
Abstract
During evolution, the innate and adaptive immune systems were developed to protect organisms from non-self substances. The innate immune system is phylogenetically more ancient and is present in most multicellular organisms, whereas adaptive responses are restricted to vertebrates. Arthropods lack the blood cells of the lymphoid lineage and oxygen-carrying erythrocytes, making them suitable model animals for studying the regulation of the blood cells of the innate immune system. Many crustaceans have a long life span and need to continuously synthesize blood cells, in contrast to many insects. The hematopoietic tissue (HPT) of Pacifastacus leniusculus provides a simple model for studying hematopoiesis, because the tissue can be isolated, and the proliferation of stem cells and their differentiation can be studied both in vivo and in vitro. Here, we demonstrate new findings of a physical link between the HPT and the brain. Actively proliferating cells were localized to an anterior proliferation center (APC) in the anterior part of the tissue near the area linking the HPT to the brain, whereas more differentiated cells were detected in the posterior part. The central areas of HPT expand in response to lipopolysaccharide-induced blood loss. Cells isolated from the APC divide rapidly and form cell clusters in vitro; conversely, the cells from the remaining HPT form monolayers, and they can be induced to differentiate in vitro. Our findings offer an opportunity to learn more about invertebrate hematopoiesis and its connection to the central nervous system, thereby obtaining new information about the evolution of different blood and nerve cell lineages.
Introduction
N
Invertebrate blood cells (hemocytes) are essentially comparable to cells of the myeloid lineage in vertebrates, because they participate in innate immunity and blood clotting [1]. Drosophila melanogaster and the crayfish Pacifastacus leniusculus are well established as model organisms in the field of invertebrate innate immunity [2 –4], and these species have provided major contributions to hematopoiesis research in invertebrate animals. Since they lack oxygen-carrying red blood cells (erythrocytes) and blood cells of the lymphoid lineage, which participate in adaptive immune defense, these animals offer a simple model system that studies the development of the cells of the innate immune system. In Drosophila, mature hemocytes are formed in the early embryonic head mesoderm and during the larval stage in a specialized organ called the lymph gland (LG) [3]. The progenitor cells in the LG differentiate during the larval stages but are not released into the hemolymph until the pupal stage, when most hemocytes are involved in the phagocytosis of degraded larval tissues [5]. Thus, no LG exists in the adult flies. By contrast, crayfish hematopoietic development is an ongoing process throughout the entire life cycle and takes place in the hematopoietic tissue (HPT) covering the dorsal part of the stomach [1]. It is well established that several transcription factors and signal transduction pathways are involved in hematopoiesis and that they are highly conserved between Drosophila and mammals [6]. We have also confirmed the importance of GATA transcription factors in crayfish HPT [7]. Further, the Runx protein homologs in crustaceans and Drosophila are of special interest, because they are closely associated with the differentiation of cells that express the prophenoloxidase (proPO) gene [4,7]. ProPO is the key enzyme in the proPO-system, an important innate immune system in invertebrates [2]. In the freshwater crayfish, 3 morphologically different classes of hemocytes, hyaline cells, semigranular cells (SGCs), and granular cells (GCs), are observed within the hemolymph, and we have demonstrated that Runx expression is a prerequisite for the final differentiation of SGCs and GCs, both of which express proPO [7].
The transcriptional regulation of hematopoietic cell development in vertebrates and invertebrates shares many common features. Hematopoiesis is regulated by humoral factors in vertebrates, such as numerous cytokines, but the knowledge about the cytokine regulation of hematopoiesis is limited in invertebrates [1]. The evolution of these factors appears to be far more complex, and a few hematopoietic cytokines are found in invertebrate genomes [1]. A technique to culture the crayfish hematopoietic progenitor cells in vitro has enabled us to isolate and characterize a new group of cytokines named astakines (Ast1 and Ast2) from crayfish plasma [8,9]. Astakines contain a conserved domain that is similar to vertebrate prokineticins, which are known to influence the processes related to development of the nervous system, immunity, and hematopoiesis [10,11]. Ast1 is an important regulator at the crossroads between proliferation/differentiation and apoptosis [8,12]. Ast1 acts by regulating the expression of a protein that is responsible for preventing apoptosis, crustacean hematopoietic factor (CHF), which has similarity to the N-terminal region of the human cysteine-rich transmembrane BMP regulator 1 (chordin-like), also known as CRIM1 [12]. A schematic illustration showing the different hematopoietic lineages in crayfish is shown in Fig. 1. When searching GenBank, we found at least 20 astakines in different crustaceans, scorpions, spiders, ticks, and hemipteran, hymenopteran, and blattodean insects. Surprisingly, these small peptides appear not to be present in dipterans, such as Drosophila or Anopheles [9].
