
Abstract
Multipotent mesenchymal stromal cells (MSCs) from the human olfactory mucosa (OM) are cells that have been proposed as a niche for neural progenitors. OM-MSCs share phenotypic and functional properties with bone marrow (BM) MSCs, which constitute fundamental components of the hematopoietic niche. In this work, we investigated whether human OM-MSCs may promote the survival, proliferation, and differentiation of human hematopoietic stem cells (HSCs). For this purpose, human bone marrow cells (BMCs) were co-cultured with OM-MSCs in the absence of exogenous cytokines. At different intervals, nonadherent cells (NACs) were harvested from BMC/OM-MSC co-cultures, and examined for the expression of blood cell markers by flow cytometry. OM-MSCs supported the survival (cell viability >90%) and proliferation of BMCs, after 54 days of co-culture. At 20 days of co-culture, flow cytometric and microscopic analyses showed a high percentage (73%) of cells expressing the pan-leukocyte marker CD45, and the presence of cells of myeloid origin, including polymorphonuclear leukocytes, monocytes, basophils, eosinophils, erythroid cells, and megakaryocytes. Likewise, T (CD3), B (CD19), and NK (CD56/CD16) cells were detected in the NAC fraction. Colony-forming unit–granulocyte/macrophage (CFU-GM) progenitors and CD34+ cells were found, at 43 days of co-culture. Reverse transcriptase–polymerase chain reaction (RT-PCR) studies showed that OM-MSCs constitutively express early and late-acting hematopoietic cytokines (i.e., stem cell factor [SCF] and granulocyte- macrophage colony-stimulating factor [GM-CSF]). These results constitute the first evidence that OM-MSCs may provide an in vitro microenvironment for HSCs. The capacity of OM-MSCs to support the survival and differentiation of HSCs may be related with the capacity of OM-MSCs to produce hematopoietic cytokines.
Introduction
H
In vitro, it has been difficult to enhance the self-renewal and/or expansion of HSCs without stromal cells, even if all known exogenous growth factors and other materials are added to the cultures [4 –6]. Despite this, several approaches have shown that human and mouse long-term hematopoiesis can be maintained by co-culturing HSCs with cell lines [7 –13] or stromal cells, as feeder layers [14 –19].
To date, multipotent mesenchymal stromal cells (MSCs) have shown the most promise cells for promoting in vitro hematopoiesis, as they support not only similar stromal and HSC interactions as those seen in the BM microenvironment [20,21], but also maintain the pluripotential characteristics of HSCs and the functionality of progenitor cells [22 –24]. Although the most important source of MSCs is BM, these cells have been also isolated from various other sources [25 –32]. Thus, MSCs from placenta, lung, and umbilical cord blood have been shown to support expansion of HSCs and hematopoietic progenitor cells [33,34].
MSCs from human olfactory mucosa (OM) have been recently isolated and characterized [35 –38]. It has been reported that OM-MSCs have morphologic and phenotypic similarities with BM-MSCs [37]. Likewise, OM-MSCs have also the capacity to differentiate into ectoderm and mesoderm cell types [37,39]. These similarities have led us to investigate whether human OM-MSCs can be used as an in vitro microenvironment to support expansion and differentiation of human HSCs. In the present study, we show that OM-MSCs support in vitro hematopoiesis.
Materials and Methods
Reagents
Fluorescein isothiocyanate (FITC) or phycoerythrin (PE)-conjugated mouse monoclonal antibodies anti-human CD90, CD73, CD166, CD49b, CD45, CD3, CD19, CD16, CD56, and CD34 were purchased from Becton Dickinson. Trizol was obtained from Sigma-Aldrich.
Isolation and culture of OM-MSCs
Human OM-MSCs were isolated from nasal mucosa biopsies obtained from patients undergoing nasal surgery under general anesthesia, as described before [40]. All patients gave their informed consent to participate in the study, and the study protocol was approved by the institutional review board of the participating institutions. OM-MSCs were obtained as previously described [35,38]. Briefly, biopsy specimens were dissected for explant cultures. Each explant was placed in 24-well plates with alpha-minimum essential medium (MEM)/Chang medium containing 20% fetal bovine serum (FBS) (Sigma). Two days after plating, cells began to migrate from explants as plastic-adherent cells. Two weeks later, adherent cells reached confluency and they were collected and seeded in 24-well plates with alpha-MEM/Chang medium containing 20% FBS.
