differentiation marker Tuj1 were decreased. Expression of genes involved in EMT, Snail1, Snail2, Twist1, and Twist2, was also increased in Geminin-overexpressing cells, and E-cadherin downregulated. Taken together, these data indicate that Geminin controls the expression of genes involved in EMT.
A wounding assay was used (Fig. 4) to characterize the highly migratory phenotype resulting from Geminin overexpression. Geminin-overexpressing cells (n=3 transgenic lines±Dox) migrated rapidly from the edge of a scraped monolayer; after 4 days, the denuded region had been completely recolonized by Geminin-overexpressing ESC, while in uninduced control cultures after 5 days, a mean gap of 178.3±7.1 μm remained (Fig. 5). This is consistent with our observations that Geminin promotes rapid EMT both in EBs and in the early embryo [20], and with the induction of genes involved in cell migration in Geminin-overexpressing ESC (Fig. 3B).
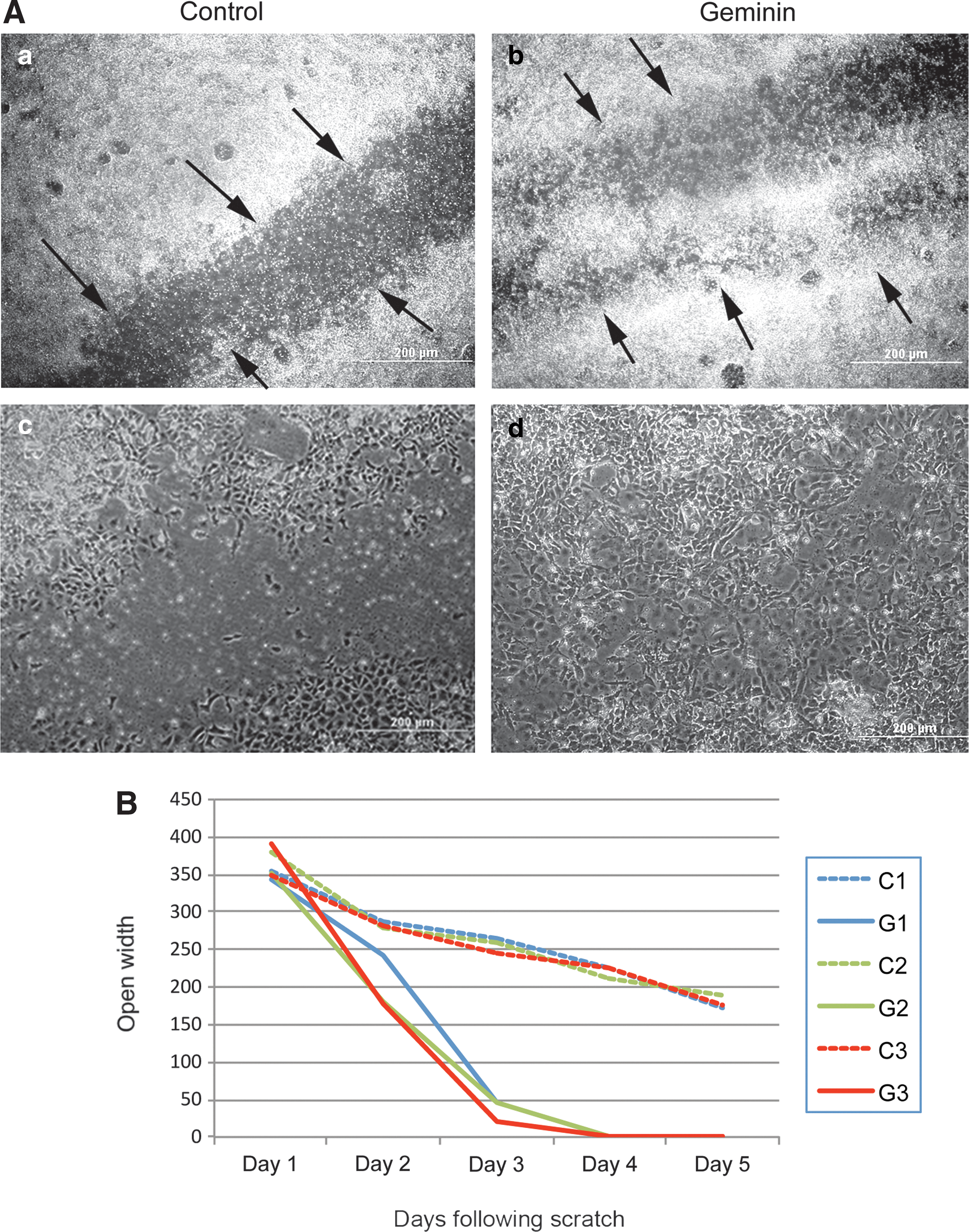
Geminin promotes rapid recolonization of a scrape wound.
