
Abstract
Embryonic stem (ES) cell- and induced pluripotent stem (iPS) cell-derived hematopoietic stem/progenitor cells (HSPCs) are considered as an unlimited source for HSPC transplantation. However, production of immature hematopoietic cells, especially HSPCs, from ES and iPS cells has been challenging. The adaptor protein Lnk has been shown to negatively regulate HSPC function via the inhibition of thrombopoietin (TPO) and stem cell factor signaling, and Lnk-deficient HSPCs show an enhanced self-renewal and repopulation capacity. In this study, we examined the role of Lnk on the hematopoietic differentiation from mouse ES and iPS cells by the inhibition of Lnk using a dominant-negative mutant of the Lnk (DN-Lnk) gene. We generated mouse ES and iPS cells stably expressing a DN-Lnk, and found that enforced expression of a DN-Lnk in ES and iPS cells led to an enhanced generation of Flk-1-positive mesodermal cells, thereby could increase in the expression of hematopoietic transcription factors, including Scl and Runx1. We also showed that the number of both total hematopoietic cells and immature hematopoietic cells with colony-forming potential in DN-Lnk-expressing cells was significantly increased in comparison with that in control cells. Furthermore, Lnk inhibition by the overexpression of the DN-Lnk gene augmented the TPO-induced phosphorylation of Erk1/2 and Akt, indicating the enhanced sensitivity to TPO. Adenovirus vector-mediated transient DN-Lnk gene expression in ES and iPS cells could also increase the hematopoietic cell production. Our data clearly showed that the inhibition of Lnk in ES and iPS cells could result in the efficient generation and expansion of hematopoietic cells.
Introduction
S
Recently, inhibitors of differentiation (ID) genes, which are negative regulators of E proteins (E2A, HEB, and E2-2) [11], were shown to negatively regulate the hematopoietic differentiation in ES and iPS cells [12]. The same study also showed that the suppression of the ID genes, ID1 and ID3 increased the number of ES and iPS cell-derived hematopoietic progenitor cells [12]. These data indicate that negative regulators play an important role in the hematopoietic differentiation process in ES and iPS cells, and that manipulation of the expression of negative regulators would be an effective strategy for the efficient generation of hematopoietic cells from ES and iPS cells.
An adaptor protein Lnk/SH2B3 (hereafter referred to Lnk) is shown to negatively regulate the thrombopoietin (TPO) and stem cell factor (SCF) signaling, both of which are crucial cytokine-signaling pathways involved in hematopoietic stem cell (HSC) self-renewal, since Lnk-deficient HSCs exhibit an augmented response to TPO and SCF stimulation, and thereby Lnk-deficient mice show the marked HSC expansion in the bone marrow [13 –16]. In addition, Lnk is highly expressed in immature hematopoietic cells, particularly in HSCs [17], in contrast to ID genes, which are ubiquitously expressed in many tissues [11,18]. Therefore, we speculated that an inhibition of Lnk function in ES and iPS cells would lead to the efficient generation and expansion of immature hematopoietic cells. In the present study, we investigated the effects of Lnk inhibition on the hematopoietic differentiation of mouse ES and iPS cells, and we found that a suppression of Lnk function by the enforced expression of a dominant-negative mutant of the Lnk (DN-Lnk) gene in ES and iPS cells resulted in an increase in the number of both mesodermal cells with hematopoietic differentiation potential and immature hematopoietic cells. These findings indicate that the suppression of the Lnk would be useful for the efficient generation and expansion of ES cell- and iPS cell-derived hematopoietic cells.
Materials and Methods
Plasmid construction and adenovirus vectors
pEF-IRESneo, which contains internal ribosome entry sites (IRES) and a neomycin-resistant gene (Neo) downstream of the human elongation factor (EF)-1α promoter, was constructed by replacing the cytomegalovirus (CMV) promoter of pIRESneo (Clontech) with the EF-1α promoter, which is derived from pEF/myc/nuc (Invitrogen). Mouse DN-Lnk cDNA, derived from pMY-DN-Lnk [19], was inserted into pEFIRESneo, resulting in pEF-DNLnk-IRESneo. Adenovirus (Ad) vectors were constructed by an improved in vitro ligation method [20,21]. Mouse DN-Lnk cDNA was inserted into pHMCA5 [22], which contains the CMV enhancer/β-actin promoter with an β-actin intron (CA) promoter (a kind gift from Dr. J. Miyazaki, Osaka University) [23], resulting in pHMCA5-DN-Lnk. pHMCA5-DN-Lnk was digested with I-CeuI/PI-SceI and ligated into I-CeuI/PI-SceI-digested pAdHM4 [20], resulting in pAd-DN-Lnk. Ad-DN-Lnk and Ad-DsRed were generated and purified as described previously [24]. The CA promoter-driven β-galactosidase (LacZ)-expressing Ad vector, Ad-LacZ, and the CA promoter-driven DsRed-expressing Ad vector, Ad-DsRed, were generated previously [24,25]. The vector particle (VP) titer was determined using a spectrophotometric method [26].
