
Abstract
Multipotent skin-derived progenitors (SKPs) can be traced back to embryonic neural crest cells and are able to differentiate into both neural and mesodermal progeny in vitro. Neural stem cells (NSCs) are capable of self-renewing and can contribute to neuron and glia in the nervous system. Recently, we derived porcine SKPs and NSCs from the same enhanced green fluorescent protein (EGFP) transgenic fetuses and demonstrated that SKPs could contribute to neural and mesodermal lineages in vivo. However, it remains unclear whether porcine SKPs and NSCs can generate ectoderm and mesoderm lineages or other germ layers in vivo. Embryonic chimeras are a well-established tool for investigating cell lineage determination and cell potency through normal embryonic development. Thus, the purpose of this study was to investigate the in vivo developmental potential of porcine SKPs and fetal brain-derived NSCs by chimera production. Porcine SKPs, NSCs, and fibroblasts were injected into precompact in vitro fertilized embryos (IVF) and then transferred into corresponding surrogates 24 h postinjection. We found that porcine SKPs could incorporate into the early embryos and contribute to various somatic tissues of the 3 germ layers in postnatal chimera, and especially have an endodermal potency. However, this developmental potential is compromised when they differentiate into fibroblasts. In addition, porcine NSCs fail to incorporate into host embryos and contribute to chimeric piglets. Therefore, neural crest-derived SKPs may represent a more primitive state than their counterpart neural stem cells in terms of their contributions to multiple cell lineages.
Introduction
N
Embryonic chimeras are a well-established tool for investigating cell lineage determination and cell potency through normal embryonic development [8]. Mouse pluripotent stem cells, such as embryonic stem (ES) cells [9], embryonic germ [10] cells, and induced pluripotent stem (iPS) cells [11], can contribute to postnatal chimeras and germline formation. In rats, germline-competent ES cells have recently been derived in a medium supplemented with small molecules specifically inhibiting GSK3, MEK, and FGF receptor tyrosine kinases [12]. Nevertheless, in nonhuman primates, such as rhesus monkeys, ES cells fail to incorporate into host blastocysts and develop into postnatal chimeras [13]. In pigs, chimeras have been produced by injecting blastomeres [14], inner cell mass (ICM) cells [15], primordial germ cells [16], putative ES cells [17], and even iPS cells [18] into blastocysts. Although most of these cells can contribute to the somatic cell lineages of term offspring, only porcine ICM and iPS cells are competent for germline transmission. Recently, we have recovered 2 chimeric fetuses from porcine SKP cells and found that EGFP transgenic SKP cells can be tracked to neural and mesodermal lineages in vivo [6]. However, it remains unclear whether SKP cells can contribute to an endodermal lineage and have competence for germline transmission. Moreover, the plasticity of NSCs in terms of transdifferentiation is still controversial [19,20]. Thus, the purpose of this study was to investigate the in vivo developmental potential of porcine SKPs and NSCs by injecting them into early stage embryos. We also injected SKP-derived fibroblasts as a control in this study. We found that porcine SKPs, but not NSCs or SKP-derived fibroblasts, incorporate into the host embryos and contribute to the derivatives of 3 germ layers in chimeric piglets, albeit at very low levels.
Materials and Methods
Animal use and care have been reviewed and approved by the Animal Care and Use Committee (ACUC) at the University of Missouri-Columbia.
Cell cultures
Unless indicated otherwise, the media and components used in this study were purchased from Sigma (St. Louis, MO).
Porcine SKPs were derived from the back skin of day 45–50 heterozygous pCAG-EGFP transgenic fetuses [21] and cultured in a serum-free medium that included DMEM/F12 (1:1; Invitrogen, Carlsbad, CA), B27 supplement (50×; Invitrogen), 20 ng/mL EGF, and 40 ng/mL bFGF. The SKP cells were kept as spheres in suspension culture plates (Sarstedt, Newton, NC) [22]. The porcine NSCs were isolated from the brain of the same fetuses where the SKPs cells had been isolated as previously described [5]. Neurospheres appeared after 5–7 days of culture in a serum-free medium as just mentioned. Porcine SKPs and NSCs were harvested by centrifuge and stored in liquid nitrogen as small spheres. The fibroblasts were induced from porcine SKP cells by switching to a induction medium that contained DMEM (high glucose; Invitrogen) and 15% fetal bovine serum (Hyclone, Logan, UT) and were cultured on an adhesion plate (Corning Incorporated, Corning, NY). After 3 days of culture, the fibroblast cells were harvested by 0.05% trypsin/EDTA, neutralized by an induction medium, and then stored in liquid nitrogen [4]. The sex of each cell line was identified through polymerase chain reaction (PCR) screening by using SRY-specific primers (Table 1).
