
Abstract
High expectations surround the area of stem cells therapeutics. However, the cells' source—adult or embryonic—and the cells' origin—patient-derived autologous or healthy donor genetically unrelated—remain subjects of debate. Autologous origins have the advantage of a theoretical absence of immune rejection by the recipient. However, this approach has several limitations with regard to the disease of the recipient and to potential problems with the generation, expansion, and manipulation of autologous induced pluripotent stem cells (iPS cells) preparation. An alternative to using autologous cells is the establishment of a bank of well-characterized adult cells that would be used to generate iPS cells and their derivatives. In the context of transplantation, such cells would come from genetically unrelated donors and the immune system of the recipient would reject the graft without immunosuppressive therapy. To minimize the risk of rejection, human leukocyte antigen (HLA) compatibility is certainly the best option, and the establishment of an HLA-organized bank would mean having a limited number of stem cells that would be sufficient for a large number of recipients. The concept of haplobanking with HLA homozygous cell lines would also limit the number of HLA mismatches, but such an approach will not necessarily be less immunogenic in terms of selection criteria, because of the limited number of HLA-compatible loci and the level of HLA typing resolution.
Introduction
S
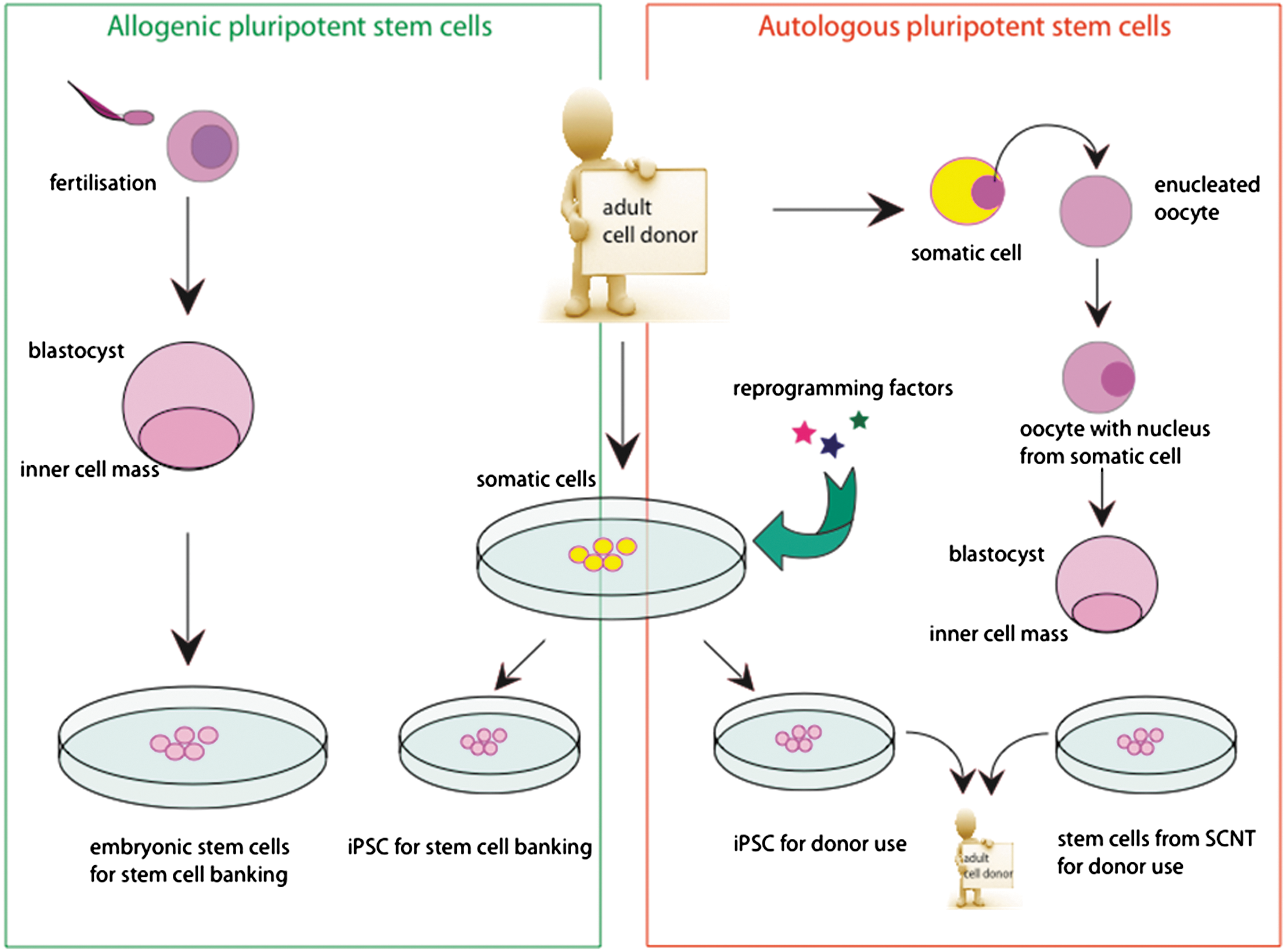
Stem cell grafts can be categorized into 2 types. Cells being grafted on the same individual from whom they were obtained form an autologous graft. Such a graft can be produced with induced pluripotent stem (iPS) cells or with stem cells from somatic cell nuclear transfer (SCNT). An allogenic graft on the other hand is formed of stem cells obtained from a genetically diverse individual, for example acquired in a stem cell bank. Such cells can be iPS cells or human embryonic stem cells.