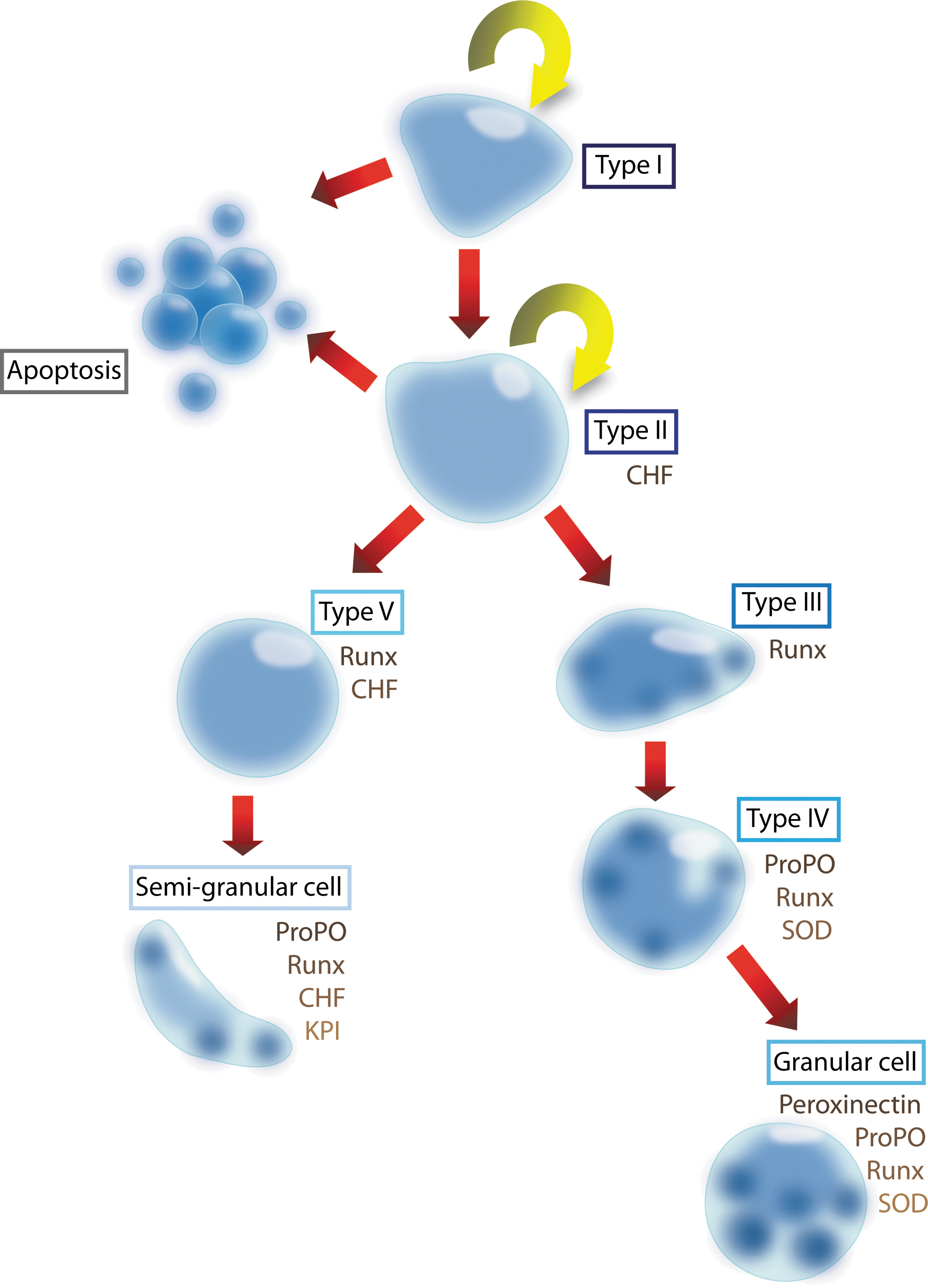
Schematic overview of the hematopoietic lineages in crayfish. Hemocytes are produced in HPT located on the dorsal side of the stomach. Five different cell types are identified in the HPT, where cell type I is a stem cell and type II is a SGC/GC precursor. Cells in the HPT follow 2 main cell lineages: one is from type I-II (the stem cells) via type V cells to SGCs, and the other is from type I-II to types III and IV cells to GCs. Two cytokines are identified in this process: Ast1 and Ast2. Ast1 stimulates the proliferation of HPT cells, induces differentiation of the SGC lineage, and stimulates the release of hemocytes into the circulation by affecting extracellular TGase activity [1,19]. Ast1 induces the expression of CHF, an antiapoptotic factor, and KPI, an SGC marker. By contrast, Ast2 increases the number of mature GCs by an unknown pathway. SOD is a GC lineage marker. proPO and the Runt transcription factor are expressed in mature hemocytes of both the SGC and the GC lineages. HPT, hematopoietic tissue; SGC, semigranular cells; GC, granular cell; CHF, crustacean hematopoietic factor; proPO, prophenoloxidase; KPI, kazal proteinase inhibitor; SOD, superoxide dismutase. Color images available online at
Since nothing was known about the location of different cell types and precursor cells in the HPT, we decided to analyze this tissue in greater detail. In this study, we have characterized different areas of the HPT and found that the tissue has more connections in the animal than was previously thought. We revealed for the first time that the crayfish HPT has a connection to the brain, and we have identified a distinct area of highly proliferative cells close to this connection which constitutes a stem cell center (anterior proliferation center [APC]). We also demonstrated that APC cells can be cultured in vitro and that they differ from other HPT cells in culture.
Materials and Methods
Animals
P. leniusculus (crayfish) from Lake Hjälmaren in Sweden was maintained in aquaria at 10°C with aerated running fresh water. Intermolt crayfish were used for the experiments.
Tissue preparation
The whole stomach covered with the HPT was dissected from crayfish and immediately fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) at 4°C overnight. The HPT was then carefully removed from the stomach, dehydrated, and embedded into paraffin blocks. The embedded tissue was horizontally sectioned with the thickness of 7–11 μm and placed on MENZEL super frost glass slides. For whole mount staining, the tissue was collected from crayfish at 0, 2, or 4 h after a lipopolysaccharide (LPS) or a laminarin injection. After fixation and washing with PBS, the tissue was placed on super frost glass slides, air dried, and subjected to hematoxylin and eosin staining.