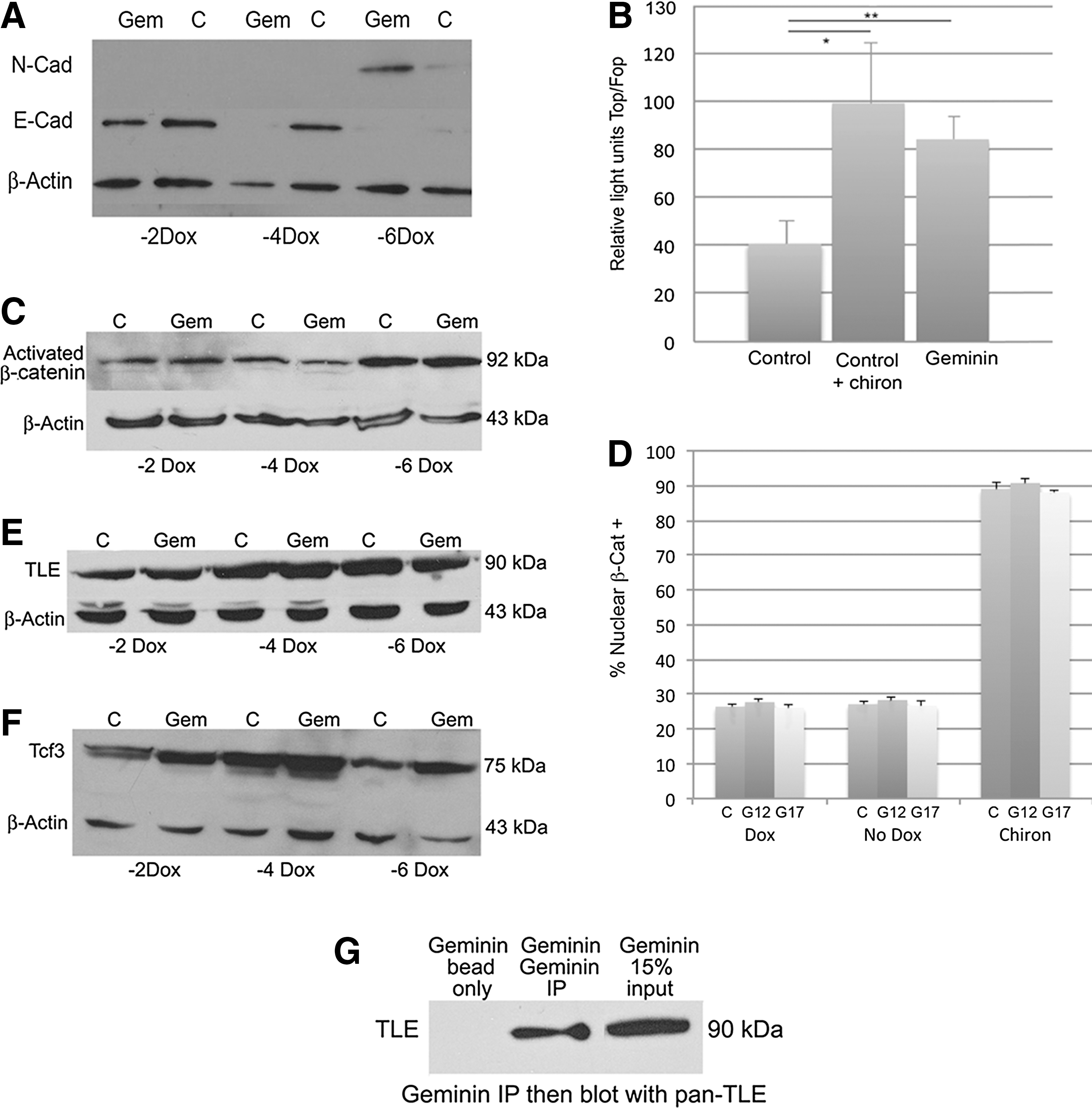
Geminin overexpression stimulates Wnt signaling.