Human ES cells have a variety of characteristics that make them an excellent tool to generate tissue and organs. The most important attribute of ES cells is their pluripotency: they can differentiate into all 3 germ layers and are thus a source for all types of differentiated cells [2]. Another advantage is their ability to proliferate indefinitely in the appropriate medium [2]. Due to these qualities, ES cells are seen not only as a potential cure for various diseases and injuries such as Parkinson's, diabetes, spinal cord injury, and arthritis, but also as important for transplantation and regenerative medicine.
Human ES cells research has stirred some controversy and for some, their use is a major ethical issue, as it requires the use of embryos to produce cell lines. However, the 2006 discovery that it is possible to reprogram differentiated cells and produce iPS cells has brought a new alternative without such ethical concerns [3].
Although their potential is considerable, there are still many challenges ahead before clinical use of pluripotent stem cells becomes a reality. The main focus of this review will be the possible immune reactions to pluripotent stem cell transplantation, strategies to counteract graft alloreactivity, and the potential of pluripotent stem cell banks for clinical applications.
Immunogenicity of Pluripotent Stem Cells
Transplantation of cells, tissues, or organs from a donor will be associated with a high risk of graft rejection, except between identical twins or if the immune system is inhibited by immunosuppressive agents.
Graft rejection is caused by allorecognition: the host immune cells, the transplanted organ, tissue, or cells as nonself. These types of immune reactions have been categorized according to their time of occurrence as hyperacute (minutes, hours), acute (days, weeks) or chronic (months, years) [4]. Chronic rejection has multiple causes, not all of which are mediated by the immunogenic system [5]. Hyperacute rejection is characterized by the presence of preformed antibodies specific to donor antigens, which directly bind the graft without any further need to stimulate the immune system [4]. The typical example of hyperacute rejection is the reaction between incompatible blood groups between donor and recipients due to the anti-ABO antibodies of the recipients. Although protocols of ABO incompatible transplantation have been developed [6,7], blood group compatibility is strongly recommended in solid organ transplantation. In addition to ABO groups, hyperacute rejection can also ensue if there has been prior exposition to the nonself-human leukocyte antigen (HLA) type of the donor, through blood transfusion, pregnancy, or previous transplantation. When considering the use of pluripotent stem cells or their derivatives for transplantation, hyperacute rejection can be prevented through testing for the presence of recipient anti-HLA antibodies directed against the donor tissue. ABO compatibility is not mandatory in hematopoietic cell transplantation [8], but progenitors and mature cells or tissues derived from pluripotent stem cells could express ABO antigen; therefore, it seems reasonable to consider blood group compatibility. Acute rejection is driven by the activation of specific allogenic T cells by antigen-presenting cells (APCs). In organ transplantation, APCs can be of donor and recipient origin. These 2 types of APCs are able to activate T cells through 2 different pathways: direct and indirect [9] (Fig. 2). Direct recognition is the result of the donor APCs stimulating the recipient T cells, while indirect recognition occurs when a recipient APC presents a donor peptide. The direct pathway is mediated by HLA incompatibility between the donor and the recipient leading to HLA allorecognition and recipient T cell activation [10]. In solid organ transplantation, this pathway is driven by donor APCs present in the graft that can migrate into the recipient lymph nodes to activate T-lymphocytes. The indirect pathway results from graft infiltration by recipient APCs. Those APCs migrate to the lymph nodes to present donor antigens to activate T lymphocytes.