Embryo injection and embryo transfer
Porcine in vitro fertilized (IVF) embryos were produced from in vitro matured oocytes purchased from ART LLC (Madison, WI). After fertilization, the IVF embryos were cultured in PZM3 medium for 4 days at 38.5°C in 5% CO2 in air. Both 4-cell and 8-cell stage (precompact) embryos were selected for the injection. Three types of cells were used as donor cells: SKPs, NSCs, and SKP-derived fibroblasts. Only male cell lines were chosen for the injection, as the resulting chimeras tend to be male when male and female embryos are paired. The SKPs were dispersed by using Accumax solution into single cells or small clusters. For fibroblasts, 10–15 individual cells were injected into the host precompact embryos. For SKPs, 10–15 individual cells or a small cluster with less than 20 cells was injected. However, for NSCs, 1 or 2 small spheres were injected. The injected embryos were then cultured in PZM3 medium overnight at 38.5°C in 5% CO2 in air. The next day, approximately 30–40 early blastocysts (preferred if enough) and morulae were transferred to the oviduct of a surrogate on day 4 of her estrous cycle. The putative chimeric fetuses were recovered on day 40–50 of gestation, and piglets were collected by either a Cesarean section or a natural vaginal delivery.
Tissue collection and genomic analysis
For each piglet, earnotches were collected for fibroblast cell cultures, while tail cuts were collected for genomic DNA extraction. In addition, various tissues (lung, heart, liver, spleen, kidney, brain, skin, muscle, intestine, ovary, pancreas, and stomach) were surgically collected. A portion of them were snap frozen by liquid nitrogen and then stored at −80°C for subsequent genomic analysis. The other portion of the tissue was fixed and subjected to immunohistochemistry (IHC) staining. Genomic DNA was isolated by using a DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA) following the manufacturer's instructions. The PCR was performed by using the GoTaq Green Master Mix (Promega, Madison, WI) and visualized after 1% agarose gel electrophoresis. The PCR program was 94°C for 5 min, 94°C for 30 s (denaturation), 58°C for 30 s (annealing), and 72°C for 45s (extension), followed by 45 cycles.
Immunohistochemistry
Fetal and piglet tissues were harvested and fixed with 4% paraformaldehyde (Electron Microscopy Sciences, Hatfield, PA) for 12–18 h at 4°C. Then, they were serially dehydrated by 30% ethanol, 50% ethanol, and 70% ethanol, each for 30 min, and repeating once. The dehydrated tissues were stored in 70% ethanol at 4°C until further processing. Samples were embedded in paraffin, cut at 4 μm, placed on plus-charged slides, and left flat on a 43°C slide warmer overnight. Then, the slides were heated until the paraffin was melted and hydrated through graded alcohols to water. Steaming was done in a Decloaking chamber (Biocare Medical, Concord, CA) at 98°C for 30 min, then cooled to room temperature (RT) for 10 min, rinsed with distilled water, and then placed on an intelliPATH FLX Autostainer for staining. Slides were treated with 3% H2O2 for 15 min, washed in buffer, and then treated with Sniper block (Biocare Medical) for 20 min. They were then incubated in primary anti-GFP antibody (Abcam, Cambridge, MA) for 1 h. Negative controls were treated with rabbit immunoglobulin G (IgG) (Sigma) at a 1:1,000 dilution for 1 h. Then, the slides were incubated with MACH 2 HPR-polymer secondary antibody (Biocare Medical) for 30 min with a Tris buffer rinse. Romulin Red (Biocare Medical) was used as chromogen for 10 min. The slides were then counterstained in CAT hematoxylin for 5 min, washed, dehydrated, and coverslipped.