Geminin does not affect trilineage differentiation in teratomas
Geminin overexpression increased the rate at which teratomas formed in NSG mice, but all the 3 germ lineages were present in the tumors. Of the 6 subcutaneous injections, all formed teratomas >1 cm in diameter after 3 weeks, while injection of uninduced ESC in 4 flanks produced similar-sized tumors after 4–6 weeks in vivo. Histological analysis of the teratomas indicated that the Geminin-overexpressing teratomas contained derivatives of all 3 germ layers, but also contained undifferentiated Oct3/4-positive cells, and did not exhibit the complex tissue differentiation characteristic of control teratomas, possibly because of the short interval before the animals were sacrificed (data not shown). Teratomas developed from control ESC contained a complex mixture of endoderm, mesoderm (muscle and cartilage), and neuronal cells (data not shown). The tet-off system is particularly useful in transplantation paradigms, as it insures continued expression of the transgene in vivo, unlike constructs that require the continued presence of an inducing agent.
Geminin overexpression stimulates Wnt signaling in ESC
Because Wnt signaling plays a critical role in formation of the primitive streak and mesendodermal differentiation in the early mouse embryo, we examined the status of Wnt pathway signaling in Geminin-overexpressing ESC. Induction of Geminin protein expression significantly increased canonical Wnt signaling in the TopFlash luciferase assay compared to controls (Fig. 5B, Control vs. Geminin, **P≤0.01), which was approximately equal to the increase in Wnt signaling produced by exposure of control cells to the Gsk3β inhibitor Chir99021 (Control+chiron vs. Geminin, P≤0.70, and Control vs. Control+chiron *P≤0.02), previously shown to promote Wnt signaling [21]. In addition, a well-characterized canonical Wnt target gene, Axin2 [22 –25], was upregulated over 7-fold after Dox withdrawal in an EB culture (Fig. 3B). Geminin ES cells failed to activate the TopFlash reporter, and Axin2 gene expression was unchanged in the presence of Dox (data not shown). These data indicate that in ESC, Geminin overexpression functions to stimulate Wnt signaling as was observed in Wnt indicator mice [20]. Since BMP signaling is also required for mesendodermal differentiation in the mouse embryo, we examined BMP4 and Axin2 expression after 3 days of EB differentiation by quantitative PCR. Axin2 was increased 57-fold, while BMP4 was only increased 3-fold, suggesting that Geminin promotes Wnt signaling that may in turn upregulate BMP4 expression.
To examine the mechanisms underlying Geminin's ability to stimulate Wnt signaling, we examined the percentage of cells in which β-catenin was present in the nucleus, indicating pathway activation. In the presence of Dox, 26.4%±0.7% of Control, 27.7%±1.4% of Geminin line12, and 26.1%±0.7% Geminin line17 cells exhibited nuclear β-catenin. The pathway agonist chiron strongly stimulated nuclear localization of β-catenin: 89.0%±3.0% of Control, 90.7%±4.0% of G12, and 88±0.6% of the G17 cells exhibited nuclear β-catenin. When grown in the absence of Dox to activate transgene expression, there were baseline levels of nuclear β-catenin in both Control cells (27.1%±0.9%) and in Geminin-overexpressing cells (28.3%±1.1%; Line G12, and 26.7%±1.9%; Line G17) (Fig. 5D). Differences between the means were not statistically significant. These data suggest that Geminin stimulates Wnt signaling via a mechanism that is unlikely to involve β-catenin.