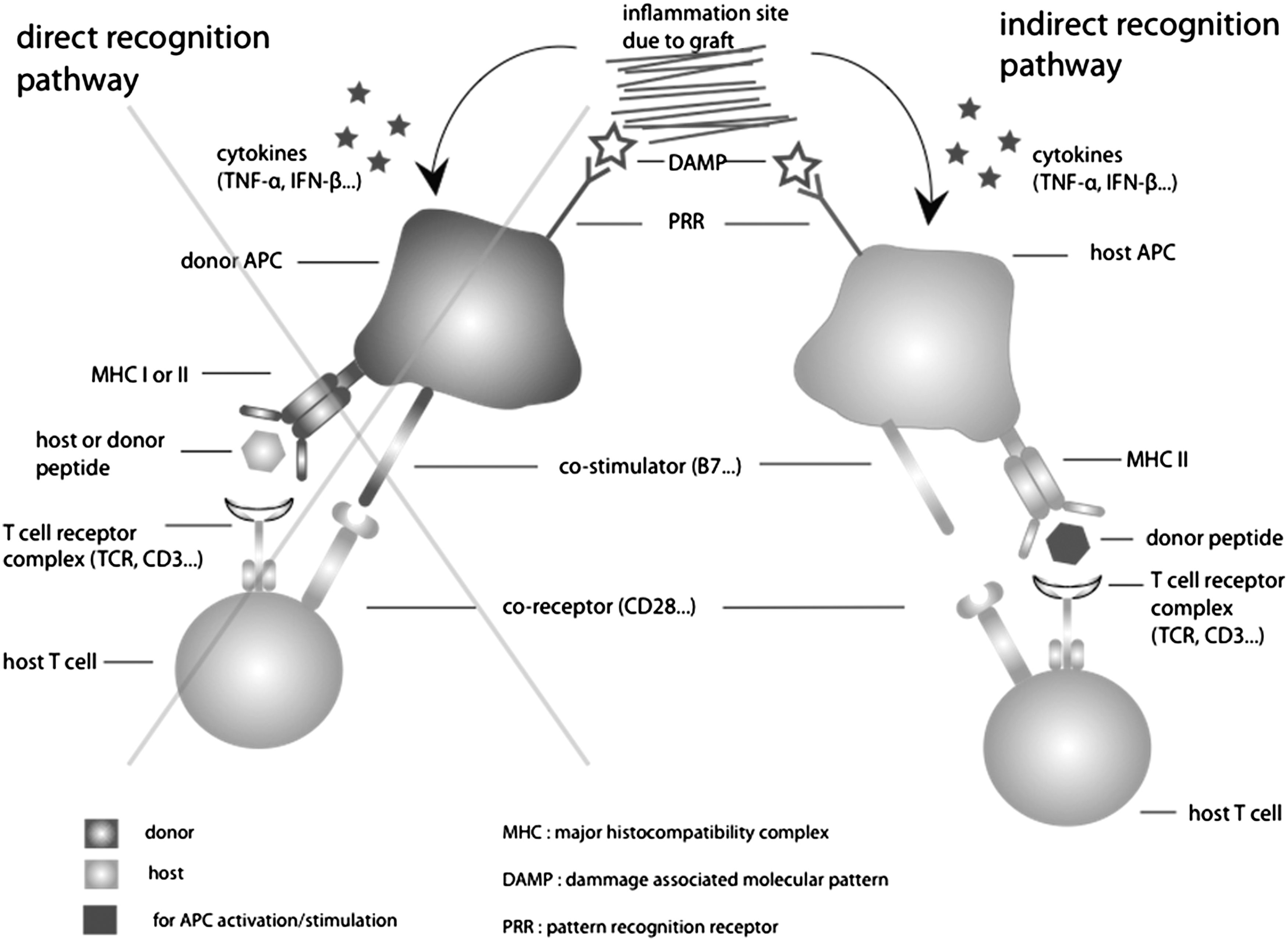
Graft-induced inflammation triggers an activation of antigen-presenting cells (APCs) through cytokine expression and DAMPs (damage-associated molecular pattern molecules). These APCs will then present intracellular peptides through their major histocompatibility complex (MHC) I, or extracellular peptides through their MHC II, to naïve host T-cells and thus activate them (with costimulation). In indirect recognition, the T-cell recognizes a donor peptide [most probably a human leukocyte antigen (HLA) fragment, as they are highly polymorphic] as nonself and is thus activated. In direct recognition, the donor MHC I/II plus a peptide (donor or host) are together recognized as nonself. In pluripotent stem cell transplantation grafts, the probability of having donor APCs is minute and thus the mechanism of rejection will be mediated by an indirect pathway only.
With cells and tissue derived from ES cells or iPS cells, the mechanism of rejection will be stimulated mainly by the indirect pathway, unless APC cells are generated from the pluripotent ES or iPS cells. Also, direct allorecogonition due to cross-reactivity of a virus-specific memory T cell cannot be excluded [11].