Transmission electron microscopy
The HPT was fixed for 1.5 h in ice-cold 1.25% glutaraldehyde in 0.1 M sucrose, 0.1 M sodium cacodylate buffer pH 7.2. The samples were subjected to 15-min rinsing in cold veronal buffer and 2-h postfixed in 2% osmium tetroxide, followed by dehydration in ethanol and acetone. Finally, the samples were embedded in TAAB 812 resin, and thin sections were stained with uranyl acetate, lead citrate, and analyzed with a Philips CM 10 transmission electron microscope.
BrdU incorporation and detection
Crayfish were injected with 10 μL/g fresh weight of 50 mM BrdU in crayfish saline (CFS) (0.2 M NaCl, 5.4 mM KCl, 10 mM CaCl2·2H2O, 2.6 mM MgCl2·6H2O, 2 mM NaHCO3, and pH 6.8). At 1 and 3 h postinjection, the whole stomach covered with the HPT and its connected brain was isolated and immediately fixed as just described. Then, the HPT and the brain were carefully removed from the stomach and used for sectioning or whole mount BrdU detection as previously described [7] (Supplementary Materials and Methods; Supplementary Data are available online at
Detection of mitotic cells in HPT
Mitotic cells in the HPT were detected using a rabbit-anti-Histone H3 (phospho S10) antibody (Abcam). For whole mount staining, the tissues were treated with 2M HCl for 30 min at room temperature before overnight incubation with rabbit-anti-Histone H3 antibody (1:500) at 4°C. Then, the tissues were incubated in the dark with FITC-conjugated anti-rabbit Ig (1:500) (Sigma) for 1 h at room temperature and observed under a fluorescence stereomicroscope. For the detection of mitotic cells in HPT sections, deparaffinized and rehydrated sections were treated with 10 mM sodium citrate buffer, pH6 at 95°C for 10 min and incubated overnight with rabbit-anti-Histone H3 antibody (1:1,000) at 4°C. The sections were then incubated in the dark with Cy3-conjugated anti-rabbit Ig (1:500) (Sigma) for 1 h at room temperature. The sections were later mounted in Vectashield mounting medium containing DAPI (Vector) and observed under a fluorescence microscope.
Detection of PO activity in the HPT
Live crayfish were fixed by an injection of 12.5% glutaraldehyde in PBS. After 15 min, the HPT was dissected and washed 5 times for 10 min in 55 mM sodium phosphate buffer pH 6.8 (PB) containing 0.3% Triton X-100, followed by 3 washes with PB, for 5 min each. For PO activity detection, the tissues were incubated in 5 mM L-dopa, 5% sucrose in PB, with or without 10 mM phenylthiourea (PTU) as a control.
In situ hybridization
Sense and anti-sense specific RNA probes were generated following the manufacturer's instructions of the DIG RNA Labeling Kit (SP6/T7) (Roche). A DNA template of CHF was amplified (see Supplementary Table S1 for primers used), and the PCR products were then purified and used as templates for DIG-labeled RNA synthesis. The labeling efficiency was determined using DIG Quantification Teststrips (Roche) following the manufacturer's instructions. After deparaffinization and rehydration, the tissue sections were treated with 20 μg/mL of proteinase K (Sigma) for 15 min, immediately washed with PBS, postfixed in 4% paraformaldehyde, and washed again. The sections were acetylated with 0.25% acetic anhydride in 0.1M triethanolamine pH 8, and prehybridized in 4× SSC buffer containing 40% deionized formamide at 37°C for at least 30 min. Each section was incubated 16 h at 52°C with 100 μL hybridization solution (40% deionized formamide, 4× SSC, 1× Denharts solution, 0.1 mg/mL yeast tRNA, 0.1 mg/mL salmon sperm DNA, 1 mM DTT, and 7.4% dextran sulfate) containing 60 ng cRNA probe. Posthybridization washes were performed at 52°C using 5× SSC and 0.2× SSC, respectively. The sections were then treated with 20 μg/mL RNase A, washed with 0.2× SSC before blocking at room temperature (RT) for 1 h in maleic acid buffer (100 mM maleic acid, 150 mM NaCl, pH 7.5) containing 1% Blocking Reagent (Roche), and incubated at RT for 1.5 h with 1% Blocking Reagent containing 1:500 of sheep anti-digoxigenin-AP IgG (Roche). A dark purple color was developed for about 12 h in the dark in detection buffer (100 mM Tris-HCl, 100 mM NaCl, and pH 9.5) containing the NBT/BCIP substrate (Roche), and then, methyl green nuclear staining was performed. Finally, the sections were dehydrated and mounted in Euparal mounting medium (WALDECK).
Preparation of collagen I as matrix for the HPT cells
A collagen I solution was prepared by diluting 10.6 mg/mL collagen I stock solution (BD™) to 0.106 mg/mL with PBS; then, 150 μL of the diluted solution was added into 96-well plates; and allowed to form a gel at 37°C for 80 min. The collagen-I matrix-gel plate was sealed, stored at RT, and used within 1 week.