Geminin may induce Wnt signaling by binding TLE proteins
To begin to identify the mechanisms by which Geminin promotes Wnt signaling, we first examined the expression of the Wnt pathway members. Geminin did not dramatically increase the level of activated β-catenin protein (Fig. 5C) or affect its localization (Fig. 5D), suggesting that Geminin does not regulate expression of Wnt ligands or inhibit GSK3β. Expression of TLE proteins, which are required for Wnt pathway inhibition, was largely unchanged by Geminin overexpression (Fig. 5E). There was a small, but consistent, increase in expression of Tcf3 (Fig. 5F), which typically inhibits the Wnt pathway, in response to increased Geminin. While this may seem inconsistent with the observed increase in the Wnt pathway activity, it is likely a negative feedback mechanism.
Since Geminin has been reported to bind proteins in the nucleus and inhibit their function, we performed coimmunoprecipitation experiments with Tcf3 and Geminin antibodies, but were unable to detect an interaction (data not shown). However, there was a weak, but consistent, interaction of Geminin with TLE proteins. Based on this interaction, we hypothesize that Geminin may bind and inhibit TLE proteins from repressing Wnt signaling in the nucleus, thereby activating Wnt target gene expression.
Geminin reduces proliferation and alters cell cycle of ESC
Since Geminin inhibits DNA replication to prevent endoreduplication, we assayed proliferation and cell cycle characteristics of Geminin and control ESC in monolayer culture. In the presence of Dox, Geminin cell lines grow similarly to the control cell line (data not shown). However, overexpression of Geminin significantly reduced proliferation (Fig. 6A), increased the length of time the cells spent in the G2/M phase of the