Therefore, the role of HLA compatibility between donor and recipient, which is associated with better survival of the graft in solid organ transplantation and also in hematopoietic stem cells transplantation [12], is questionable in the case of cells or tissue derived from pluripotent stem cells that normally will not generate APCs. However, HLA antigens are strongly immunogenic and can be presented as minor antigens to T cells and, as mentioned above, lead to the development of anti-HLA antibodies. HLA class I antigens will be expressed by any cell of the graft and HLA class II will be expressed following inflammatory stimuli induced by dead or dying cells. Thus, we believe that HLA compatibility remains a suitable objective to reduce the risk of rejection following transplantation of cells or tissue derived from pluripotent stem cells. In addition, as soon as more complex tissue will be generated, the possibility of having cells with antigen-presenting capacity would significantly increase.
The use of immunosuppressive drugs is mandatory for nearly every type of solid organ transplantation, even with full HLA matching such as observed in HLA genotypically identical siblings. This is due to the presence of minor antigen mismatches, which differ between any individuals except identical twins and have the potential to activate T-cells through the indirect pathway (the recipient APC presents a donor allogenic minor antigen) (Fig. 2). In most cases, compatibility for all HLA antigens is extremely difficult to achieve due to the large extent of allelic polymorphisms in the HLA system [13].
It has long been suggested that ES cells and their derivatives might be immune privileged, and that grafts with such cell types would produce only a very weak immune response, if any. This hypothesis seemed confirmed by the very low expression of HLA class I antigens (which appeared only weakly increased through differentiation), and by the lack of expression of HLA class II [14,15]. However, recent publications have contradicted these assumptions and demonstrated that human ES cells, as well as their derivatives, induce an immune reaction in T and NK cells [16 –18], suggesting a problematic immune reaction to ES cells engrafts. A lower immune reaction of ES cells and differentiated cells derived from ES cells could be of interest because energy or tolerance could be induced and be of great benefit for stem cell therapy [19,20].
Before clinical trials in stem cell therapy can be considered, it is necessary to comprehensively study immune reactions to stem cell grafts and to develop therapeutic strategies. Since HLA mismatch plays a central role in graft rejection, using HLA compatible stem cells would limit the immune response.
HLA System and HLA Haplotype
The classical HLA system is a multigenic and multiallelic system that codes for the cell surface antigens, HLA class I (loci A, B, and C) antigens, which are composed of a heavy chain that pairs with β2-microglobulin, and HLA class II (DR, DQ, and DP) antigens that are heterodimers formed by an α-chain and a β-chain. The HLA region thus comprises 3 HLA-A, B, and C genes and 6 HLA class II genes, DRA/DRB1, DQA1/DQB1, and DPA1/DPB1. Some individuals express a second DR antigen that is encoded by one of the DRB3, DRB4, or DRB5 genes. The total number of HLA antigens expressed by a given individual ranges between 12 and 14 because of the codominant expression of HLA antigens. The classical HLA system is highly polymorphic to present a large variety of peptides to T lymphocytes; the major part of this polymorphism resides in the exons encoding the peptide-binding site. Classification of these polymorphisms is defined according to the technology that has been developed to discriminate the HLA molecule and follows strict rules that are internationally recognized. Historically, the first classification of HLA diversity was based on the use of allo-antisera that was able to recognize cell-surface HLA antigens in a test called complement-dependant microlymphocytotoxicity, or more commonly serological typing. Approximately 100 HLA class I and II specificities, or serotypes, are now routinely discriminated, representing the so-called low resolution HLA typing level. The progress in the molecular technology led to the use of a molecular analysis to define the HLA polymorphism and rapidly showed that HLA polymorphism was much greater than expected, so that each serotype now comprises a high number of alleles that differ by one or just a few nucleotides. These alleles are classified within a given serotype because they share determinants that are recognized by allo-antisera; importantly, though, they most frequently differ in the peptide binding site and thus have the potential to affect peptide presentation and T-cell alloreactivity. This new high resolution HLA typing based on sequence information leads to an enormous increase in the HLA allelic polymorphism, with a continuously growing number of alleles being discovered (Fig. 3). Therefore, HLA compatibility at a low resolution level does not mean compatibility at the allelic level in most patient/donor combinations. Because of the HLA diversity described above, finding HLA compatibility between recipient and donor is very challenging. HLA-A, B, and DR compatibility based on a low resolution level is recommended in solid organ transplantation and is at least partially achieved depending on organ exchange programs. On the other hand, HLA compatibility based on high resolution typing for all HLA loci is practically impossible to achieve. Fortunately, the distribution of HLA alleles is not random and is at least segregated by haplotypes. A haplotype is a set of alleles encoded by a group of closely linked genes, which are usually inherited as a unit, an individual inheriting a complete haplotype from each parent. These haplotypes can be used to trace migrations of human populations because they often represent a type of fingerprint of important events in human evolution. As an example, A*01:01, C*07:01,B*08:01,DRB1*03:01 (A1-B8-DR3) is a frequent haplotype in Western European populations, which declines along gradients away from that region. Despite the very large allelic diversity of the HLA system, the occurrence of a limited number of haplotypes due to linkage disequilibrium of the HLA region present in a given population makes HLA compatibility an option even at high resolution typing [21].