In vitro cell culture
The HPTs were collected and washed several times with PBS. The most anterior part of the HPT that connected to the brain was dissected separately and was named anterior proliferation center (APC) in the following text. The rest of the tissue was named HPTrest. Both parts were treated with collagenase (0.1% of Type I and Type IV; Sigma) as previously described [7]. After suspension in modified L15-medium [8], the cells (1×105 cells/well, 150 μL) were either directly seeded into 96-well plates for the 2D cell culture, or carefully dropped onto the surface of the collagen-I matrix gel for the 3D culture. The cell culture was maintained at 16°C, and one-third of the medium was replaced every second day.
RNAi in vitro
RNAi was performed by mixing 5–10 μg dsRNA (prepared according to Supplementary Materials and Methods) with the culture medium and directly added to the HPT cell cultures. After 1 week, total RNA was extracted, and quantitative RT-PCR was performed to monitor the transcript of crayfish hemocyte maker genes (primers listed in Supplementary Table S1).
Reactive oxygen species detection in HPT
The whole fresh HPT, including the brain, was dissected from crayfish and washed thrice with PBS. Freshly prepared 5 mg/mL of 2′,7′-dichlorofluorescin diacetate (Sigma) in DMSO (Sigma) was diluted with PBS (1:1,000), and the tissue was incubated in this solution for 10 min in darkness. The tissues were then washed 4 times with PBS, transferred to 30% glycerol in PBS, and immediately observed under a fluorescent stereomicroscope.
Results
There is a physical link between the HPT and the crayfish brain
The HPT of P. leniusculus is a thin sheet-like tissue with a lobule-like structure, and it is composed of hemocyte precursor cells in different developmental stages. Earlier, this tissue was thought to be restricted to the dorsal part of the stomach. However, since previous reports have indicated that neuronal stem cells in the crayfish brain are derived from an external source [13,14] and because several genes that we have isolated from crayfish are limited in expression to the HPT and cells in nerve and brain tissues [9,12], we asked whether there is a physical link between the HPT and the brain. By a careful dissection of this thin and fragile tissue, we can now demonstrate that the HPT extends forward and connects to the brain (Fig. 2A–C), which thereby supports that there is a morphological and, possibly, a functional link between these organs.

Location and structure of crayfish HPT and brain. The HPT is located in the dorsal part of the stomach, with the anterior part of the tissue extending forward and linking the tissue to the brain
The HPT has an APC close to the brain
In the Drosophila LG, the stem cells cluster together next to the dorsal vessel [medullary zone (MZ)], whereas differentiating cells that develop mainly in the late larval stages are located in the surrounding area, called the cortical zone (CZ) [15,16]. To identify similar areas in the crayfish HPT, we first studied the structure of the HPT by light microscopy and hematoxylin and eosin staining. The tissue consists of numerous tightly packed cells, and some areas located in the middle of each side of the ophthalmic artery (AO) have more loosely associated lobules with large amounts of space between them (Fig. 3A, arrows). A difference in nuclear staining intensity after hematoxylin and eosin treatment was found in the HPT cells (Fig. 2D–G). Some cells show dark nuclear staining with mainly condensed chromatin (heterochromatin), indicating more differentiated cells (black arrow in Fig. 2G). Other cells show light nuclear staining, indicating mainly loose chromatin (euchromatin) as in immature cells and stem cells (white arrow in Fig. 2F and Supplementary Fig. S1A, B). Different amounts of those cells were found at different locations of the HPT. The cells with dark nuclear staining were found at a high percentage in the posterior HPT (75%) and were present in a significantly lower amount in the APC (43%). In contrast, the cells with light nuclear staining were high in number in APC (57%) and were found with significantly lower numbers in the posterior part of HPT (Fig. 2H). Moreover, the area with loosely associated lobules, which we named the growth zone (arrows in Fig. 3A), was filled with cells after an injection of either LPS or laminarin, both of which are infection-mimicking treatments known to induce the recruitment of new hemocytes from the HPT (Fig. 3C, D). To determine whether this growth zone close to the vessel was a center for cell division, we examined the proliferation of the HPT by using BrdU incorporation. However, proliferating cells were found mainly in the most anterior part of the HPT, especially around the area connected to the brain and at the edges around the HPT (Fig. 4A), whereas significantly fewer BrdU incorporating cells were detected either in the center or in the growth zone. When the incorporation time was extended to 3 h, the area with a strong BrdU signal expanded over the anterior part of the HPT from the edges and moved inward (Fig. 4B). However, the area surrounding the AO and the growth zone still contained a few BrdU-positive cells (Fig. 4D–I), but the percentage of BrdU-positive cells in this area is lower than in the anterior part of the tissue (Fig. 4C). In addition to investigating cell proliferation by BrdU incorporation, mitotic cell division in the tissue was also detected (Figs. 5 and 6). In agreement with BrdU incorporation, more mitotic cells were found in the APC, which is the most anterior part of the HPT (Figs. 5A, C and 6A) when compared with the posterior HPT (Figs. 5B, D and 6B). To investigate whether the cells in the growth zone were induced to proliferate only during emergency or during an infection, mitotic cells in the HPT were examined at 30 min and 4 h after the LPS injection. The amounts of these cells were significantly increased at 30 min after the LPS injection and especially in the anterior part of the HPT, and the level of mitosis decreased to the normal level again at 4-h post-LPS injection (Fig. 6).