Isolation and culture of human bone marrow cells
Human BM was obtained by aspiration from the posterior iliac crest of healthy adult volunteers. Informed consent was obtained for all BM collections, and the study was approved by the institutional bioethical committee. BM mononuclear cells were isolated after Ficoll-Paque (GE Healthcare Biosciences) density gradient, and cultured in alpha-MEM/Chang medium. After 24 h of culture, the nonadherent cell (NAC) fraction was collected and plated on a monolayer of OM-MSCs.
OM-MSC and bone marrow cell co-cultures
Bone marrow cells (BMCs) were seeded (15×104 cells per well) onto a confluent monolayer of OM-MSC cultures in 24-well plates with alpha-MEM/Chang medium containing 20% FBS. Half of the co-culture medium was collected and replaced each 5–7 days with fresh medium. The number and the viability of cells present in the NAC fraction of the BMC/OM-MSC co-cultures were determined by trypan blue staining. Aliquots of NACs from each well were examined for expression of HSCs, myeloid, and lymphoid cell markers by flow cytometry, and for morphology in cytospin and for morphology in cytospin preparations by May-Grünwald/Giemsa staining.
Flow cytometry analysis of MSCs and hematopoietic marker expression
To evaluate the expression of MSC markers, OM-MSCs were grown to 90% of confluency, and then the cells were harvested and analyzed by flow cytometry for the expression of the CD90, CD73, CD166, and CD49b markers. On the other hand, NACs were harvested from BMC/OM-MSC co-cultures and analyzed for the expression of HSC (CD34), pan-leukocyte (CD45), and lymphoid (CD3, CD19, CD16, and CD56) markers. Analysis of HSCs and blood cell markers was performed every 7 days. Simultaneous negative control staining reactions were performed by incubating the cells with the FITC- and PE-labeled IgG isotype. Data collection and analysis of the fluorescent intensities were made using an FACScalibur (Becton Dickinson). Ten thousand events were acquired and analyzed using the Cell Quest software program.
Clonogenic assays
Colony-forming unit–granulocyte/macrophage (CFU-GM) progenitors were assessed by methylcellulose colony-forming assay. For this purpose, NACs were seeded at 3×104 cells in Methocult GF H4034 (Stem Cell Technologies). Colonies were scored after 14 days of culture using standard morphological criteria. Single colonies were collected and spread on a glass slide using a cyto-centrifuge and stained with May-Grünwald/Giemsa to observe cell morphology.
Detection of hematopoietic cytokine transcripts in OM-MSCs by RT-PCR
Total RNA was extracted from monolayers of OM-MSCs using a TRIZOL reagent, following manufacturer's instructions. Reverse transcription was carried out using random hexamer oligonucleotides and 4 U AMV reverse transcriptase (Promega) for cDNA synthesis. Polymerase chain reaction (PCR) amplification of the cDNA was then performed using specific oligonucleotides (Table 1) for the detection of stem cell factor (SCF), granulocyte- macrophage colony-stimulating factor (GM-CSF), interleukin (IL)-6, IL-11, IL-7, and β-actin transcripts. PCR conditions were 1 min of denaturation at 94°C, annealing at 52°C for 1 min, and extension at 72°C for 1 min for 35 cycles. Analysis of the PCR products was performed by comparing them with the predicted PCR fragment size after ethidium bromide staining of the PCR products separated by electrophoresis in a 1.8% agarose gel.
IL, interleukin; SCF, stem cell factor; GM-CSF, granulocyte- macrophage colony-stimulating factor.