cell cycle, and reduced the time spent in the G1 phase (Fig. 6B). Decreased proliferation may itself promote differentiation, since exit from the cell cycle is required for many cell fate decisions. Additionally, cells may be unable to progress from G2/M, because they are unable to initiate new rounds of DNA replication, since Geminin is normally destroyed by the proteasome during the M phase. Clearly, Geminin plays an important role in controlling cell cycle and proliferation in ESC similar to its role in the inner cell mass.
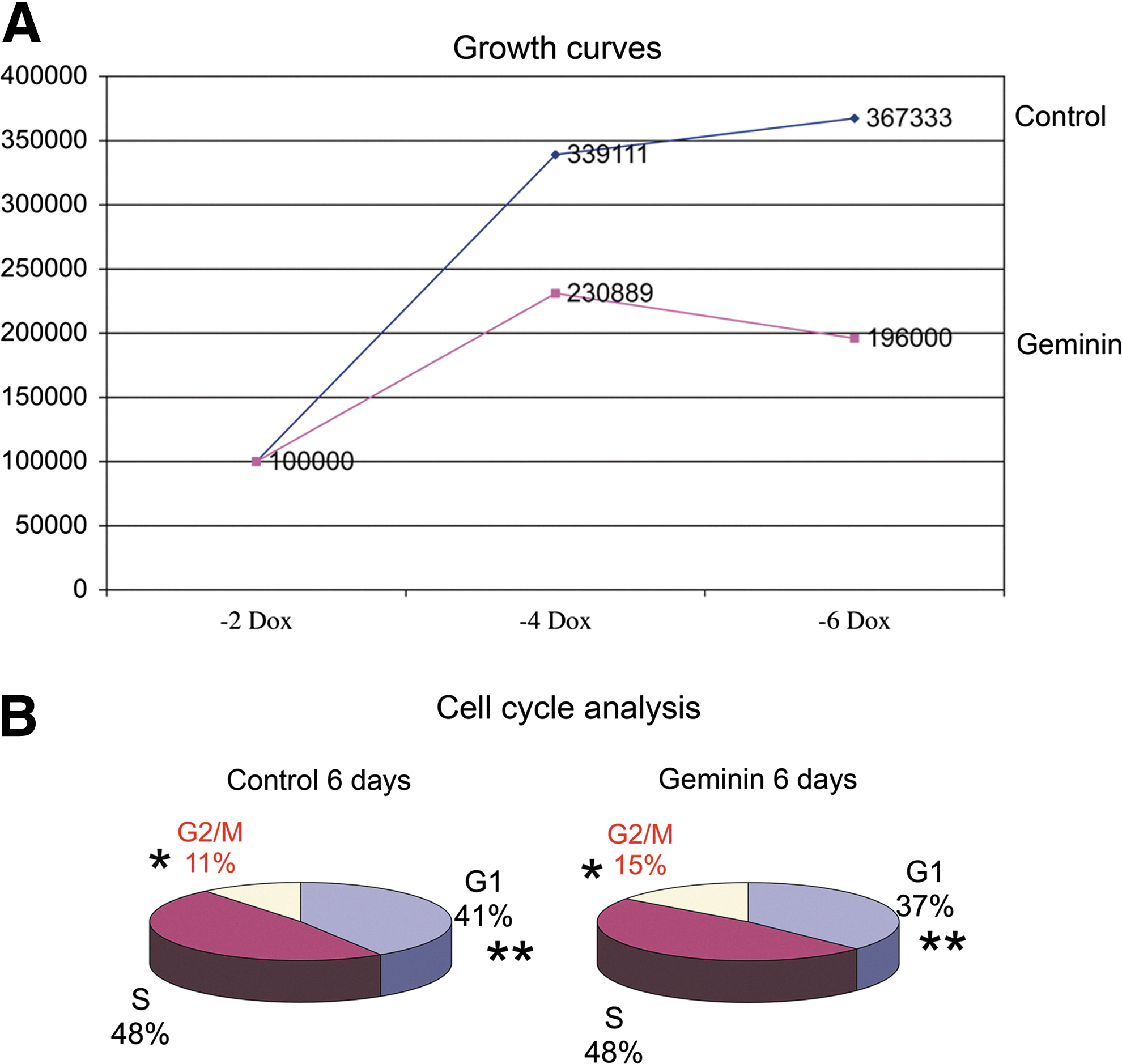
Geminin overexpression decreases proliferation and increases the length of the G2/M phase.
Geminin shRNA results in DNA damage and cell death
To determine if Geminin protein is necessary for Wnt signaling, lineage differentiation, and/or EMT, we employed shRNAs (Fig. 7A) to reduce Geminin protein levels. We were able to achieve a 6-fold knockdown of Geminin protein 24 h post-transfection (Fig. 7B); however, cells transfected with the Geminin shRNA exhibited increased DNA damage as illustrated by phosphohistone H2A.X antibody staining (Fig. 7C), precluding the analysis of differentiation and Wnt signaling. Cells transfected with the control shRNA showed some DNA damage (11.4%±0.6%) that increased dramatically when Geminin was knocked down (84.3%±0.1%), P≤0.001.
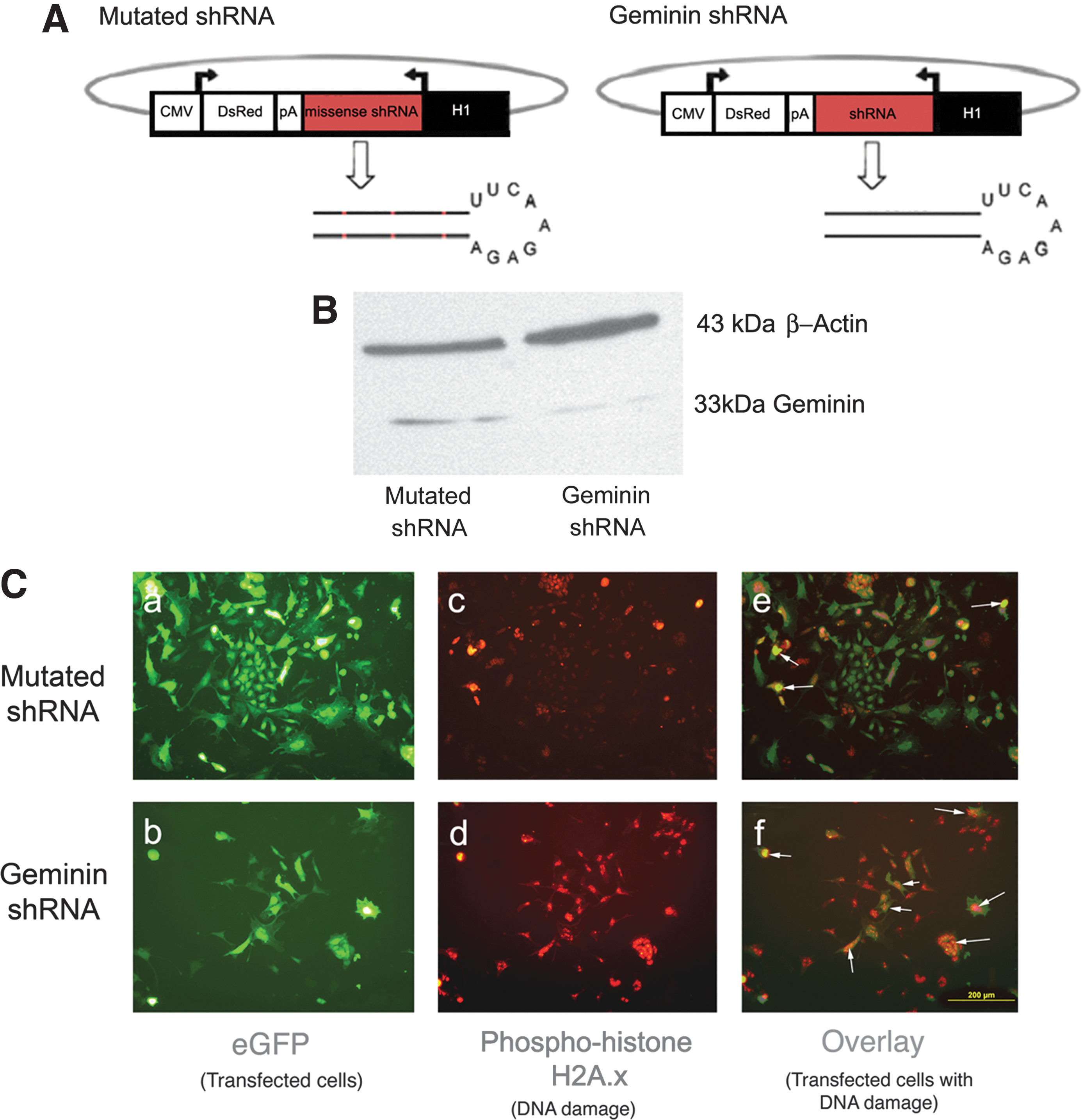
Transfection of ESC with a Geminin short hairpin RNA (shRNA) plasmid causes DNA damage.
Discussion
Geminin has previously been characterized by its ability to promote neural differentiation at the expense of alternative lineages in Xenopus [8,14] and Drosophila embryos [26]. Our results demonstrate that the outcome of Geminin overexpression in ESC is highly context dependent: in a sparse monolayer culture, in defined medium, it can promote neural differentiation, while in 3D culture, as EBs with serum, it promotes EMT and mesendodermal differentiation.