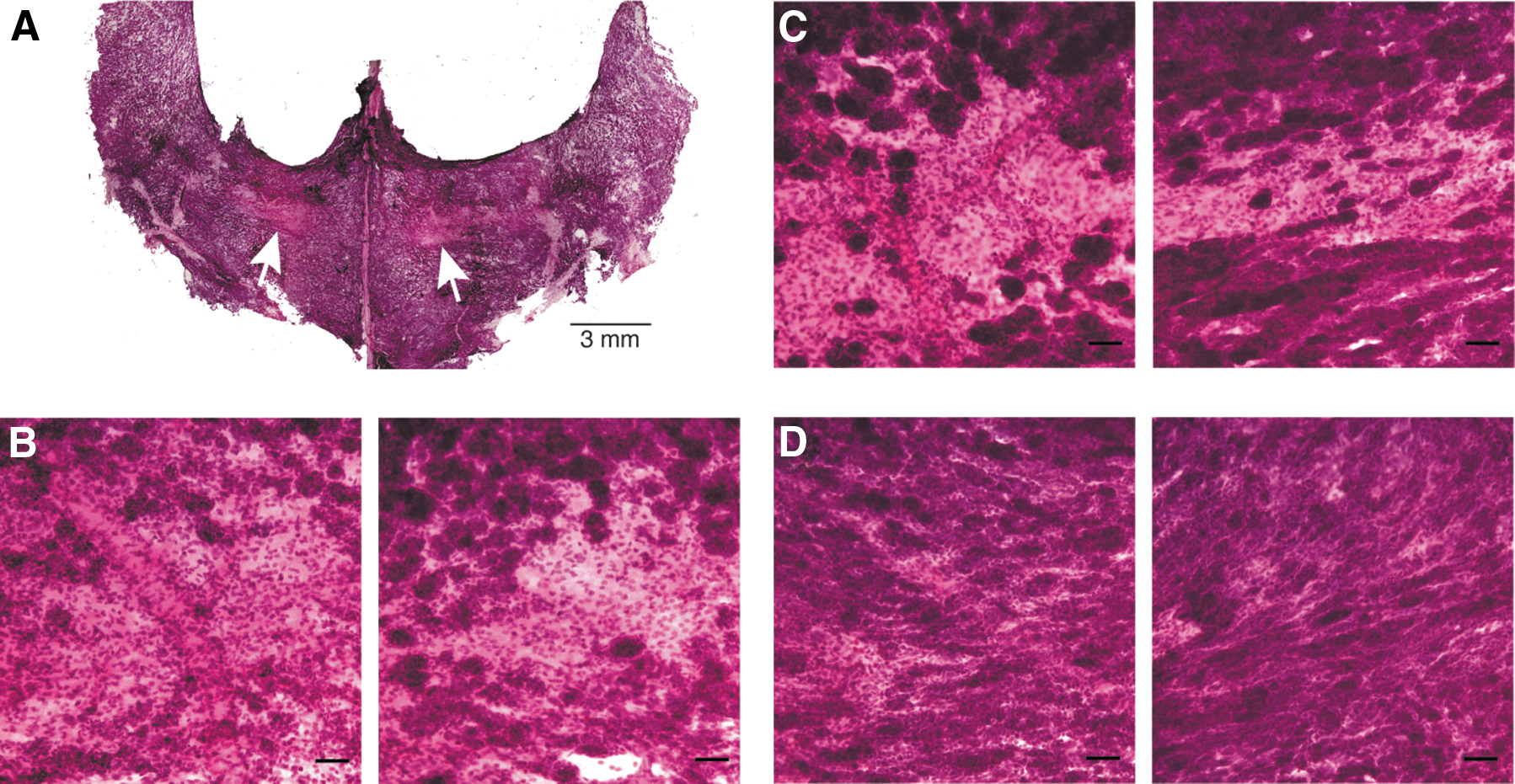
Accumulation of the cells in the growth zone after LPS injection. Some areas of the HPT consist of tightly packed cells, whereas some areas have loosely connected cell lobules

BrdU incorporation in the HPT. At 1 h

Mitotic cells in APC and posterior/dorsal HPT. Mitotic cells (red) were detected in APC

Increase of mitotic cells in HPT after the LPS injection. The mitotic cells in HPT were detected using anti-phospho-histone H3 antibody (shown in green). Under normal condition, the mitotic cells are mainly present in the anterior part
In summary, our data indicate that the HPT in crayfish is organized differently from the Drosophila LG and that cells with a high proliferation rate are located in the anterior part of the HPT, especially in the area close to the brain. We suggest that this region is the major location of the multipotent hematopoietic stem cells of the P. leniusculus HPT, and, thus, we named it the APC.
Reactive oxygen species signaling is high in the anterior part of the HPT
Since increased levels of reactive oxygen species (ROS) have been found to induce the differentiation of hematopoietic stem cells in Drosophila and in the myeloid lineage of mammals [16,17], we decided to test ROS activity during HPT differentiation. High ROS production was only found in a small area in the most anterior part of the HPT between the APC and the brain in normal crayfish and crayfish injected with CFS (Fig. 7A, C). When laminarin was injected, the hemocyte number in the peripheral blood circulation was dramatically reduced to a ∼5% within 30 min (Fig. 7B) and at 60 min postinjection, the hemocyte number started to increase, and differentiation was initiated. At 30 min postinjection with laminarin, a stronger signal of ROS was detected that was also extended throughout the APC (Fig. 7D) relative to that observed 30 min after the CFS injection (Fig. 7C). However, at 1 h after the injection, no obvious difference in the ROS signal was observed between CFS- and laminarin-injected crayfish (data not shown). The increase in ROS activity after the laminarin injection corresponds in time with a rapid loss in circulating hemocyte number that is followed by a rapid recruitment of new hemocytes from the HPT (Fig. 7B), which suggests that an increase in metabolic activity is associated with differentiation. In a previous study, we demonstrated that hematopoiesis in crayfish is regulated in a circadian rhythmic manner [18]. Interestingly, the ROS activity in close proximity to the APC was either very low or nearly absent, at 30 min after the light comes on in the morning (Fig. 7E, F). This time coincides with the time of the day when the hemocyte number in the circulation is at its highest, which could indicate that the hematopoietic activity goes down when there is a lesser need for new hemocytes.