Cell culture
The mouse ES cell line, BRC6 (Riken Bioresource Center), and the mouse iPS cell line, 38C2 (a kind gift from Dr. S. Yamanaka, Kyoto University) [27], were used in this study. DN-Lnk- or Neo-expressing mouse ES and iPS cell lines were generated as follows. The pEF-IRESneo and pEF-DNLnk-IRESneo were linearized and were then electroporated into mouse ES cells and iPS cells by using Gene Pulser Xcell (250 V, 500 μF; Bio-Rad Laboratory). pEF-IRESneo- or pEF-DNLnk-IRESneo-transfected ES cells and iPS cells were cultured in an ES cell medium containing 100 μg/mL G418 (for ES cells) or 200 μg/mL G418 (for iPS cells) for 10–14 days, and G418-resistant colonies were picked up and expanded. The expression of DN-Lnk was confirmed by conventional reverse transcription–polymerase chain reaction (RT-PCR). Mouse ES cells, iPS cells, and Neo- or DN-Lnk-expressing mouse ES and iPS cells were cultured in a leukemia inhibitory factor-containing ES cell medium (Millipore) on mitomycin C-treated mouse embryonic fibroblasts (MEFs) [28]. OP9 stromal cells were cultured in an α-minimum essential medium (α-MEM; Sigma) supplemented with 20% fetal bovine serum (FBS), 2 mM
In vitro hematopoietic differentiation
For embryoid body (EB) differentiation, mouse ES and iPS cells were trypsinized and collected in an EB medium (EBM) containing the Dulbecco's modified Eagle's medium (Wako) supplemented with 15% FBS, non-essential amino acids (Millipore), penicillin/streptomycin (Invitrogen), 2 mM
Flow cytometry
The following primary monoclonal antibodies (Abs), conjugated with fluorescein isothiocyanate, phycoerythrin, or allophycocyanin, were used for flow cytometric analysis: anti-CD45 (30-F11; eBioscience), anti-CD11b (M1/70; eBioscience), anti-Sca-1 (D7; eBioscience), anti-Ter119 (Ter-119; eBioscience), anti-CD34 (RAM34; eBioscience), anti-CXCR4 (2B11; BD Bioscience), anti-Gr-1 (RB6-8C5; eBioscience), anti-c-Kit (ACK2 or 2B8; eBioscience), and anti-CD41 (MWReg30; BD Bioscience). Purified rat anti-mouse c-Mpl/TPOR monoclonal Ab was obtained from IBL. Cells (1×105–5×105) were incubated with monoclonal Abs at 4°C for 30 min and washed twice with a staining buffer (phosphate-buffered saline/2% FBS). For detection of Mpl/TPOR, Dylight649-conjugated goat anti-rat IgG (BioLegend) was used as a secondary Ab. After staining, the hematopoietic cells were analyzed and isolated by flow cytometry on an LSR II and FACSAria flow cytometer, respectively, using FACSDiva software (BD Bioscience).
Colony assay and May-Giemsa staining
The cells (5×104 cells) were cultured in a Methocult M3434 medium containing IL-3, IL-6, SCF, and erythropoietin (StemCell Technologies, Inc.) for 10 days. The number of individual colonies was counted by microscopy. The colony number was normalized to the total number of hematopoietic cells. The multipotent hematopoietic progenitor cell-derived colonies (colony-forming unit–granulocyte, erythrocyte, monocyte, megakaryocyte (CFU-GEMM)/CFU-Mix) were picked up, fixed on glass slides using a cytospin centrifuge (Cytospin 4; Thermo Shandon), and stained with May-Grünwald Stain solution (Sigma) and Giemsa solution (Wako).
Western blotting
The adherent hematopoietic cells and OP9 stromal cells were collected, and were then incubated in a new tissue culture dish for 40 min to eliminate adherent OP9 cells. Floating cells were harvested and were subsequently starved in an RPMI1640 medium containing 0.1% FBS and penicillin/streptomycin for 4–6 h. Cells were stimulated with 20 ng/mL TPO for 10 min (for Jak2) or 30 min (for Erk and Akt) before being lyzed in a lysis buffer [20 mM Tris–HCl (pH 8.0), 137 mM NaCl, 1% Triton X-100, 10% glycerol] containing a protease inhibitor cocktail (Sigma) and a phosphatase inhibitor cocktail (Nacalai Tesque). Cell lysates were loaded onto polyacrylamide gels and were transferred to a polyvinylidene fluoride membrane (Millipore). After blocking, the membrane was exposed to mouse anti-phospho-Erk1/2 (Cell Signaling), rabbit anti-Erk1/2 (Sigma), mouse anti-phospho-Akt (Cell Signaling), rabbit anti-total Akt (Cell Signaling), rabbit anti-phospho-Jak2 (Tyr1007/1008; Cell Signaling), or rabbit anti-Jak2 (Cell Signaling), followed by horseradish peroxidase-conjugated secondary antibody. The band was visualized by ECL Plus Western blotting detection reagents (GE Healthcare) or Pierce Western Blotting Substrate Plus (Thermo Scientific), and the signals were read using an LAS-3000 imaging system (Fujifilm).
Reverse transcription–polymerase chain reaction
RT-PCR was carried out as described previously [25]. The sequences of the primers used in this study are listed in Table 1.
Results
Expression of Lnk in mouse ES and iPS cells
We initially investigated Lnk expression in mouse ES cells, iPS cells, ES cell-derived EBs (ES-EBs), and iPS-EBs. As shown in Fig. 1a, Lnk was expressed in undifferentiated ES and iPS cells, and the expression levels of Lnk were significantly increased after EB formation. We further examined whether Lnk was expressed in Flk-1-positive (+) cells, because hematopoietic cells were generated from Flk-1+ cells, a common hemoangiogenic progenitor during ES cell differentiation [29 –31]. Quantitative RT-PCR analysis after the purification of Flk-1+ cells from ES-EBs and iPS-EBs revealed that Lnk was highly expressed in Flk-1+ cells (Fig. 1b). These data suggest that Lnk plays some role in the hematopoietic differentiation process in ES and iPS cells.