Given its context-dependent effects, it is not surprising that 2 recent reports of Geminin gain and loss of function in ESC come to remarkably different conclusions. The first proposed that Geminin escapes degradation in ESC and inhibits the action of Brg1 to sustain Oct4, Sox2, and Nanog expression, thereby maintaining pluripotency [27]. The second study reported that Geminin controls neuronal differentiation via regulation of chromatin, although without corresponding alterations in protein expression or in promoting differentiation [28]. Low levels of overexpression and downregulation obtained in that study may have created a state of chromatin readiness to respond to lineage differentiation factors, rather than neuronal differentiation per se. Recently, targeted deletion of Geminin in neural stem cells suggested that in vivo Geminin may have a primary role in cycle regulation rather than in lineage choice [29], although another analysis demonstrated that deletion expanded the neural precursor population, and overexpression promoted migration and premature neuronal differentiation [30].
Our overexpression studies consistently demonstrate that Geminin promotes, rather than inhibits, differentiation in EBs; however, it remains possible that the outcome is dependent on the level of expression or on the differentiation state of the cell—producing different outcomes in ESC versus lineage-committed neural precursors. Additionally, we were unable to assess ESC differentiation in the absence of Geminin protein, as we observed DNA damage in shRNA transfected cells, likely not observed using siRNAs, since Geminin is degraded and resynthesized during each cycle.