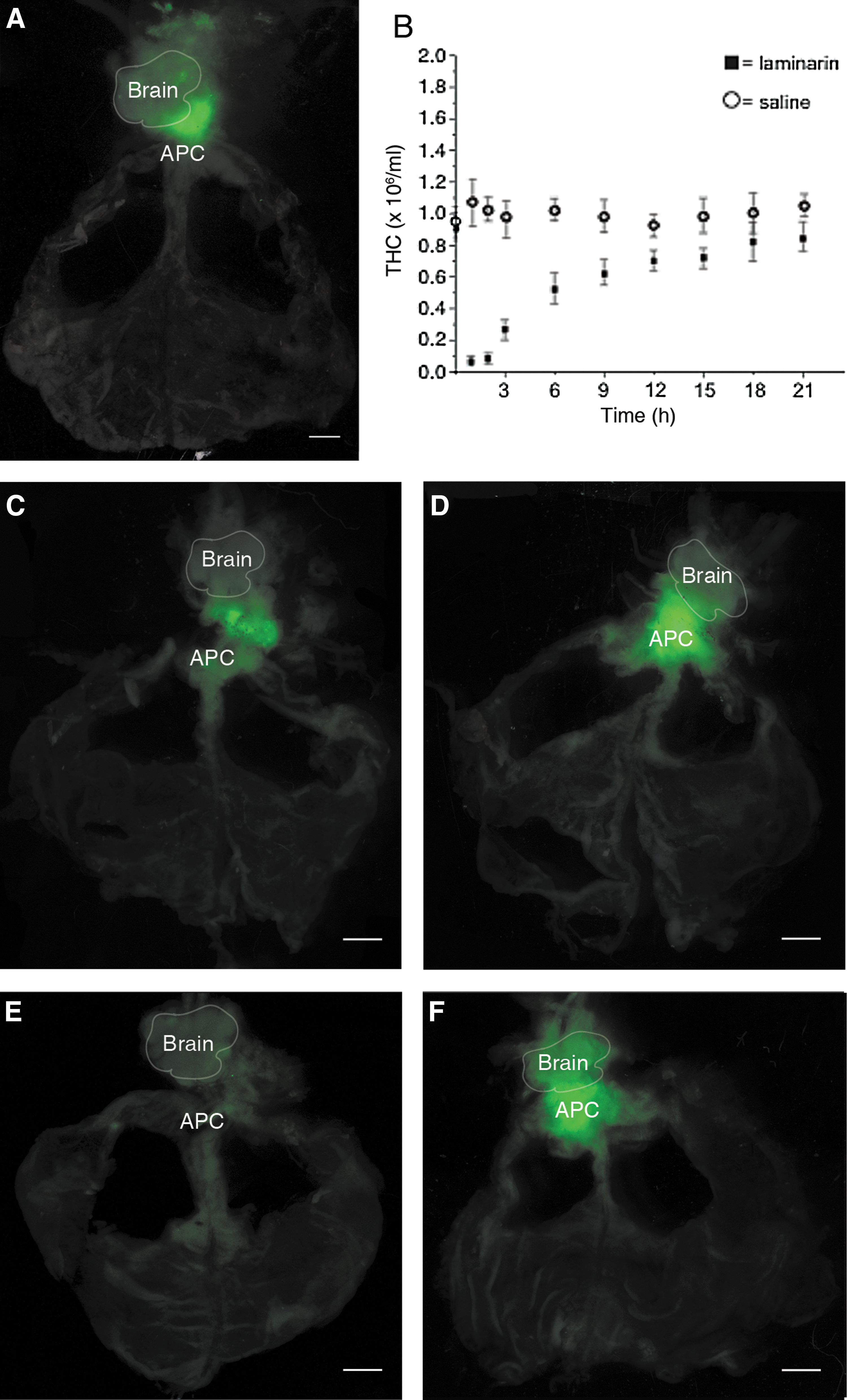
ROS activity in the HPT. ROS (green) was detected in fresh HPT. In normal crayfish
Low expression of the anti-apoptotic factor, CHF, is found in proliferative areas
Apoptosis is an ongoing process in the crayfish HPT [7], and CHF is important for limiting apoptosis in the HPT when new hemocytes have to be produced and secreted into the circulation [12]. Earlier, this gene was found to be exclusively expressed in HPT and mature SGCs [12]. In order to establish whether SGCs and GCs develop in the same or in separate lobules, we investigated where CHF-expressing cells are located in the HPT. We performed in situ hybridization, and the results revealed CHF mRNA expression in all parts of the HPT excluding the edges areas (Fig. 8A, B), where cell proliferation was high. The cells expressing CHF were spread over the remaining tissue; however, not all cells in each HPT lobule had high CHF expression (Fig. 8D), which may reflect that CHF is restricted to SGC and HPT cells and that cells of the GC lineage do not express this transcript [12]. In some lobules, no CHF mRNA was detected, indicating that these lobules are only producing GCs (Fig. 8D, black arrow). When we further analyzed the localization of apoptotic cells by TUNEL assay, the highest rate of apoptosis was detected in the lobules near the edges of the tissue, whereas apoptosis (green areas Supplementary Fig. S2A) was either very low or even absent in the central part near the artery. Positive cells were carefully examined for typical morphology to confirm apoptosis (Supplementary Fig. S2A inset). To stimulate new hemocyte synthesis, we injected LPS or laminarin into the hemolymph and observed that CHF expression was increased in all parts of the HPT (Supplementary Fig. S2B). New hemocytes are needed to compensate for the loss after the laminarin injection (Fig. 7B), and a similar decrease in apoptosis was observed in corresponding areas of the HPT after an LPS injection (data not shown).