Results
Isolation and culture of human OM-MSCs
OM explants were placed in 24-well plates containing culture medium supplemented with 20% FBS. After 48 h of culture, adherent cells were observed migrating from OM explants (not shown). These cells were maintained in culture until they reach a confluent monolayer. Microscopic evaluation of cell cultures showed the presence of cells with elongated and fibroblastoid morphology (Fig. 1A). Flow cytometric analysis of these cells showed that they express the MSC markers CD90, CD73, CD166, and CD49b, but not CD34 (Fig. 1B). Differentiation assays showed the osteogenic, adipogenic, and chondrogenic capacity of these cells (Fig. 1C–F).

Growth, expression analysis, and differentiation of OM-MSCs. MSCs were isolated from human OM and cultured until they reached confluency. Phase-contrast micrographs show cells with fibroblast-stromal morphology
OM-MSCs promote proliferation and survival of human BMCs
We evaluated the capacity of OM-MSCs to support the proliferation and survival of human BMCs in vitro, by seeding BMCs on confluent OM-MSC monolayers, in the absence of exogenous hematopoietic cytokines. Previously we showed by flow cytometry that these cells express the following markers: CD34, CD45, CD3, CD19, CD16, and CD56 (data not shown). At different intervals, and for almost 8 weeks, NACs were harvested from BMC/OM-MSC co-cultures. The highest number of NACs was collected after 54 days of co-cultures, as compared with day 13 (5-fold increase) (Fig. 2A). The viability of NACs harvested from BMC/OM-MSC co-cultures was always >90% (Fig. 2B). Microscopic evaluation of BMC/OM-MSC co-cultures showed the presence of rounded cells, which were increasing in number from day 4 of co-culture, on the OM-MSC monolayer (Fig. 2C).

OM-MSCs promote the proliferation and survival of human BMCs. NACs were collected from BMC/OM-MSC co-cultures and the cell number and viability were determined by trypan blue technique. Results show an increase in the number of NACs at 54 days with respect to 13 days of co-cultures
OM-MSCs promote myelopoiesis from human BMCs
We were interested in evaluating the capacity of OM-MSCs to induce myeloid differentiation from BMCs. For this purpose, NACs were harvested from BMC/OM-MSC co-cultures and evaluated, using flow cytometry, for the expression of the leucocyte common antigen CD45. After 20 days, high percentage (73%) of cells expressing CD45 marker were detected in the NAC fraction (Fig. 3A). Cytospin and May-Grünwald/Giemsa staining of NACs showed the presence of cells of myeloid origin, including polymorphonuclear cells (neutrophils, basophils, and eosinophils), monocytes, erythroid cells, and megakaryocytes (Fig. 3B).

OM-MSCs promote in vitro myelopoiesis from human BMCs. NACs were collected, at day 17, from BMC/OM-MSC co-cultures, and the cells were analyzed for their expression of the CD45 marker by flow cytometry
OM-MSCs promote lymphopoiesis in vitro from human BMCs
We evaluated the capacity of OM-MSCs to support lymphopoiesis from BMCs. Morphological evaluation of the NACs collected from BMC/OM-MSC co-cultures, after 20 days, showed the presence of lymphoid-like cells (Fig. 4A). Flow cytometric analysis showed the presence of a high percentage of CD3+ cells in the NAC fraction (Fig. 4B). Low percentages of cells expressing CD19+ and CD56+/CD16+ were also found in the NAC population (Fig. 4C, D, respectively).