Immunocytochemistry
Cell cultures were fixed by 4% paraformaldehyde for 10–15 min at RT and then permeabilized with 0.2% Triton X-100 in phosphate-buffered saline (PBS) for 20 min followed by washing thrice in PBST (0.05% Tween-20 in PBS). The fixed cells were blocked with blocking solution (1% bovine serum albumin in PBST) for 1 h at RT and subsequently incubated with primary antibody (1:500, diluted in blocking solution) overnight at 4°C. After washing thrice with blocking solution, they were incubated with secondary antibody (1:500, diluted in blocking solution) for 1 h at RT followed by 3 washes with blocking solution. Finally, they were incubated with Hoechst 33342 (in PBST) for 15 min. Then, the slides were covered by coverslips with a drop of Fluoromount-G mounting medium (SouthernBiotech, Birmingham, AL) and sealed with nail polish. Primary antibodies were rabbit polyclonal anti-GFP antibodiespurchased from both Invitrogen (Catalog No. A11122) and Abcam (Catalog No. ab290). Secondary antibodies wereAlexa Fluor® 594 goat anti-rabbit IgG (H+L) (Invitrogen). Parallel cultures with only secondary antibody or rabbit IgG were employed as negative controls. The images were captured by a DS Camera Control Unit DS-U2 (Nikon, Melville, NY) and processed by the NIS-Elements imaging software (Nikon).
Results and Discussion
The incorporation of porcine SKPs into early-stage embryos
The overall procedure for chimera production is summarized in Fig. 1. We used an EGFP transgene driven by a CAG promoter that tracked the donor cells in the host embryos and recovered fetuses and piglets. Here, we only selected precompact stage embryos (4-cell and 8-cell), as we hypothesized that they had a better chance to incorporate the donor cells through the process of early embryonic compaction. Although embryonic day (E) 3.5 blastocysts are routinely employed as host embryos for chimera production in mice, they may not be suitable for other species. For example, rhesus monkey ES cells failed to integrate into host blastocysts postinjection. In contrast, chimeras were produced by the aggregation of 4-cell stage embryos in rhesus monkeys [13]. In this study, individual SKP cells and fibroblastswere injected and placed between blastomeres of a 4-cell or 8-cell stage embryo, and cultured for another 2 days in PZM3 medium until the blastocyst stage. Then, they were fixed and stained with Hoechst 33342 to visualize the nuclei. Interestingly, in 45.5% of the injected embryos (n=22), the SKP cells dispersed into different locations of the host blastocysts; whereas in the remaining embryos, they were maintained as a cluster of cells within the blastocysts (Fig. 2A).The dispersion of SKP cells indicates their incorporation into host blastocysts, possibly through the process of embryonic compaction and the formation of the blastocoel. The interactions between blastomeres during lineage segregation and during cavity formation in blastocysts may trigger the dispersion of donor cells outside the host blastomeres. However, it is still mysterious why some SKP cells dispersed while others did not. It is speculated that some SKP cells might have undergone differentiation when they were dissociated into single cells; thus, they could not incorporate into host embryos. However, the fibroblast cells that were differentiated from SKP spheres did not spread around the host blastocysts (Fig. 2B). In contrast, they stayed as a clump of cells residing inside the embryos that could be identified by the green fluorescence. Thus, the differentiation of SKPs results in the loss of the ability to incorporate into host embryos on an injection.

Schematic overview of the chimera production. Transgenic (EGFP) donor cells (SKPs, NSCs, and fibroblasts) were injected into 4-cell or 8-cell IVF porcine embryos and then transferred into a surrogate at a synchronizedstage of the estrous cycle 24 h postinjection. The putative chimeric fetuses were recovered in the middle of the gestation, and piglets were collected by either a Cesarean section or a natural vaginal delivery. A large number of tissue samples were collected, fixed, and subjected to immunohistochemistry (IHC). Genomic DNA was also extracted from a portion of collected tissues and subjected to genomic analysis. Note: not real chimeras, for illustration only. SKP, skin-derived progenitor; EGFP, enhanced green fluorescent protein; IVF, in vitro fertilized.