Expression of CHF in different parts of the HPT. Expression of CHF in the different parts of HPT detected by ISH indicates that CHF is evenly distributed throughout the tissues excluding the edges
Melanization of cells owing to PO activity does not occur inside the HPT lobules
PO is a marker for hemocytes that are close to full maturation and for mature hemocytes; nondifferentiated cells do not have PO activity. Mature SGCs as well as GCs and the late GC precursors (type 4 cells, Fig. 1) express proPO mRNA. Therefore, we decided to localize the HPT cells, which are almost mature, by staining for PO activity in situ (by incubating the tissues in

PO activity in the HPT. PO-positive cells are found in the posterior part of the HPT, and PO-positive hemocytes can also be observed within the AO (arrow)
Our new findings show proliferating cells at the edges of the HPT and an APC, which is adjacent to the brain. More developed cells are found in the central parts of the tissue near a growth zone and around the large artery.
HPT cells from the APC appear to be different from cells from other parts of the HPT in culture
We have previously reported a method to culture HPT cells in vitro and used this method to isolate the astakine cytokines. The differentiation of the stem cells can be judged by the induction of the marker transcripts KPI for SGCs, and SOD for GCs [1]. However, in our previous culture system, APC cells were not included, and cells belonging primarily to types 2 and 3 (Fig. 1) were cultured (Fig. 10A).

Culture of HPT cells isolated from different parts of the tissue. Type 2 cells are the major dividing cells in HPTrest, as shown by TEM
In our new study, we were able to isolate single cells from the APC and compared their appearance in culture with that of cells of the remaining part, now named HPTrest. The cells from HPTrest could grow as monolayers (Fig. 10B), and they can be induced to differentiate into GC and SGC precursor cells (type 4 and type 5) in accordance with our previous results [9]. However, the cells isolated from the APC had a different appearance in culture. These cells divided rapidly during the first days, and after 3–5 days, small densely packed cell clusters appeared (Fig. 10C–E). Several cells formed bowl-shaped cell clusters after more than 1 week in culture. However, after reaching a certain size at ∼12–14 days, the cell clusters did not increase, and growth stopped. We did not find any signs of differentiation of these cells even when Ast1 or 2 was added to the culture. Conversely, as already demonstrated, HPTrest cells can be induced to differentiate in vitro [9]. The survival and differentiation into SGC of the HPTrest when cultured in a collagen I matrix could be dramatically improved, as indicated by the increased transcription of PCNA (proliferating cell marker), KPI, and SOD (Fig. 10F). Notably, the transcription of KPI increased more than 7-fold. This may imply that the collagen I matrix is favorable for the differentiation of HPT cells along the SGC lineage and also that the differentiation of SGCs is a faster process compared with that of GCs. The HPT cells cultured in the collagen I matrix exhibited higher CHF expression, which implies lower apoptosis. Down-regulation of TGase further stimulated differentiation along the SGC lineage (Fig. 10G), in accordance with our previous results showing that Ast1 stimulates HPT cells to differentiate and induces a decrease in the extracellular TGase activity of these cells [19].
Discussion
Hemocyte production in crayfish occurs in the HPT, which is located in the dorsal part of the stomach surrounding the AO [1]. The HPT contains densely packed hematopoietic lobules, and the cells in the APC and the dorsal HPT have a different morphology. More than 50% of the cells in the APC contain nuclei with loose euchromatin, whereas most of the cells in the dorsal HPT contain condensed heterochromatin. The structure of the chromatin has been reported to be less condensed in undifferentiated embryonic stem cells in mammals, and the chromatin rearranges to form highly condensed heterochromatin during cell differentiation [20]. Thus, we speculate that the cells in the APC constitute the multipotent stem cells of the HPT. Further, we were able to culture cells from the APC in vitro, and these cells developed into densely packed cell clusters after 3–7 days of culture; conversely, the cells from the posterior HPT (HPTrest), as were previously demonstrated, formed monolayers, divided sporadically, and differentiated in vitro after induction [8].
When LPS or laminarin is injected, cells accumulate in the central areas of the tissue, resulting in the release of new hemocytes. This injection also induced the proliferation of the HPT cells, especially in the anterior part of the tissue. This observation indicates that there is a dynamic change in the HPT in response to LPS-induced blood loss, similar to that observed in response to an infection by bacteria (which contain LPS) or fungi (which contain laminarin). This is in agreement with our previous reports demonstrating that the injection of LPS or laminarin into crayfish results in the elevated synthesis of hemocytes and the subsequent release of hemocytes from the HPT [7,8]. A similar response has also been observed during physiological stress in mice. Bleeding or an LPS injection into mice induced the release of hematopoietic progenitors from the bone marrow (BM) into the blood circulation through the activation of osteoclasts, which degraded the BM endosteal compartment. Osteoclasts are bone cells derived from HSCs, and they require signals from osteoblasts to proliferate, differentiate, and function [21,22].