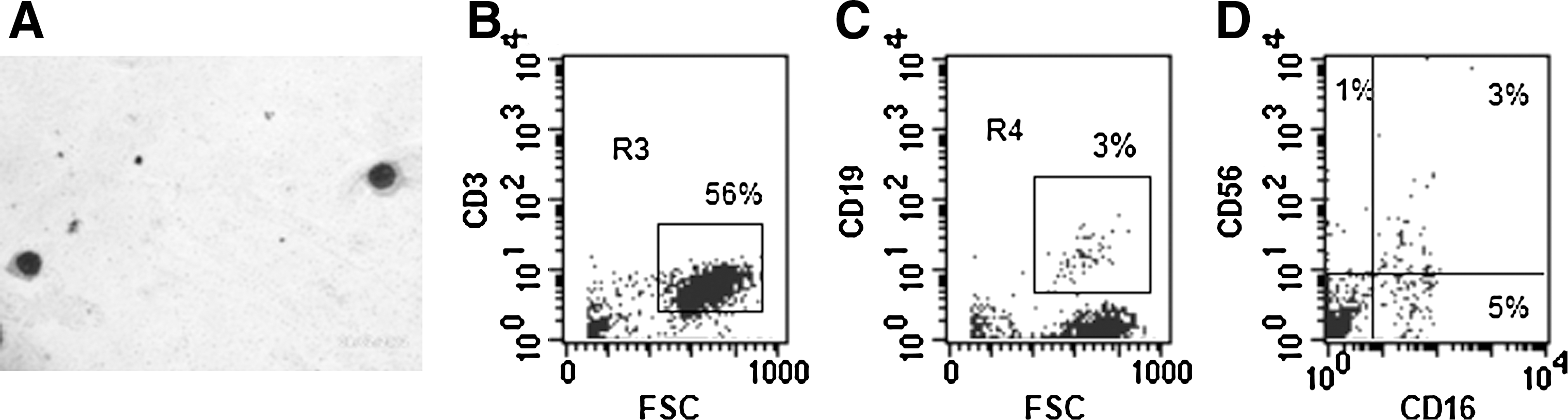
OM-MSCs promote in vitro lymphopoiesis from human BMCs. NACs were collected from BMC/OM-MSC co-culture, at day 17, and the cells were analyzed for their morphology and surface expression markers. Microscopic evaluation shows the presence of lymphoid cells
Detection of hematopoietic progenitors in the nonadhering fraction of human BMC/OM-MSC co-cultures
Because our results show that OM-MSCs promote hematopoiesis in vitro from BMCs in long-term cultures, we investigated the possible presence of hematopoietic progenitors in these cultures. For these purposes, NACs were collected from BMC/OM-MSC co-cultures, after 17 days, and evaluated in clonogenic assays for the presence of CFU-GM progenitors, using a MethoCult system. CFU-GM colonies were detected in the NAC fraction collected from BMC/OM-MSC co-cultures (Fig. 5A). Cytospin and May-Grünwald/Giemsa staining of desegregated CFU-GM colonies showed the presence of granulocytes and monocytes (Fig. 5B). The possibility that OM-MSCs support the survival of primitive HSCs in long-term cultures was investigated by evaluating the presence of CD34+ cells in the adherent fraction of the BMC/OM-MSC co-cultures. After 43 days of co-culture, CD34+ cells were detected in the adherent fraction of BMC/OM-MSC co-cultures (Fig. 5C). It is important to note that OM-MSCs do not express hematopoietic markers (CD34) (data not shown). Together, these findings indicate that OM-MSCs can support the survival of human primitive hematopoietic progenitor cells in vitro.