Geminin has been shown to bind and inhibit the function of several nuclear factors, including AP4 [31], the chromatin-remodeling protein Brg-1 [32], Hox proteins [33], and Six3 [34]. Geminin also has the potential to act as a molecular switch between proliferation and differentiation by binding additional transcription factors [35], including TLE proteins. Tcf/Lef transcription factors inhibit Wnt signaling in the absence of nuclear β-catenin [36,37], and our results suggest that Geminin may inhibit interactions between Tcf/Lefs and TLEs to de-repress Wnt signaling.
Wnt signaling is critically involved in establishing the primitive streak and promoting the EMT required for mesendodermal differentiation in the embryo [38 –44]. The transcriptional repressors Snail1 and Snail2 both downregulate the expression of E-cadherin, and in the absence of Snail1, mesendoderm is still specified; however, gastrulation fails because cells are unable to leave the primitive streak [44,45]. It has been hypothesized that downregulation of E-cadherin releases β-catenin from cytoskeletal junctional complexes, increasing Wnt signaling [46,47] that in turn feeds back to increase Snail1 [48 –52]. However, it appears that in our EB model, Geminin de-represses Wnt signaling to promote Snail1 and Snail2 expression, suggesting that Geminin should now be included in the list of genes that initiate EMT and mesendodermal differentiation.
Wnt signaling is required for the proliferation of neural precursors [53 –56], for neuronal differentiation [57,58], and patterning of the developing nervous system [59,60]. The key bHLH transcription factors that drive neuronal specification, including Neurog1 and NeuroD1, are transcriptional targets of the canonical Wnt pathway [54,57,61]. Therefore, while Wnt signal inhibition may initially be required to promote neural precursor differentiation by inhibiting mesendodermal lineage differentiation, active signaling appears to be required for precursor proliferation and neuronal differentiation [62].
Wnt signaling has previously been shown to be required for the differentiation of ESC to mesendoderm [63 –65] and is also implicated in the switch from the pluripotent to the differentiated state via Tcf3 binding to a protein complex with Oct4, Sox2, and Nanog on gene promoters [66 –68]. In many cell types, cell fate decisions are tightly coupled to the cell cycle exit [36,69,70], and the most crucial decision in the cell cycle, that is, to proliferate, differentiate, quiesce, senesce, or apoptose, occurs at the G1 checkpoint [71]. The attenuated G1 phase in adult stem cells, ESC, and the epiblast may prevent differentiation by insulating cells from growth factor exposure [72 –74]. Overexpression of Geminin decreased proliferation and increased the length of G2/M in both ESC (above) and in early postimplantation embryos [20]. Persistent Geminin protein expression without cyclic degradation is likely to maintain the sequestration of Cdt1 and block progression from the M phase. In turn, lengthening G2/M would increase the time Geminin is available to interact with other proteins in the nucleus and promote differentiation. Our results suggest that Geminin, via its ability to de-repress Wnt signaling, may be a key regulatory factor controlling cell cycle, EMT, and differentiation at gastrulation.
Geminin expression has been negatively correlated with patient prognosis in several cancers [10,75,76], where it may promote EMT and therefore metastasis. In fact, when Geminin was overexpressed in the MCF-10 nontumorigenic mammary epithelial cell line, there was increased proliferation and anchorage-independent growth [77,78]. Knockdown of Geminin protein might therefore be a powerful approach to both inhibit metastasis and initiate endoreduplication and apoptotic cell death in dividing tumor cells.
Footnotes
Acknowledgments
The authors are great to Brian Magnuson and Diane Fingar for help with western blotting and immunoprecipitation, Maria Morell for cell cycle analysis and DNA damage assessment, Lisa S.D. Emmett for developing ESC lines, Yao-Chang Tsan for thoughtful discussions, Mike Klymkowsky for Sox3 antibody, and Yuji Masui for cell lines and plasmids, the Flow Core at the University of Michigan; supported by the NIH grants RR-023187 and NS-39438.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.