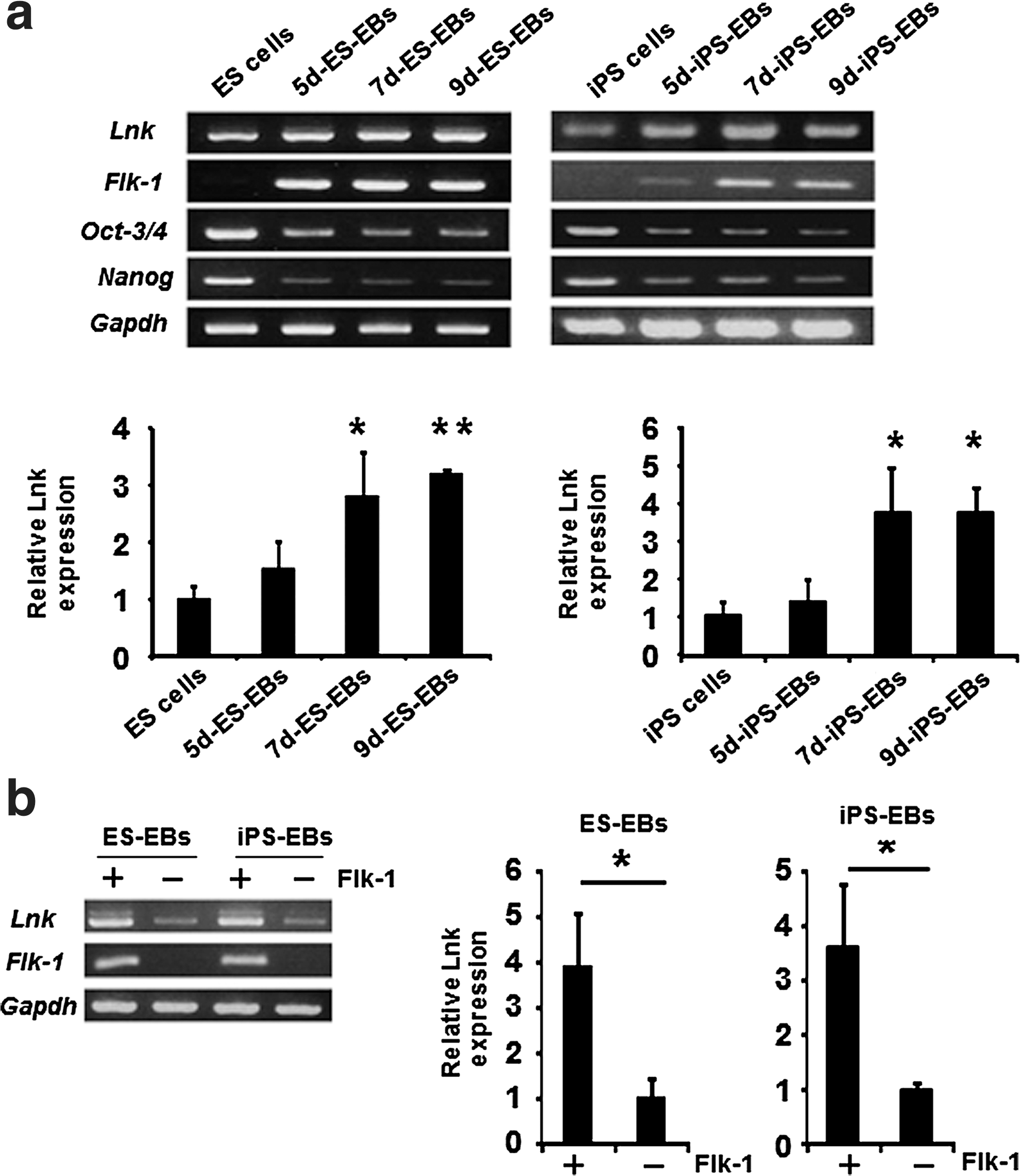
Lnk is expressed in mouse ES cells, iPS cells, and Flk-1+ hemoangiogenic progenitor cells.