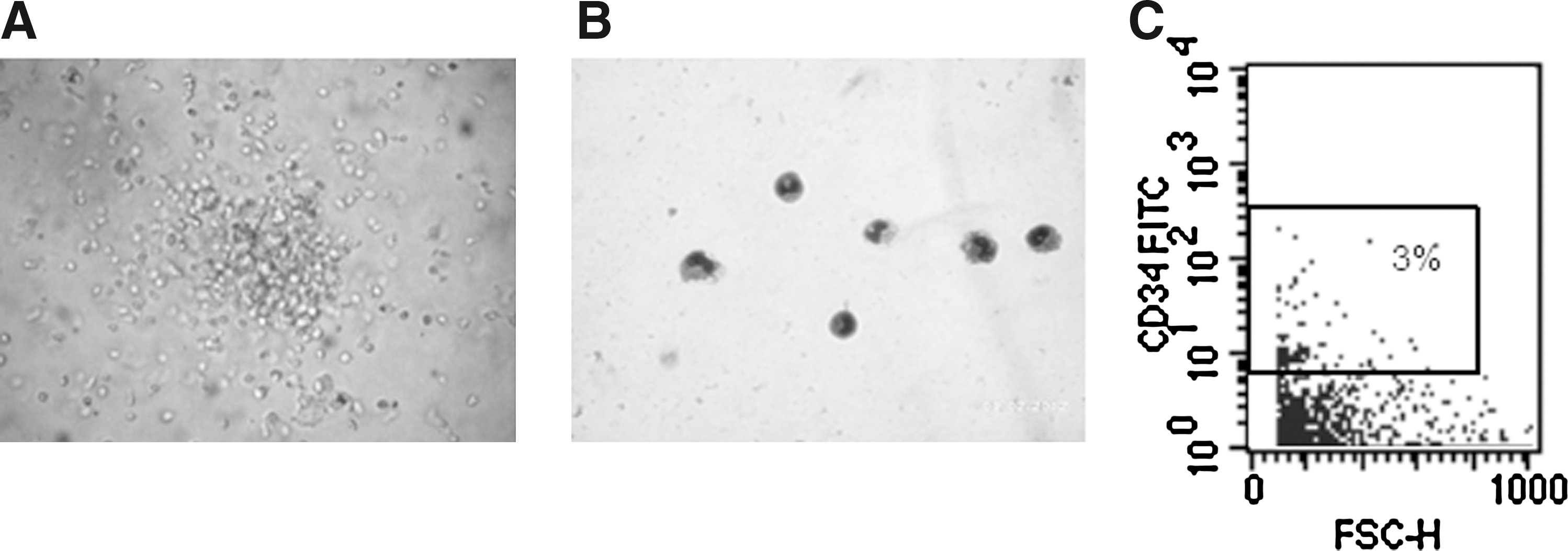
Detection of CFU-GM progenitors in BMC/OM-MSC co-cultures. NACs were collected from BMCs/OM-MSCs, after 17 days of co-culture, and colony-forming assays were performed. Microscopic evaluation shows the presence of a CFU-GM colony
Expression of hematopoietic cytokines by OM-MSCs
Based on results presented earlier that OM-MSCs support in vitro human hematopoiesis from BMCs, we investigated whether these cells express hematopoietic cytokines. OM-MSCs express mRNA messages for the early acting cytokine SCF, which exerts its effect on primitive hematopoietic progenitors (Fig. 6). They also express messages for GM-CSF, IL-6, and IL-11, which act on more committed hematopoietic progenitors (Fig. 6). Additionally, mRNA message was found for the lymphopoietic cytokine IL-7 (Fig. 6). Together, these results demonstrate that OM-MSCs constitutively express hematopoietic cytokines at the mRNA level.

OM-MSCs constitutively express hematopoietic cytokines. Reverse transcription and amplification were performed as described in the Materials and Methods section using specific primers for SCF, GM-CSF, IL-6, IL-11, and IL-7.
OM-MSCs maintain their mesenchymal phenotype after long-term co-culture
We evaluated the possibility that OM-MSCs change their phenotypical characteristics because of co-culturing with BMCs. Flow cytometric analysis of OM-MSCs showed that a high percentage of these cells maintain the expression of the MSC marker CD90 on their surface (Fig. 7), after long-term culture.