Low resolution and high resolution HLA typing defined the nomenclature at the antigen or alleles level. High resolution typing by molecular biology leads to a high number of alleles, which increase the polymorphism and render HLA compatibility between donor and recipient difficult to achieve at this level of resolution.
Pluripotent Stem Cell Banking
Having established that HLA matching is associated with a reduced immune reaction against pluripotent stem cell grafts, the question remains how to obtain such compatible stem cells. Current research can be divided into 2 main approaches: patient-specific pluripotent stem cells and stem cell banking [22].
Some other possibilities are also being pursued, such as inducing mixed chimerism to produce tolerance toward donor cells, but this method relies mainly on the thymic activity, which is significantly less efficient in adult individuals [23]. Genetic modification to produce a universal donor cell has also been put forward as a solution, but more research is needed on the consequence of such manipulations before it can be further considered [24]. All of the aforementioned solutions might have the potential to eliminate the immune reaction or at least decrease it considerably. However, they are currently still in the research stage and serious technical challenges will likely remain in the foreseeable future.
It seems obvious that the use of patient-specific stem cells would result in complete-HLA matching; nevertheless, new insights discussed below seem to question this assumption, which needs to be examined further. iPS cells or somatic cell nuclear transfer (SCNT) are thought to be 2 solutions where one could create a genetically identical and perfectly HLA matched cell line for any recipient [25], but neither of these procedures can be considered on a clinical level yet. SCNT on human cells poses major ethical and technical issues, and even though it would produce a perfect match for any nuclear antigens, the mitochondrial antigens would still be mismatched, the consequence of which is not yet clear.
There are a number of methods to produce iPS cells from adult somatic cells, which will briefly be described before addressing the immunogenicity of these cells. One of the most common and efficient methods to produce iPS cells seems to be through viral expression of a certain number of reprogramming factors (OCT4, SOX2, c-MYC, etc.), for which various protocols exist [26]. Another possibility is plasmid transfection of these same reprogramming factors, the advantage being no integration of DNA into the host genome, but this method seems to offer a relatively low yield [27,28]. Finally, a recent protocol for induction of pluripotency using a small molecule has proven that the reprogramming factors, thought essential in the beginning of iPS cell research, may in fact be replaced by molecular activation of the reprogramming pathways [28,29].
It is often thought that autologous tissue transplant-derived iPS cells reprogrammed from patient somatic adult cells (patient-derived iPS) would eliminate the need for immunosuppressive therapy [30]. However, the use of patient-derived iPS cells still has several important limitations to consider. As mentioned above, viral vector transduction technology to reprogram adult cells is very efficient, but it has many drawbacks when considering the clinical application of iPS cells. If the viruses used to reprogram the cells are integrating the genome this may cause genetic defects, but even with nonintegrating viruses like adeno-virus the high expression of the reprogramming factors presents a risk in itself, as some are oncogenes [27,28]. Thus, the mechanisms underlying induction of pluripotency still need to be studied further so that new protocols producing more clinically useful iPS cells may be established; first steps in this direction have been made with the new molecular protocols.
Another disadvantage when considering iPS cells is that treatment of genetic disease would not be feasible using adult cells from sick patients without definitive genetic correction before derivation and transplantation. This step would certainly complicate the procedure and would require optimization of the procedure for each genetic defect, leading to unacceptable delay. Furthermore, the age of the donor could be a problem because of the reduced capacity for reprogramming adult cells [31]. Recent publications have indicated that iPS cells show a high risk of chromosomal and mitochondrial DNA mutations, the chromosomal mutations were observed frequently at locations highly enhancing the probability of tumor formation [32,33]. Even though it seemed that the mitochondrial mutations catalogued in most iPS cells did not affect their pluripotency, disease-inducing mutations are still a possibility, and mitochondrial DNA would have to be closely monitored in iPS cells destined for clinical use [33]. Thus, iPS cells would need to be tested regularly for DNA mutations and stability, a further technical burden in this type of pluripotent stem cell production. Finally, the ability to work in good medical or laboratory practice conditions at reasonable costs also must be addressed.