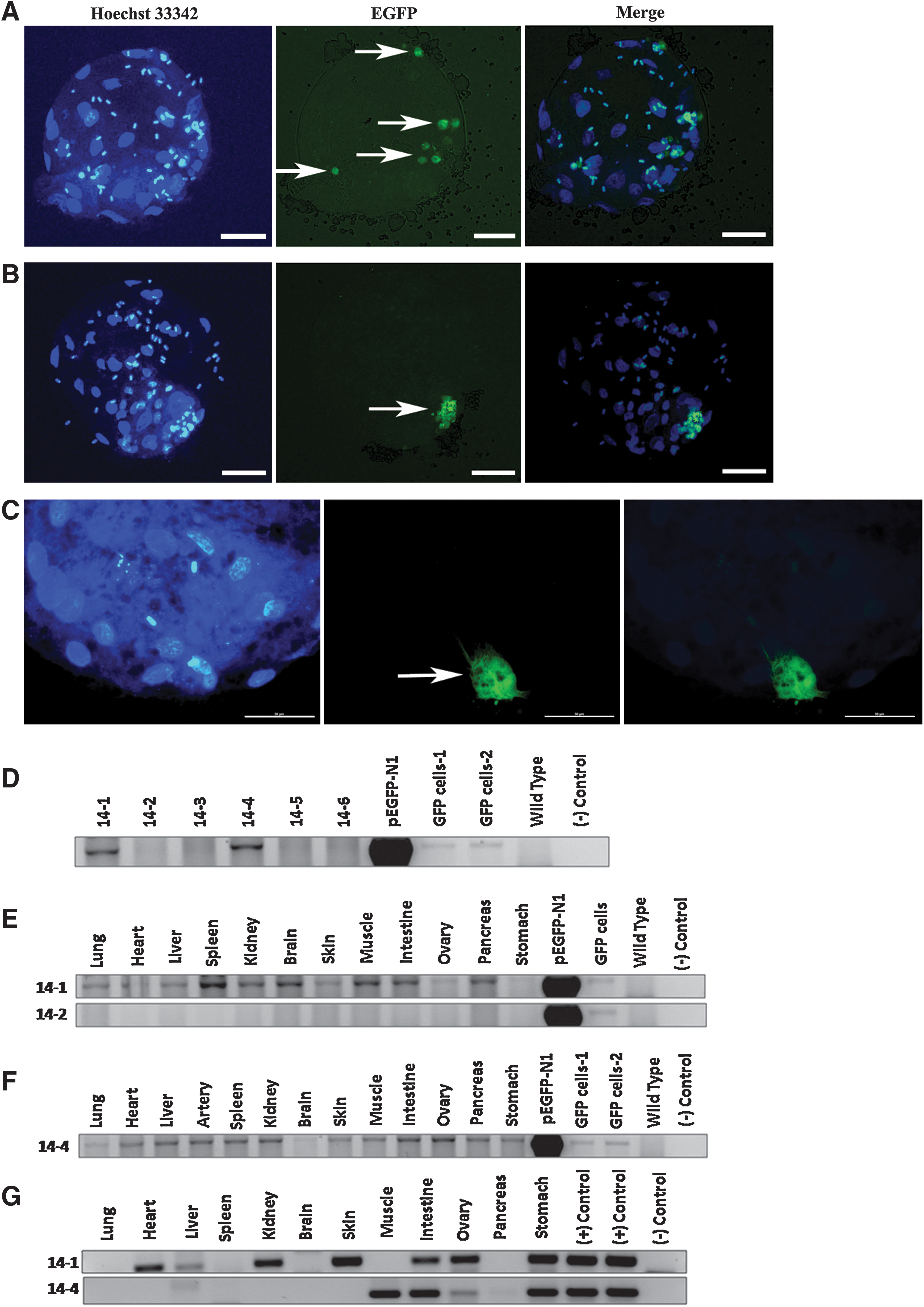
The in vivo developmental potential of porcine SKPs and NSCs. Some of the injected SKPs migrated and dispersed into different locations of the host blastocysts
In addition, NSCs also appeared to cluster together 2 days postinjection, and there was no sign of migration (Fig. 2C). We did not dissociate them into single cells, because it would induce the differentiation process within the spheres. Apparently, the PZM3 medium is not favored by NSCs, as it lacks some essential supplements. These results suggest that NSCs do not incorporate into the host embryos postinjection.