Enhanced mesodermal differentiation in EB by the inhibition of Lnk
The data described above led to the expectation that hematopoietic cells, including hematopoietic progenitor cells, could be efficiently generated from ES cells and iPS cells by the suppression of Lnk. To inhibit the function of Lnk, we utilized the DN-Lnk gene, which was developed by Takizawa et al. [19]. DN-Lnk binds to Lnk, and forms a multimer complex by a homophilic interaction with the N-terminal domain, thereby inhibiting Lnk function [19]. DN-Lnk-expressing ES and iPS cells were generated by introducing a DN-Lnk-expressing plasmid, and DN-Lnk mRNA expression was confirmed by RT-PCR (Fig. 2a). In this report, we present data from one DN-Lnk-expressing ES and iPS cell clone, because the same results were obtained from other DN-Lnk-expressing clones. Notably, the expression levels of wild-type Lnk in DN-Lnk-expressing cells were similar to those in Neo-expressing cells and their parent cells (Fig. 2a). DN-Lnk-expressing iPS cells maintained the undifferentiated state in culture and possessed pluripotency, as demonstrated by alkaline phosphatase staining, immunostaining, and teratoma formation (Supplementary Fig. S1; Supplementary Data are available online at
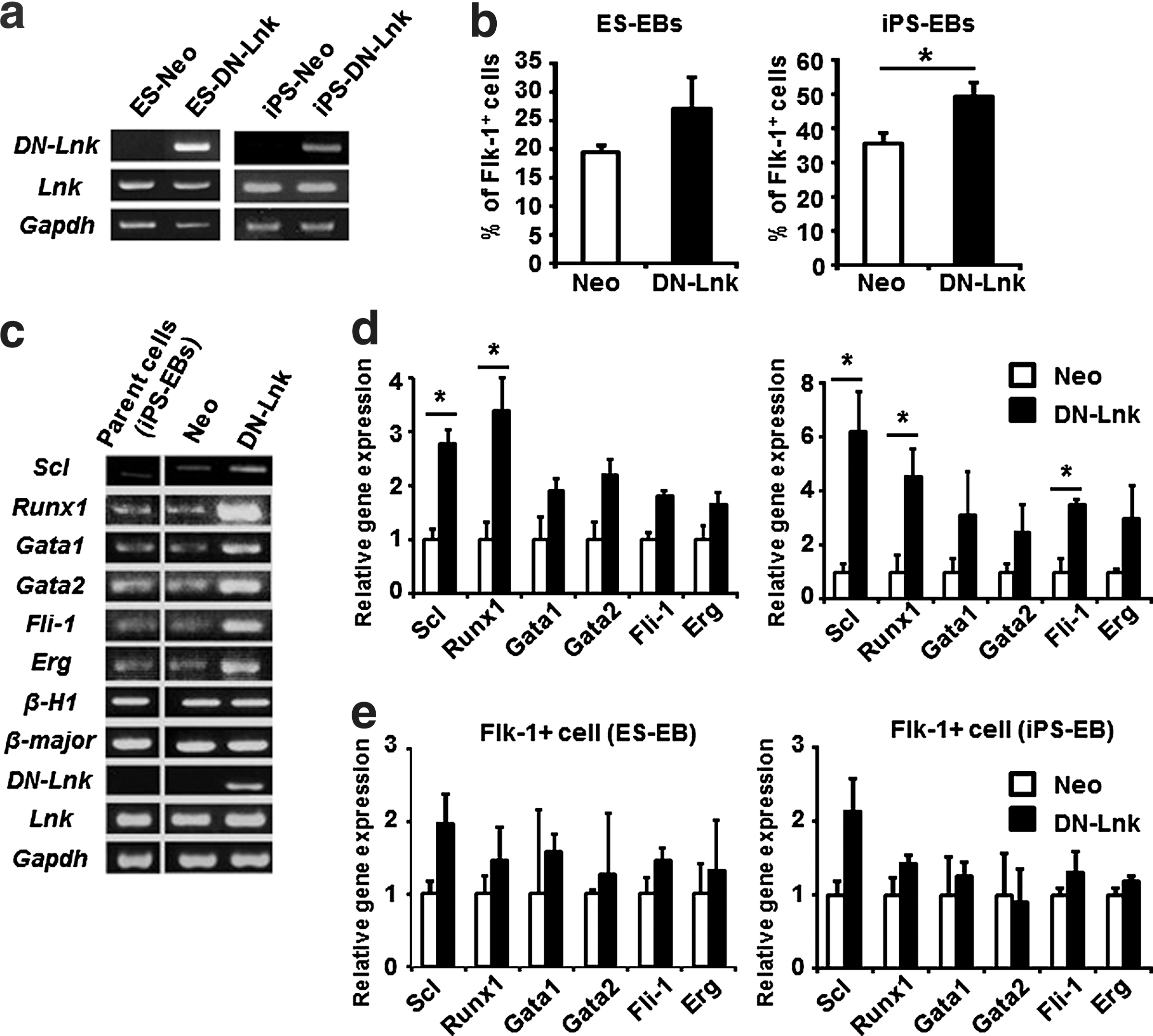
Increased expression of hematopoietic transcription factors in DN-Lnk-expressing EB cells.
Next, we generated EBs to induce mesodermal cells from DN-Lnk- or Neo-expressing ES and iPS cells. EBs were cultured for 7 days, because the proportion of Flk-1+ cells in EBs increased to a peak on day 7, and decreased over the next 2 days in our culture conditions (Supplementary Fig. S2). We found that DN-Lnk-expressing cells on day 7 of the EB culture yielded a modest increase in the number of Flk-1+ hemangiogenic progenitor cells relative to that of Neo-expressing cells (Fig. 2b). Interestingly, elevated expression of Scl/Tal-1, Runx1, and Gata-1 was observed in DN-Lnk-expressing total EB cells (Fig. 2c). Besides the expression levels of these genes, those of other key transcription factors of blood stem/progenitor cells, including Gata-2, Fli-1, and Erg [32], in DN-Lnk-expressing cells were also upregulated in comparison with those in Neo-expressing cells (Fig. 2d). To examine whether increased expression of these transcription factors in DN-Lnk-expressing cells was due to the increased generation of Flk-1+ cells, we performed the gene expression analysis after purification of Flk-1+ cells from DN-Lnk- or Neo-expressing total EB cells (Fig. 2e). No difference in the expression of Runx1, Gata-1, Gata-2, Fli-1, or Erg was observed between DN-Lnk-expressing cells and Neo-expressing cells, indicating that elevated expression of these hematopoietic genes in DN-Lnk-expressing EB cells would be largely because of the increased population of Flk-1+ cells. On the other hand, DN-Lnk-expressing Flk-1+ cells showed a 2-fold increase in the expression of Scl/Tal-1, an essential transcription factor for the hematopoietic development [33,34], compared with Neo-expressing Flk-1+ cells. The increased Scl/Tal-1 expression thus suggests that an inhibition of Lnk in Flk-1+ cells might contribute to enhance the production of hematopoietic progenitor cells. Taken together, these results raise the possibility that mesodermal cells with a hematopoietic differentiation potential would be efficiently generated in DN-Lnk-expressing cells during EB formation.