Even so, the technical difficulties mentioned above would surely be manageable if they brought about absolute immune tolerance in the form of iPS cells. However, recent findings seem to indicate that these cells do induce an immune reaction even in an individual genetically identical to the one from which they were derived, as seen in transplantation to syngeneic mice [34,35]. This reaction seems to be due to in part to a prolonged in vitro culture of iPS cells and a consequential expression of immunogenic markers, as well as an expression of developmental genes that could be immunogenic when in an environment in which they are normally lacking [35]. In fact, adult somatic cells express antigens specific to their identity, while during reprogramming iPS cells express specific developmental markers. Thus, these cells, after re-differentiation and re-implantation in a different environment, may still express these specific markers that could lead to an autologous immune reaction. In the context of patient-derived iPS, this immune reaction against autologous tissue could lead to a lesion that should be considered as an autoimmune phenomenon and finally to autoimmune disease.
The expression of silent neo antigen-like tumoral markers was also observed, producing an immune reaction [36], and it could be linked to the genetic insertions used for viral iPS cell production as mentioned above. Thus immunogenicity of iPS cells and their derivatives needs to be revaluated in full, and their induction protocols as well as in vitro culture conditions should be optimized in regard to the risk of immunogenic and tumoral marker expression.
Thus, it seems apparent that these types of patient-specific manufacturing of stem cell lines are difficult at both the technical and financial level. They are also time consuming and would be of little use in an emergency situation. Moreover, current research indicates they do not represent the perfect solution for avoiding immune rejection they were initially thought to be.
Another possibility is to create stem cell lines specific to the patient by deriving stem cell lines from parthenogenetic embryos. This has been successfully done [37], but the possibilities provided by the use of such cells still need to be studied further. These cells can only be produced for female patients, so would only be of use to half of the patients. In addition, in some countries, this type of cell derivation is forbidden by the governmental legislation.
Using iPS cells reprogrammed from adult cells coming from genetically unrelated donors, who have been validated through the critical processing steps, could be a more realistic alternative. The establishment of a panel of iPS cells derived from well characterized donors representative of the diversity of a given population has to be carefully evaluated.
Due to the high diversity of HLA genes, a perfect compatibility between cells and any recipient will rarely occur, although even a partial match might result in a decreased risk of immune rejection. As partial HLA matching is far from completely preventing graft rejection, immunosuppressive drugs will still be needed, as is the case in solid organ transplantation. Therefore, further research to evaluate the effect of the diverse drugs on ESC differentiation is necessary before considering clinical trials [17]. Incomplete HLA matching between stem and recipients is the main limitation of stem cell banking when compared with patient-specific lines, but the consequence of mismatching for multiple minor histocompatibility antigens also needs to be evaluated. Still, these are counterbalanced by the technical and financial viability of such a facility, which could provide cells in a timely manner; in pressing circumstances, there would be off-the-shelf stem cells.
In fact, stem cell banking is already a reality in countries such as the United Kingdom and Spain. Banks and registries of the different existing stem cell lines are a useful tool for stem cell research [38]. Unfortunately, even though some of these banks already have facilities for storing clinical grade pluripotent stem cell lines, they remain unused. Creating a stem cell bank for medical, rather than solely research, purposes is not simply a question of adding clinical grade iPS cell lines or human ES cell lines; there are many issues that still need to be solved.
Before addressing the issues around the facility per se, it is important to acknowledge the difficulties behind producing clinical grade stem cell lines. To be used in regenerative medicine, the cells need to be cultivated on a medium free of animal substances and comply with strict manufacture requirements [39]. These regulations apply starting with the cells' derivation from blastocysts (human ES cells) or reprogramming for iPS cells and also apply to the storage of the stem cell lines once they have arrived at the bank. This means that any facility wanting to distribute clinical grade stem cell lines must be able to certify their origin and storage conditions. This is just one of the many technical difficulties a stem cell bank for clinical grade ES cells and iPS cell lines will face [40].
The technical difficulties are not irrelevant, but they may be easier to resolve than the ethical problems of such a cell bank [41]. As stated above, the purpose of a stem cell bank containing different clinical-grade stem cell lines would be to permit HLA matching for transplantation. This means that it must contain different stem cell lines with a wide variety of HLA alleles and the necessity of such diversity seems to call for a global stem cell bank. Since its beginning, ES cell research was opposed by some parts of the population. Thus, the regulations surrounding the use of hES cells differ from country to country [38] and it is likely that the generation of clinical grade iPS cells will also be regulated in the near future. Therefore, globalization of stem cell banking will be difficult.
Another ethical concern of stem cell banking is the challenge of ethnic discrimination. The frequency of different HLA haplotypes differs enormously from one population group to another [42,43]; in countries such as the United States or the United Kingdom, a bank will likely contain haplotypes that are a match for the majority of individuals of European origin, but will contain few that match groups of different geographical origins. Again, this problem would call for a global stem cell bank [44].