The contribution of porcine SKPs in chimeric tissues
To determine the contribution of porcine SKPs and NSCs to the postnatal chimeras, the injected embryos were cultured overnight, and only early blastocysts and morulae were transferred into the corresponding surrogates. For each cell type, we performed at least 4 embryo transfers (Table 2). For SKPs-injected embryos, 2 litters of viable piglets [9] were born by a Caesarean section and by a spontaneous vaginal delivery. For NSCs-injected embryos, 8 fetuses were recovered in the middle of gestation, and 4 piglets were collected by a C-section. For fibroblast-injected embryos, 6 fetuses were collected from 1 surrogate (Table 2). Genomic DNA was extracted from the diverse tissues (lung, heart, liver, spleen, kidney, brain, skin, muscle, intestine, ovary, pancreas, and stomach) of putative chimeric piglets and subjected to PCR amplification with EGFP primers (Table 1). Two (piglets 14-1 and 14-4) out of 9 piglets carried the EGFP transgene in SKP-derived chimeras, but EGFP was present in neither the fibroblast-derived fetuses (n=6) nor the NSCs-derived fetuses (n=8) and piglets (n=4) (Fig. 2D). The failure of contribution to chimeras in fibroblast-derived embryos suggests that SKPs compromise their developmental potential when they differentiate into fibroblasts. Furthermore, porcine fetal-derived NSCs are incapable of incorporating into early-stage embryos and contributing to postnatal chimeras, which is consistent with the previous report that NSCs purified by fluorescence-activated cell sorting (FACS) failed to contribute to mouse chimeras [23].
For SKPs, 4 embryo transfers were initially performed by using oocytes obtained from the ovaries from a local slaughterhouse, and the other 4 transfers used ART oocytes purchased from ART LLC. For NSCs and SKP-derived fibroblasts, only ART oocytes were selected, because they showed a higher pregnancy rate in later studies. We did not intend to recover SKP-derived chimeric fetuses, because we had already had 2 chimeric fetuses collected in our previous study [6].
NA, nonassessment; SKP, skin-derived progenitor; NSCs, neural stem cells.
Surprisingly, the EGFP transgene was present in various tissues of 2 SKP-derived piglets, including the lung, heart, liver, spleen, kidney, brain, skin, muscle, gut, ovary, pancreas, and stomach (Fig. 2E, F), thus representing the derivatives of 3 germ layers (ectoderm: skin, brain; mesoderm: kidney, muscle, heart, ovary, and spleen; and endoderm: lung, liver, gut, stomach, and pancreas).These results suggest that porcine SKP cells can contribute to the derivatives of the 3 germ layers in chimera, and are especially capable of contributing to the tissues derived from the endoderm.
Interestingly, both these EGFP positive piglets were female, but we used male donor cells for the injection. The sex ratio is biased toward males (XY) in mouse chimeras when male and female embryos are paired. In addition, it is highly unlikely that any functional gametes are of female lineage in the phenotype of male chimeras [24]. Previous studies showed that in pigs when the donor ICM cells were XX female and blastocysts were XY male, the resulting chimera was male with no germline cells formed from the donor cell lineages. However, when the donor ICM cells were XY male and the blastocysts were XX female, the chimera was male and the germline cells were derived only from male donor cells [15]. To further confirm the chimerism of the 2 female piglets in the current study, PCR amplification (45 cycles) was performed by using male-specific SRY gene primers. The SRY copy was distributed in several tissues of the 2 EGFP-positive female chimeric piglets (Fig. 2G), thus verifying their chimerism. Notably, the SRY gene copy was detected in the ovaries of these 2 female chimeric piglets. Since there are many somatic cells in the ovary, the germline transmission ability is still questionable. In addition, the male XY donor cells did not dominate the phenotypic sex of the chimeras, because they were visibly female. Furthermore, various tissues of these 2 chimeric piglets were fixed, embedded, sectioned, and then immunostained with anti-GFP antibody from different vendors (Abcam and Invitrogen). Though these antibodies worked efficiently with positive and negative controls, no reliable signals were observed from tissue sections of these 2 SKP-derived chimeric piglets (Fig. 3). In addition, ear fibroblasts were derived from the chimeric piglets and immunostained with anti-GFP antibody. Unexpectedly, we did not detect any EGFP signal either by immunocytochemistry (ICC) or by directly observing green fluorescence under UV light (Fig. 4). We reasoned that the extremely low amount of donor SKP cells in chimeras could only be detected at the level of genomic DNA rather than at a protein level, and the contribution of porcine SKP cells into chimeras was very low. The injected SKPs could incorporate into host embryos but did not proliferate as rapidly as the host embryonic cells. Thus, the majority of injected cells may either undergo apoptosis, or only a very few survive and finally contribute to an extremely limited number of somatic cells in various tissues around the chimeras.