Inhibition of Lnk function increases the production of hematopoietic cells
To induce hematopoietic cells, EB-derived cells were cultured on OP9 stromal cells in the presence of hematopoietic cytokines. During culture, cobblestone-forming cells were more frequently observed in DN-Lnk-expressing cells than in Neo-expressing cells (Fig. 3a), indicating that DN-Lnk-expressing cells were immature hematopoietic cells with expansion potential. In support of this observation, DN-Lnk-expressing cells showed a significant increase in the number of hematopoietic cells compared to that of Neo-expressing cells (Fig. 3b). Importantly, compared to Neo-expressing cells, DN-Lnk-expressing cells could more efficiently proliferate on OP9 stromal cells for a period exceeding 14 days (Fig. 3b). Therefore, the proliferation of hematopoietic cells could be augmented by the inhibition of Lnk.

Efficient generation of hematopoietic cells from iPS cells by overexpression of the DN-Lnk gene. EBs derived from Neo- or DN-Lnk-expressing iPS cells were cultured for 7 days, and were then plated and cultured on OP9 cells with hematopoietic cytokines to induce and expand the hematopoietic cells.
To investigate whether primitive and definitive hematopoiesis could occur in DN-Lnk- or Neo-expressing cells, we measured the expression levels of red cell globin by RT-PCR analysis. In both DN-Lnk- and Neo-expressing hematopoietic cells, the expression levels of the embryonic globin, β-H1, and the adult globin, β-major, were decreased and increased, respectively, after culturing on OP9 stromal cells in comparison with those in total EB cells (Fig. 3c). This indicates that DN-Lnk- or Neo-expressing cells can show the primitive hematopoiesis followed by definitive hematopoiesis under our culture conditions.
We next examined the colony-forming potential of DN-Lnk-expressing cells. As shown in Fig. 3d, DN-Lnk-expressing cells showed a significant increase in the total colony-forming cell (CFC) number and CFU–granulocyte, macrophage number. Note that the number of CFU-GEMM/CFU-Mix, the most immature multipotent hematopoietic cells, in DN-Lnk-expressing cells was ∼5 times as much as that in Neo-expressing cells (Fig. 3d). May-Giemsa staining after picking up the colonies revealed that mixed colonies derived from DN-Lnk-expressing cells contained the erythroblasts, granulocytes, macrophages, and megakaryocytes (Fig. 3e), thus confirming the generation of multipotent hematopoietic cells. An elevated CFU-Mix number in DN-Lnk-expressing cells might have been due to the fact that Lnk is highly expressed in immature hematopoietic cells, especially in hematopoietic stem/progenitor cells [13,17]. We also analyzed surface antigen expression in DN-Lnk- or Neo-expressing cells by flow cytometry, and found that DN-Lnk-expressing cells showed a higher percentage of CD34+ cells and CD41+ cells (Fig. 3f), suggestive of an increased number of immature hematopoietic cells. In addition, the proportion of CD45+ cells, CD11b+ cells, Gr-1+ cells, or CXCR4+ cells was also increased in DN-Lnk-expressing cells (Fig. 3f). By contrast, a lower percentage of Ter119+ cells were observed in DN-Lnk-expressing cells (Fig. 3f). Consistent with this flow cytometric analysis, we found an increased expression of Sfpi1 (encoding Pu.1) and E47, which are the key factors responsible for hematopoiesis, and a decreased expression of β-major globin in DN-Lnk-expressing cells after the cultivation on OP9 stromal cells (Fig. 3c and Supplementary Fig. S3). These results clearly showed that Lnk inhibition promoted the production of hematopoietic cells, including multipotent immature hematopoietic cells and myeloid cells, from mouse ES and iPS cells.
Inhibition of Lnk function in pluripotent stem cell-derived hematopoietic cells augments TPO-mediated signaling
It was previously shown that Lnk negatively regulates various types of hematopoietic cytokine signaling, such as TPO [16]. To investigate whether the increased production of hematopoietic cells from DN-Lnk-expressing cells, described above, is due to the enhanced TPO-mediated signaling, we analyzed protein phosphorylation after TPO stimulation using DN-Lnk-expressing cells. Hematopoietic cells were starved and subsequently stimulated with 20 ng/mL of TPO before the preparation of the cell lysates. The results showed the elevated phosphorylation of Jak2, Erk1/2, and Akt, all of which are downstream of TPO signaling, in DN-Lnk-expressing cells (Fig. 4a). We also found almost no difference in the percentage of Mpl/TPOR-positive cells between DN-Lnk-expressing cells and Neo-expressing cells (Fig. 4b), indicating that enhanced TPO signaling in DN-Lnk-expressing cells does not result from the increased percentage of Mpl/TPO-expressing cells. Thus, our data suggest that Lnk inhibition by DN-Lnk gene transduction would augment the activation of signaling molecules upon stimulation with cytokines, and thus Lnk inhibition would promote the production of hematopoietic cells in DN-Lnk-expressing cells.