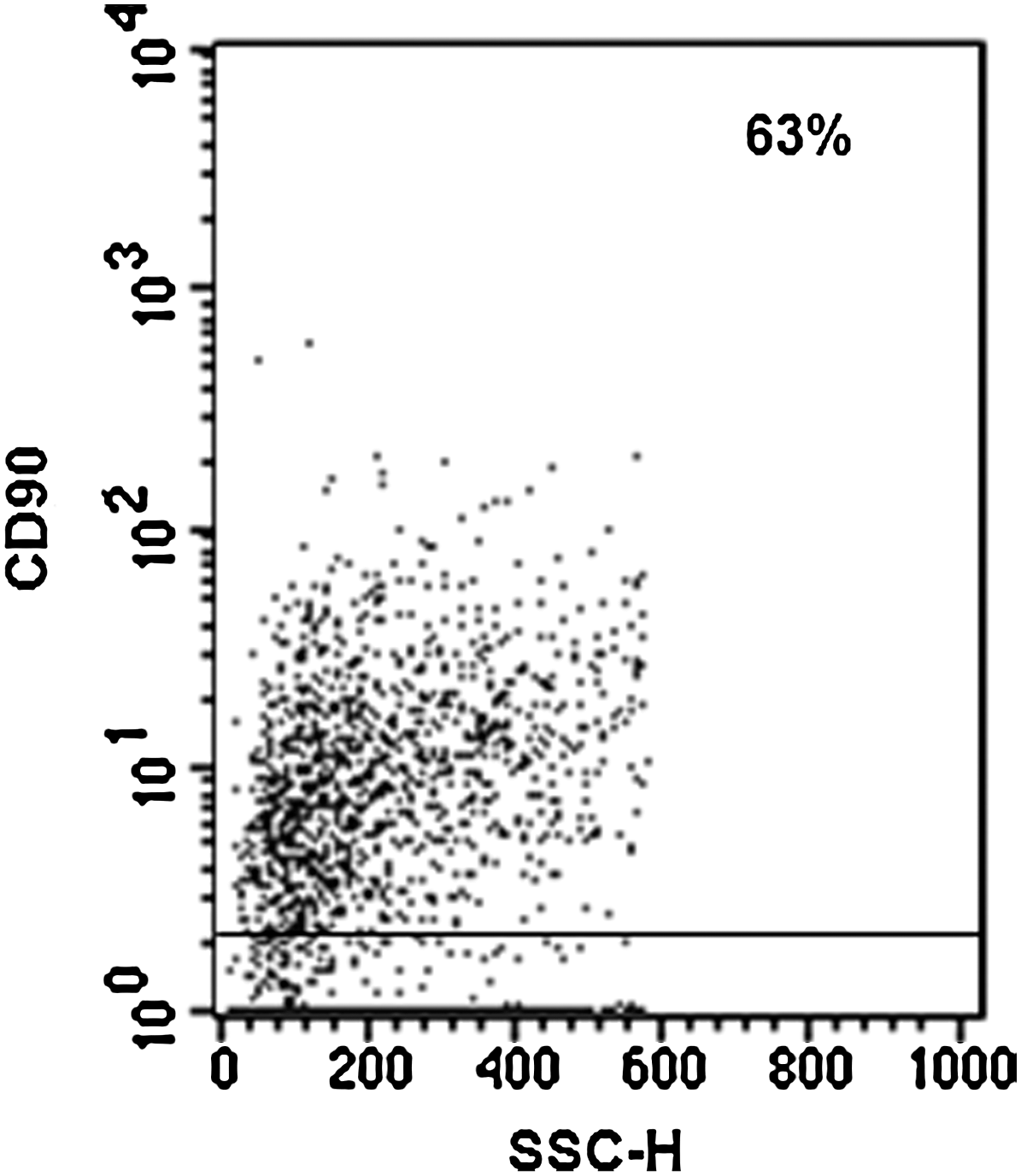
OM-MSCs maintain mesenchymal phenotype after long-term co-culture with BMCs. BMC/OM-MSC co-cultures were harvested after 54 days of co-culture, and the cells were analyzed for their expression of the CD90 MSC marker by flow cytometry. Negative controls were stained with the respective isotype (gray profile). Histogram analysis demonstrates that OM-MSCs maintain the CD90 expression on their surface. The results correspond to 1 experiment representative of 2 different assays, all with similar results.
Discussion
It is well known that BM stromal cells play an important role in the regulation of hematopoiesis [41 –43]. There is evidence indicating that the ability of BM stromal cells to support hematopoiesis is due to their capacity to produce numerous hematopoietic cytokines and express adhesion molecules, both implicated in different aspects of hematopoiesis [1]. Numerous reports have shown the capacity of MSCs from BM to support the survival, proliferation, and differentiation of HSCs, in vitro. Although BM is the most well-known source for MSCs, there is evidence showing the presence of these cells in other tissues, and their capacity to maintain in vitro hematopoiesis [44 –47]. Recently, MSCs from the human OM have been characterized [37], and it has been suggested that these cells might constitute a fundamental element of the neural stem cell niche in the human OM [36]. Because OM-MSCs share similar phenotypical and multipotential characteristics with MSCs from BM [39], in this work, we investigated the capacity of human OM-MSCs to support the survival, proliferation, and differentiation of human HSCs, in vitro. We show that OM-MSCs promote hematopoiesis from human BM hematopoietic cells.