This is the major challenge of creating a human pluripotent stem cell bank: it would have to contain clinical grade cell lines with matching HLA-A-B-DR haplotypes for a large portion of the patient population, without ethnic discrimination. The question of how many lines this would necessitate and of how to obtain them has been the focus of many studies, but there is still no clear answer. For human ES cells, a significant number of embryos should be analyzed to cover the most frequent haplotype from which to derive new cells; to accomplish this, it may be necessary to make embryo donation from IVF treatment mandatory under certain circumstances or by offering monetary compensation for embryos [44]. Homozygous stem cell lines could also be produced through parthenogenesis, but such cell lines seem to be genetically unstable [45]. Most of these suggestions raise ethical concerns and would be difficult in practice because they often go against public opinion [41,46]. A more feasible initiative would be to make use of the numerous cord blood banks already established in many countries [47,48]. These could be an appropriate source for iPS cells as they are already HLA typed and are probably representative of the HLA distribution in local population. Another advantage of cord blood is that mutation rates could likely to be lower than other sources such as fibroblasts. In the end, the key question is whether there is a need for such measures; in other words, how many human ES cells lines are actually required to create a pluripotent stem cell bank? The answer to this question naturally depends on how good a HLA match is needed for pluripotent stem cell transplantation, but since there are no clinical trials yet, there is no clear answer to this question. Thus, most studies on the subject calculated the number of lines needed for different levels of HLA matching and different haplotypes according to the population targeted.
Several studies have addressed this question and the data are summarized in Table 1. Using the haplotypes of organ donors, Taylor et al. [49] have estimated the matching (HLA-A, -B, and-DR at a low resolution level) of hypothetical lines, with patients waiting for an organ transplant in the United Kingdom. According to their study, 150 stem cell lines would provide a full match for 20%, one mismatch (HLA-A or HLA-B) or better for 37.9% and an HLA-DR match only or better for 84.9% of the patients. Extending the number of lines above 150 did not change the number of matches significantly [49]. Another study by Nakajima et al. [50] estimated the number of lines by using a group of unrelated individuals living in or near Tokyo as the donor and recipient pool. According to their results, 170 different ES cell lines would provide a single mismatch or less for 80% of the population (HLA-A, -B, and -DR typing performed at a low resolution). The difference between the 2 studies most likely results from the lower allelic and haplotypic HLA variability of the Tokyo population compared to that of the entire United Kingdom population. A similar study done in China by Lin et al. [51] established real ES cell lines to be used for transplantation. The prospective recipients were taken from the Hunan population, which is composed mostly of the Han ethnic group that represents about 94% of the Chinese population [52]. 174 lines resulted in a perfect match for 24.9% of the population (at the HLA-A-B-DR low resolution level), similar to the results of Taylor et al. Another study in Korea reported that only 27 cell lines would give 2 mismatches or better for 16% of the population [53].
It is important to note that these studies use mostly low resolution matching.
These lines being in most cases hypothetical.
The hypothetical patient populations are mostly taken from national donor registries.
HLA, human leukocyte antigen.
From these 4 studies, one can make the general statement that with less than 2 hundred different ES cell lines one should be able to offer advantageous HLA-A, -B, and -DR matches for a substantial fraction of the population (the meaning of advantageous for pluripotent stem cell transplantations still being left open for discussion). Still, this number will vary depending on the country; in the United States, for example, there is extreme heterogeneity of HLA haplotypes between different population subgroups and the number of lines currently available would be not sufficient to consider their use based on HLA matching for a large portion of patients, not to mention that they are not of clinical grade [54]. As suggested by the above studies, it is not yet possible to set up a bank with enough cell lines to provide HLA-A, -B, and -DR low resolution matches for a large majority of a given population. Of note, the degree of HLA typing in the above studies is performed at a low resolution (serological nomenclature). High resolution HLA compatibility would greatly augment the complexity of the system. Among the few possibilities to reduce the number of lines needed, we will focus on the use of homozygous hES cell lines.
Homozygous Stem Cell Lines
A homozygous human ES cell DNA will contain only half the number of different HLA alleles of a completely heterozygous line (the maternal and paternal haplotypes being identical in a homozygous line). Thus, it will match any heterozygous patient expressing this reduced number of diverse HLA alleles on one of his haplotypes. Such a cell line is therefore expected to provide a match for a larger group of patient compared to heterozygous cell lines (Fig. 4).

Considering only the HLA-A allele, we represent here an example of a homozygous donor for a hypothetical patient with HLA-A1 and HLA-A3 alleles. A homozygous donor, with 2 HLA-A1 alleles, similar to the recipient HLA-1, will not induce T cells alloreactivity (left part). The heterozygous donor expressing the 2 different HLA-A alleles 1 and 2 can induce T-cells alloreactivity though the HLA-A2 allele that differ from the recipient HLA-A (right part).