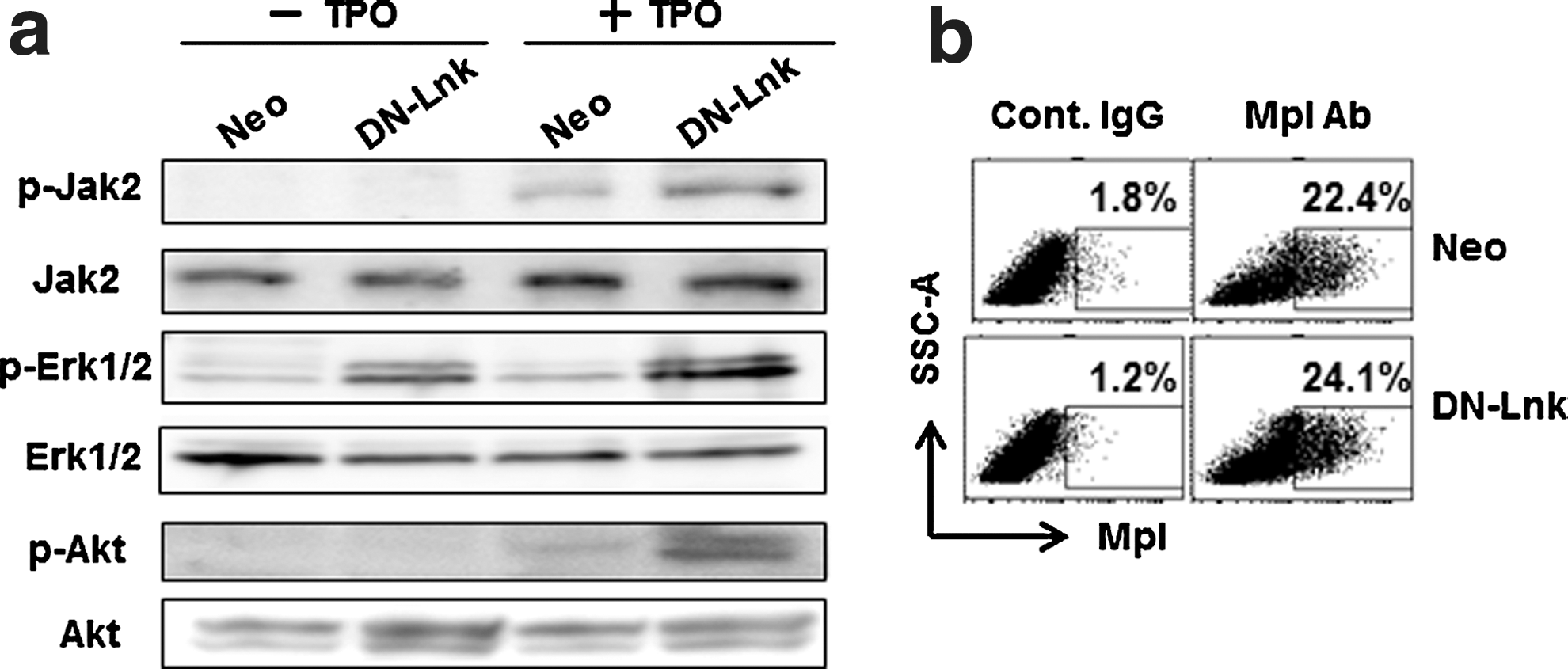
Enhanced TPO-induced signaling pathway in DN-Lnk-expressing cells. After the EB cells, which were cultured for 7 days, had been plated and cultured on OP9 stromal cells for 14 days, and the hematopoietic cells were harvested as described in the Materials and Methods section.
Increased generation of hematopoietic progenitor cells from mouse pluripotent stem cells by transient transduction of a DN-Lnk gene
Our groups have shown that Ad vector-mediated transient, but not constitutive, transduction of differentiation-related genes in pluripotent stem cells could result in the efficient generation of functional cells, such as adipocytes, osteoblasts, hepatocytes, and hematopoietic cells [25,28,35 –37]. We expected that the transient inhibition of Lnk in iPS cells could also accelerate the hematopoietic differentiation. To test this expectation, we generated a DN-Lnk-expressing Ad vector, Ad-DN-Lnk, and examined the effects of transient Lnk inhibition on hematopoietic cell differentiation. The transduction efficiency in EBs, which was transduced with a DsRed-expressing Ad vector, was approximately 40%, as determined by flow cytometry (data not shown). A colony assay after transduction with Ad vectors revealed that the number of total colonies and mixed colonies in the cells transduced with Ad-DN-Lnk was slightly increased in comparison with that in the cells transduced with Ad-LacZ (control vector) (Fig. 5a, c). Moreover, the number of hematopoietic cells increased in Ad-DN-Lnk-transduced cells after 7-day cultivation on OP9 stromal cells (Fig. 5b, d). Thus, our data indicate that the transient inhibition of Lnk also enhances the differentiation and proliferation of hematopoietic cells derived from pluripotent stem cells.