In this work, we provide evidence that OM-MSCs promote myeloid differentiation from BMCs. High percentage of cells (73%) expressing the leukocyte common antigen CD45 were detected in the NACs of the BMC/OM-MSC co-cultures. Cytological analysis showed the presence of cells belonging to the myeloid lineage with variable morphology, including neutrophils, monocytes, basophils, and eosinophils. These results support previous studies showing that BM-MSCs and other stromal cell lines promote hematopoietic differentiation to myeloid lineages [48 –53]. However, most of these studies were performed using co-culture systems supplemented with exogenous cytokines [46,51,52,54,55]. In our study, OM-MSCs promoted myeloid differentiation in the absence of exogenous cytokines. It may be explained by the capacity of OM-MSCs to constitutively express hematopoietic cytokines that promote myeloid differentiation, such as GM-CSF. Importantly, the percentage of cells expressing the CD45 marker, in co-cultures with other stromal cell lines, were very low (0.1%–2% of CD45+ during 8–20 days of culture), as compared with those generated in co-cultures with OM-MSCs (73% of CD45 cells). Taken together, these results indicate that OM-MSCs promote myelopoietic differentiation from human BM hematopoietic cells, and provide the first evidence that OM-MSCs constitute a myelopoietic in vitro microenvironment.
Although previous reports have shown that hematopoietic differentiation promoted by stromal cells is mainly to the myeloid lineage [48,56], there is also evidence showing that MSCs from BM may also promote B and T-cell development [57]. Here, we provide evidence that OM-MSCs promote not only differentiation to myeloid lineages, but also lymphopoiesis from BMCs. We detected T, B, and NK cells in the NAC fraction of BMC/OM-MSC co-cultures. Similar results have been reported using co-cultures of murine ESCs with OP9 cell line, in which T, B, and NK cells were found [49,58]. However, these co-cultures were supplemented with lymphopoietic cytokines and lymphoid differentiation was mainly to generation of B-lineage cells. Our results showing that OM-MSCs promote lymphopoiesis from BMCs, in the absence of exogenous cytokines, may be explained by the capacity of these cells to constitutively express the lymphopoietic cytokine IL-7. Recent reports have also shown a restricted generation of B-lineage cells without using exogenous cytokines [59]. Other MSCs lines reported to promote hematopoiesis, in vitro, do not have the capacity to support lymphopoiesis in vitro [51]. Together, our results show for the first time that OM-MSCs constitute an in vitro lymphopoietic cell microenvironment.
Our results show that OM-MSCs express a wide array of hematopoietic cytokines, including the early acting cytokine SCF, which is known to maintain HSCs in quiescence or promote their self-renewal or differentiation [60,61]. Additionally, OM-MSCs express late-acting cytokines, such as GM-CSF, IL-6, IL-11, and IL-7, which are known to play an important role in various stages of myeloid and lymphoid differentiation [58,62]. The expression of other growth factors by OM-MSCs has been also previously reported [37,63].
In our co-culture system, we also found that OM-MSCs maintain their mesenchymal phenotype after long-term culture, expressing high levels of molecules characteristics of MSCs, such as CD90. This is very important because it has been reported that HSCs need to be re-plated into newly fresh feeder layers, to prevent transformation due to ageing of stroma.
In summary, our work provides evidence that OM-MSCs are capable to maintain in vitro hematopoiesis, in long-term cultures. The promoting effect of OM-MSCs is not limited to any specific hematopoietic lineage, because it involves myeloid and lymphoid cells, indicating that these cells may provide an excellent ex vivo microenvironment for hematopoietic progenitor expansion. The ability of OM-MSCs to support in vitro hematopoiesis could be related to the fact that OM-MSCs express a variety of hematopoietic cytokines.
Footnotes
Acknowledgments
This work was supported by IVIC and FONACIT grant G2005000405.
Author Disclosure Statement
The authors declare that they do not have any conflict of interest to disclose.