IHC analysis of the chimeric piglets. Various tissues from chimeric piglets 14-1 and 14-4 were fixed, embedded, sectioned, and immunostained with anti-GFP antibody. No significant signals were detected in these tissue sections, indicating a lack of EGFP expression at the level of proteins. Here, only chimeric piglet 14-1
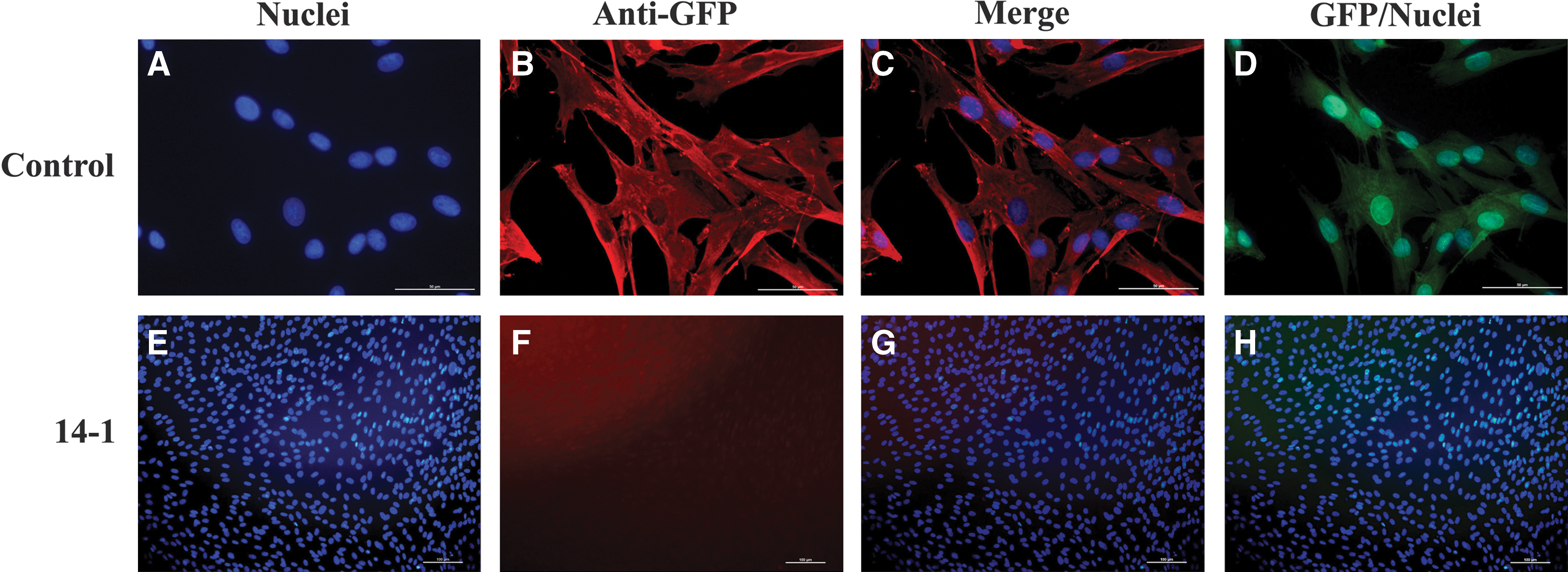
Immunocytochemistry analysis of the fibroblast cells derived from the chimeric piglets. Ear fibroblast cells derived from the piglets 14-1 and 14-4 were fixed and immunostained with the antibody against EGFP. Unexpectedly, no EGFP-positive signal was detected by either ICC or direct fluorescence observation under UV light in the 2 chimeric piglets. Only the piglet 14-1 was shown here
Conclusions
In conclusion, porcine SKPs can incorporate into early-stage embryos; contribute to various somatic tissues of the 3 germ layers in the postnatal chimera; and, especially, have an endodermal potency. However, this developmental potential is lost when they differentiate into fibroblast cells. Fetal brain-derived NSCs fail to incorporate into host embryos and contribute to chimeric piglets. Therefore, neural crest-derived SKPs may represent a more primitive state than their counterpart NSCs in terms of their contributions to multiple cell lineages. Alternatively, precompact (4-cell and 8-cell stage) embryos may provide a unique environment for reprogramming porcine SKPs into a more primitive state during the process of embryonic compaction.
Footnotes
Acknowledgments
The authors would like to thank Dr. Mark D. Kirk (Division of Biological Sciences, University of Missouri) for a critical reading of the article and Jason Henderson for IHC analysis (VMDL, College of Veterinary Medicine, University of Missouri). This study was funded by the NIH National Center for Research Resources (R01RR013438) and Food for the 21st Century at the University of Missouri-Columbia.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.