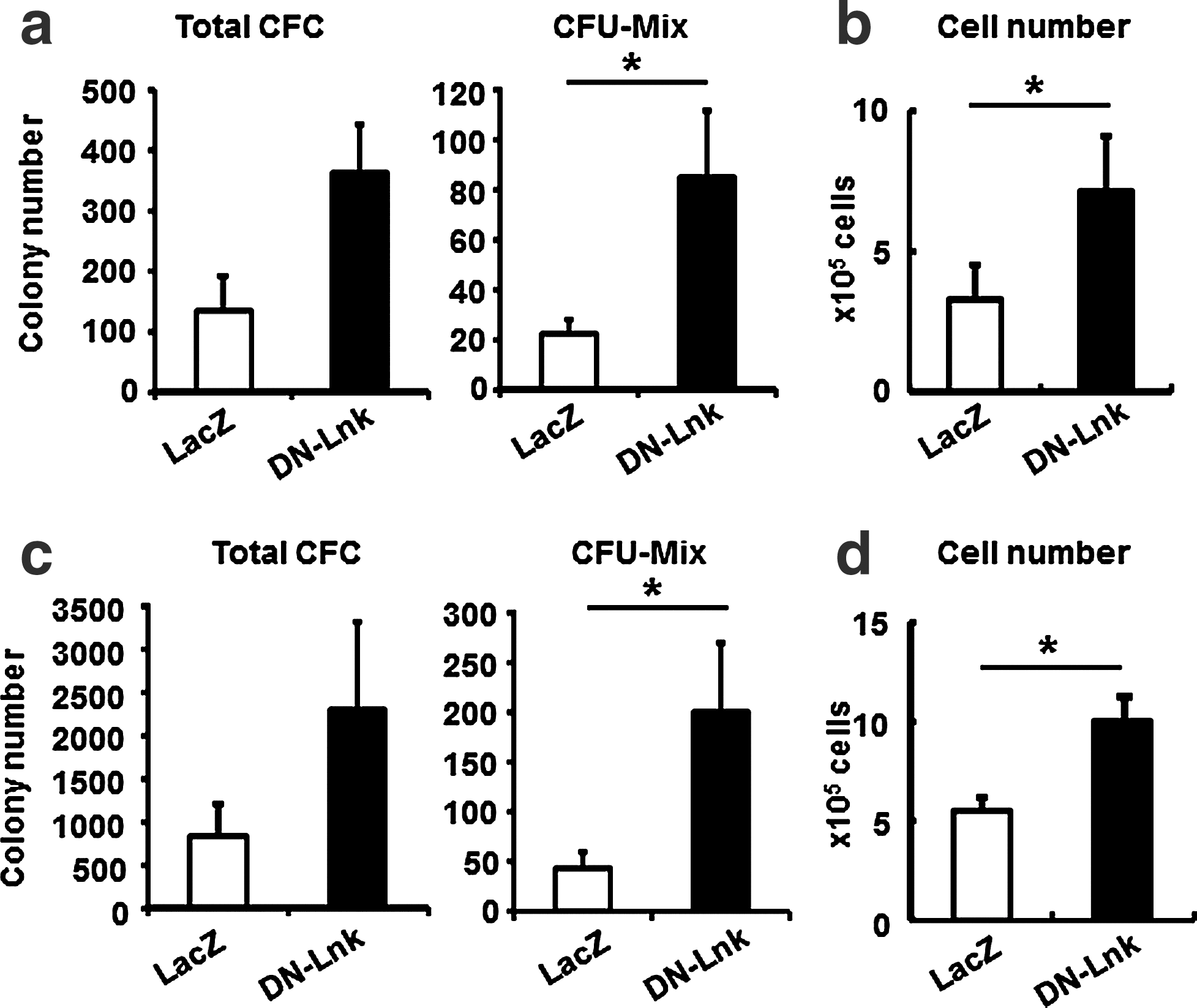
Expansion of ES cell- and iPS cell-derived hematopoietic cells by the transient expression of DN-Lnk in EB cells. After ES cell-
Discussion
In this report, we successfully generated and expanded hematopoietic cells, including immature hematopoietic cells, with colony-forming potential, from mouse ES and iPS cells by the suppression of an adaptor protein Lnk (Fig. 3). We also demonstrated that the expression levels of hematopoietic transcription factors such as Scl and Runx1 in DN-Lnk-expressing total EB cells were significantly increased in comparison with those in Neo-expressing total EB cells (Fig. 2c, d), and that cytokine response was augmented in DN-Lnk-expressing cells (Fig. 4). Therefore, the data obtained in this study suggest that Lnk inhibition by enforced expression of a DN-Lnk gene in ES and iPS cells would lead both to a promotion of mesodermal differentiation during EB formation and to an increase in the expansion potential of ES and iPS cell-derived hematopoietic cells on an OP9 coculture system, and thus Lnk inhibition could enhance the hematopoietic cell production.
In developing mouse embryos, Lnk is shown to be expressed in the aorta-gonad-mesonephros (AGM) region, the site of hematopoiesis [38]. It has also been reported that the production of CD45+ hematopoietic cells was severely impaired by the enforced expression of Lnk in AGM-derived cells, suggesting that Lnk suppresses hematopoietic commitment [38]. However, the function of Lnk in hematopoiesis is not fully understood. In the current study, we found that Lnk was highly expressed in Flk-1+ cells (Fig. 1b), which are known to be hemoangiogenic progenitor cells during ES cell differentiation [29]. Furthermore, it was of note that levels of expression of Scl/Tal-1, which is essential for hematopoietic commitment of hemoangiogenic progenitor cells derived from ES cells [34], were slightly upregulated in Flk-1+ cells by the inhibition of Lnk function (Fig. 2e). Thus, it is possible that Lnk might negatively regulate the hematopoietic commitment in Flk-1+ cells by modulating the expression of Scl/Tal-1. We also showed that the percentage of Flk-1+ cells was increased in DN-Lnk-expressing EB cells (Fig. 2b), and this could result in the elevated expression of other key hematopoietic transcription factors, such as Runx1 and Gata-1, in DN-Lnk-expressing total EB cells compared with that in Neo-expressing total EB cells (Fig. 2c, d). This indicates that the functional Flk-1+ mesodermal cells would be efficiently generated from DN-Lnk-expressing ES and iPS cells. On the other hand, at earlier days of differentiation, the percentage of CD41+ cells, an early hematopoietic progenitor cells generated from pluripotent stem cells [39], in DN-Lnk-expressing EB cells was mostly equal to that in Neo-expressing EB cells (Supplementary Fig. S4). Taken together, the findings suggest that Lnk inhibition in ES and iPS cells could be effective for the generation of mesodermal cells with the potential for hematopoietic differentiation, but would not enhance the emergence of hematopoietic progenitor cells at earlier days of EB differentiation.