In all of the previously mentioned studies, this obvious potential of homozygous stem cell lines has been analyzed. In the Lancet study, while 150 lines provided a perfect match for 20% of the population, it was noted that only 10 well-chosen homozygous lines could provide a perfect HLA-A, -B, and -DR match for 37.7% [49]. This was reiterated in the study by Nakajima et al. [50] where it was suggested that 55 randomly selected homozygous embryos could produce lines to provide a perfect match for 80% of the population. In the study on the Chinese population, it was noted that just 5 different homozygous haplotypes of their 174 stem cell lines contributed to 50%–80% of the different levels of HLA matching [51].
All these results indicate that the number of cell lines needed in a pluripotent stem cell bank, which would provide matches for a majority of the population, could be drastically reduced if it contained homozygous lines. The question that now arises is to know how feasible it would be to produce such lines. In the Lancet study, for 10,000 cadaveric organ donors, 145 were homozygous; if we assume that these donors are a good representation of the general population, one could say that 1.5% of the population is homozygous for HLA-A, -B, and -DR, but these lines represented only 26 different haplotypes [49]. In the study by Lin et al. [51], however, 8 of 174 different ES cell lines were homozygous, representing 5 different haplotypes. Such a high rate of homozygous lines is explained by the fact that they were derived from abnormal embryos; the rate in the healthy population is much lower (1.1%) [55]. Studies have addressed the surprisingly low number of homozygotes in the population [55]. It seems unlikely that a bank with lines obtained by deriving ES cells from randomly selected embryos would contain an important number of homozygous lines. To increase homozygous cell lines, several options could be proposed. Selection of ES cells coming from homozygous individuals that have frozen embryos would be theoretically possible, but ethically unacceptable. The use of embryos obtained through parthenogenesis is also a possibility, even though some parthenogenotes are heterozygous due to crossing-over [56,57]. Selection of homozygous individuals by looking at hematopoietic stem volunteer donor registries to derive cell lines through SCNT or by producing iPS cells is a more realistic option [58,59]. As explained above, SCNT is a method that still faces many technical and ethical difficulties and iPS cells might be a more reasonable prospect. Tamaoki et al. [60] used dental pulp cells, which are easily accessible from extracted teeth, to produce 5 lines of iPS cells. Having established that this was possible, they estimated that using only the 2 homozygous, out of the 107, lines of dental pulp cells to produce iPS cells would already provide a perfect match for approximately 20% of the Japanese population. In summary, the production of iPS cells from known homozygotes, which can be found through the registries or produced from homozygous cells found in cord blood banks, seems to offer the largest possibilities for proceeding on this direction.
Conclusion
The use of pluripotent stem cells in regenerative medicine has a bright future, but there are still serious obstacles, including the potential immunogenicity of pluripotent stem cells and their derivatives.
Although producing patient-specific stem cells through induced pluripotency seemed to resolve many immunological problems, this procedure is costly, laborious, and time-consuming. A solution in the near future could be pluripotent stem cell banking, modeled after today's cord blood banks, which would contain clinical grade stem cell lines. Such banks would provide different stem cell lines from which one could choose a HLA match for the patient.
There are ethical and technical difficulties behind the creation of such a bank, but they appear to be less than those for patient-specific lines. The most important factor that needs to be considered is the number of lines needed so as to create a bank that would provide a favorable HLA match for the majority of the population, and how to reduce that number. Current literature on the subject indicates that to have human ES cell lines of the most common HLA haplotypes and to have homozygous lines has a major impact on the number of lines needed. Producing such lines by SCNT or by induced pluripotency is a possibility, as these procedures would be financially and technically worthwhile if one line can be used for a large number of patients.
Although the conception of such a bank would enable a certain level of HLA matching, as with other transplantations, the probability of a complete match of the entire major histocompatibility complex is unexpected. Even in the case of perfect HLA-matching at a high resolution between the donor cell line and the patient, the non-HLA genetic difference between these 2 individuals would lead to rejection due to the presentation of histocompatibility minor antigens by the recipient immune system.
Therefore, protocols for clinical trial would include immunosuppressive drugs. The effect of immunosuppressive drugs on hES cells, iPS cells, and their derivates has not been extensively studied and requires further evaluation.
In summary, a pluripotent stem cell bank does not eliminate the risk of an immune reaction after pluripotent stem cell transplantation, but is a promising solution to decrease the risk of rejection by the immune system. The establishment of multiple regional banks, accounting for the variability of HLA haplotypes in different human populations, should be strongly considered to advance medical and research purposes.
Footnotes
Acknowledgments
This work was supported by the FNRS (Synergia grant for the Prometheus project). The authors thank Katharine Brieger for critical reading of the article.
Author Disclosure Statement
All authors disclose no conflict of interests with regard to the content of this article.