Here, we demonstrate that the anterior part of the HPT extends forward and links the HPT with the brain. This anatomical connection possibly facilitates physiological communication between the HPT and brain. For example, it has been reported that hemocyte number and the plasma level of the hematopoietic cytokine, Ast1, oscillates in a typical circadian rhythmic manner in crayfish [18]. Moreover, a study conducted on adult Procambarus clarkia crayfish suggested that HPT cells possibly serve as a primary source of neuronal precursor cells [13]. In mammals, a brain-bone-blood link has been proposed to require signals delivered from the central nervous system directly to the hematopoietic stem and progenitor cells or indirectly to the niche that controls HSC proliferation, mobilization, and differentiation [22]. In humans, the percentage of BM cells in the S-phase and colony-forming units exhibited circadian variations [23]. In mice, circadian-regulated HSC release from the BM is correlated with the secretion of adrenergic hormones from the suprachiasmatic nucleus [24].
In this study, we discovered an APC in the anterior part of the HPT in close connection to the brain. This location is very different from that in the fly, in which an APC is localized in the MZ close to the dorsal vessel [15]. No connection between the Drosophila LG and the brain has been described, which may indicate different functions for arthropod HPTs. However, the peripheral nervous systems (PNS) has been reported to be a place where mature circulating larval hemocytes of Drosophila resides and the PNS have been shown to be important for hemocyte survival [25]. The primary lobes of the Drosophila LG develops a larger CZ during the final larval stages, and since crayfish are much more long living animals with adult hematopoiesis, the major parts of the HPT may correspond to this CZ of the Drosophila LG, whereas a putative MZ may be minor and difficult to detect without molecular markers. However, our results showing changes in ROS activity linked to proliferation indicates that the APC may play a similar role in crayfish as the MZ in Drosophila.
Aerobic metabolism usually causes the production of ROS, which exposes cells to the risk of oxidative stress. The maintenance of an optimal O2 level is, therefore, important for proper cell function, especially in stem cells. In mammals, low O2 levels or hypoxic conditions have been reported to play a role in establishing a stem cell niche and controlling stem cell behavior [26]. However, different stem cells require different optimal O2 environments. For example, mesenchymal stem cells occupy a 2%–8% O2 niche, whereas ranges of 1%–8% and 1%–6% O2 are required for neural stem cells and HSCs, respectively [27]. Moreover, in the mouse BM, 2 different niches that are related to different O2 levels are present and required for 2 subpopulations of HSCs. The low-O2/hypoxic osteoblastic niche is possibly necessary for maintaining long-term HSC quiescence and protecting the HSCs from the harmful effects of ROS accumulation, whereas the high-O2/vascular niche in the central BM is a residence for short-term HSCs and primes the HSCs to proliferate [28]. In mice, ROS were demonstrated to play a role in hematopoietic reconstitution [17], the HSC transition to myeloid progenitors, self-renewal capacity, and HSC quiescence maintenance [29,30]. In Drosophila, high ROS levels were found in the hematopoietic progenitor cells in the MZ. Stimulating an elevation of ROS resulted in the differentiation of the progenitor cells to all hemocyte types in the Drosophila LG via the activation of the JNK and FOXO signaling pathways and the down-regulation of Polycomb [16]. In addition, the increase of ROS was detected in the posterior signaling center (PSC) of Drosophila LG after a wasp infestation. This elevation regulates the Drosophila immune response, as it serves as a signal that primes the progenitor cells in LG to differentiate into lamellocytes [31]. Here, high ROS levels were detected in a specific area between the brain and the APC of the HPT, and the level of ROS formation was increased in response to a laminarin injection. The high ROS levels are possibly the result of a high metabolic rate of the cells in this area, which could serve as a unique microenvironment or a niche to maintain proliferative behavior. However, it is difficult to conclude whether the increase in ROS production is essential for differentiation or whether it only reflects a high metabolic rate during the differentiation process.
Melanization is an important innate immune reaction in invertebrates [2,32] that is mediated by mature hemocytes expressing proPO. PO-positive cells were mainly found in mature hemocytes in the AO and in a small population of HPT cells in the posterior part of the HPT. These results confirm our previous observations that a low percentage of HPT cells express the proPO transcript, whereas mature hemocytes have high proPO expression [7]. Another report in Drosophila revealed that differentiation into PO-active crystal cells is a postmitotic event [4]. From this information, we propose that the APC is where multipotent HSCs are located and that more differentiated cells reside in the central and posterior parts of the tissue. These differentiated cells are possibly already committed to undergoing final maturation on release from the HPT.
Conclusion
The HPT of P. leniusculus provides a simple model system for studying hematopoiesis, because the tissue can be isolated and the proliferation of stem cells and their differentiation into mature hemocytes can be studied both in vivo and in vitro. Moreover, our discovery of an anterior proliferation center in close connection to the brain offers an opportunity to explore the regulation of invertebrate hematopoiesis and its connection to the central nervous system.
Footnotes
Acknowledgments
The Swedish Research Council VR 621-2009-5715, VR 319-2010-6250 to K.S., and VR621-2011-4797 to I.S. supported this work.
Author Disclosure Statement
The authors hereby certify that they have no nonfinancial or commercial associations that might create a conflict of interest in connection with the submitted article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.