We examined the cytokine responses of iPS cell-derived hematopoietic cells, and observed the augmented phosphorylation of Erk and Akt in DN-Lnk-expressing cells (Fig. 4). This result is consistent with that of a previous report in which TPO-treated megakaryocytes derived from Lnk-deficient mice enhanced the extent of the activation of Erk and Akt [40]. By contrast, it was reported that Lnk-deficient adult HSCs or bone marrow-derived macrophages showed an enhanced Akt, but not Erk, activation after cytokine stimulation [16,41]. This difference in the activation of downstream molecules is most likely due to differences in cell populations. Because ES cell- and iPS cell-derived hematopoietic cells are heterogeneous, both Akt and Erk phosphorylation levels after cytokine treatment would be augmented in DN-Lnk-expressing cells relative to Neo-expressing cells. In general, Akt and Erk are known to be involved in cell survival and cell growth [42,43]. Hence, our data indicate that the suppression of Lnk by the ectopic expression of DN-Lnk in ES and iPS cells would lead to an increase in hematopoietic cell production through enhanced cytokine responses.
Recently, Dravid et al. reported the expression of Lnk in human ES cell-derived CD34+ hematopoietic progenitor cells, and they showed that the number of human ES cell-derived CD34+ cells was increased by Lnk knockdown using a lentivirus vector carrying the short-hairpin RNA against Lnk (shLnk) [44]. Their results are mostly consistent with our data, indicating the suppressive function of Lnk in hematopoietic cell generation in both mouse and human pluripotent stem cells. However, the generation of hematopoietic CFCs in shLnk-transduced cells and the molecular mechanisms associated with the generation of CD34+ cells by Lnk knockdown have not been addressed in detail. In this report, we clearly demonstrated that hematopoietic CFCs, including immature multipotent hematopoietic cells, were efficiently generated from mouse ES and iPS cells by Lnk inhibition, and these cells show potential for expansion on OP9 stromal cells (Fig. 3). In addition, we showed that the enhanced generation of hematopoietic cells in DN-Lnk-expressing cells was mediated by the promotion of mesodermal differentiation in EBs and augmented the sensitivity to cytokines in DN-Lnk-expressing cells as described above.
Another important finding of this study was that the transient inhibition of Lnk by Ad vector-mediated transduction of a DN-Lnk gene could also be an effective strategy for expanding hematopoietic cells (Fig. 5). Recently, the loss of Lnk and the mutation of Lnk have been reported to be associated with myeloproliferative diseases [45,46], indicating that oncogenesis may result from constitutive Lnk suppression in ES cell- and iPS cell-derived hematopoietic cells via the overexpression of a DN-Lnk gene or a lentivirus vector-mediated knockdown system, and such suppression would not be a directly applicable approach for clinical medicine. In this regard, our approaches using Ad vector-mediated transient Lnk inhibition are thought to be quite useful for the safe expansion of the ES cell- and iPS cell-derived hematopoietic cells. However, the number of ES cell- and iPS cell-derived hematopoietic cells in Ad-DN-Lnk-transduced cells was lower than that in the case of stably DN-Lnk-expressing cells (data not shown), possibly due to the low transduction efficiency of Ad vectors in EB cells. Therefore, it will still be necessary to establish methods for transiently inhibiting Lnk functionality using various types of Ad vectors [47] and short interference RNA.
In summary, we successfully developed efficient methods for differentiating mouse ES and iPS cells into hematopoietic cells by the suppression of the adaptor protein Lnk. Lnk functions downstream of multiple hematopoietic cytokine-signaling events, including those involving TPO, SCF, macrophage-colony stimulating factor, and erythropoietin [13,16,40,41,48,49], and Lnk-deficient mice show accumulation of pro-B cells in the bone marrow [48]. Therefore, various types of hematopoietic cells may be efficiently differentiated and expanded from ES and iPS cells by the inhibition of the Lnk function, when an appropriate cytokine is included in the culture; such work is currently ongoing in our laboratory.
Footnotes
Acknowledgments
We thank Misae Nishijima (National Institute of Biomedical Innovation) for her help. We thank Dr. Kazufumi Katayama (Osaka University, Osaka, Japan) for helpful discussion. We also thank Dr. S. Yamanaka and Dr. J. Miyazaki for kindly providing the mouse iPS cell line 38C2 and the CA promoter, respectively. This work was supported by a Grant-in-Aid for Young Scientists (B) (K. Tashiro) of the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) of Japan and the Ministry of Health, Labour, and Welfare of Japan.
Author Disclosure Statement
The authors have no financial conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.