
Abstract
Adult stem cells reside in many types of tissues and adult stem cell-based regenerative medicine holds great promise for repair of diseased tissues. Recently, adipose-derived stem cells (ADSCs) were found to be an appealing alternative to bone marrow stem cells (BMSCs) for tissue-engineered bone regeneration. Compared with BMSCs, ADSCs can be easily and abundantly available from adipose tissue. However, our previous study has discovered an important phenomenon that BMSCs have greater osteogenic potential than ADSCs in vitro. In this study, we aimed to explore its mechanism and improve the osteogenic potential of ADSCs for bone tissue regeneration. It has been reported that the epigenetic states could contribute to lineage-specific differentiation of adult stem cells. We observed that the epigenetic changes of BMSCs were much greater compared with ADSCs after a 3-day osteogenic induction. Runt-related transcription factor 2 (Runx2) is essential for osteoblast differentiation and bone formation. We found that BMSCs underwent more obvious epigenetic changes on the Runx2 promoter than ADSCs after osteogenic induction. These results suggest the epigenetic regulation involvement in Runx2 expression, and thus osteogenesis. We subsequently used a histone deacetylase inhibitor, trichostatin A (TSA), to promote the osteogenesis capacity of ADSCs. The results showed that TSA promoted rat ADSCs osteogenic differentiation by altering the epigenetic modifications on the Runx2 promoter in a bone morphogenetic protein signaling-dependent manner.
Introduction
B
Bone marrow stem cells (BMSCs) have been considered an ideal cell source for bone tissue engineering, due to their natural osteoprogenitor characteristics. However, some problems have limited the expansion of BMSC-based clinical application, such as the painful bone marrow biopsy procedure, its limited number of harvested stem cells, and aging rapidly in culture [2]. This has raised concerns over the effect and safety of BMSC-based therapies. Adipose-derived stem cells (ADSCs) can be easily and abundantly available from the adipose tissue and capable of differentiating into multiple mesenchymal cell types, such as adipocytes, chondrocytes, osteoblasts, and myoblasts [3 –6]. They can overcome some of the aforementioned BMSC disadvantages and reduce or even eliminate the need for costly and lengthy tissue culture expansion. Therefore, ADSCs may be a promising candidate cell source for tissue engineering.
However, in our previous and other studies, it has been shown that in vitro, the capacity for osteogenic differentiation of BMSCs was better compared with ADSCs [7,8]. Hence, in this study, we would like to elucidate the underlying mechanism for their different osteogenesis capacity and try to promote the osteogenesis capacity of ADSCs for further clinical applications.
Recently, epigenetic regulation has been considered as an important mechanism to influence stem cell differentiation. The unique patterns of DNA methylation and histone modifications have been found to play important roles in the induction of lineage-specific differentiation of MSCs [9]. However, whether there are differences in epigenetic modifications between ADSCs and BMSCs have not been studied before.
Osteogenic differentiation of MSCs is a complex process, which is regulated by a number of transcription factors. Runt-related transcription factor 2 (Runx2) is essential for osteoblast differentiation and bone formation. Humans with heterozygous mutations or deletions of Runx2 developed cleidocranial dysplasia [10,11]. Overexpression of Runx2 can induce and upregulate the expression of multiple osteoblast specific genes in nonosteogenic cells [12,13]. Furthermore, studies have reported that several histone deacetylase (HDAC) inhibitors promote osteoblast maturation and the expression of osteoblast-specific genes through upregulation of Runx2 activity in MC3T3-E1 preosteoblast cells [14]. Meanwhile, our previous studies have shown that BMSCs had a significantly higher expression of Runx2 during osteogenesis than ADSCs [7]. However, whether the epigenetic modifications of Runx2 promoter are different between ADSCs and BMSCs and responsible for their osteogenesis capacity have not been studied before.
Thus, in this study, we aim to compare the epigenetic modifications of Runx2 promoter between ADSCs and BMSCs and try to enhance the ADSCs osteogenesis capacity through altering their epigenetic states using a HDAC inhibitor trichostatin A (TSA).
Materials and Methods
Cell culture
Sprague-Dawley rat BMSCs and ADSCs were purchased from Cyagen Biosciences (Guangzhou, China). Cells were maintained in a complete Dulbecco's Modified Eagle's Medium (DMEM) and the osteogenic differentiation was induced in the osteogenic differentiation medium (Cyagen Biosciences). The medium was changed every 1 to 2 days.
Alizarin red and alkaline phosphatase staining
The cells in flask-style chambers on glass slides were induced in the osteogenic differentiation medium for 0, 3, 7, and 14 days. Then the cells were stained with the alizarin red solution (Cyagen Biosciences) according to the manufacturer's instruction to determine the presence of calcium deposition, an indicator of the osteogenic lineage. This is an early stage marker of matrix mineralization, a crucial step toward the formation of calcified extracellular matrix associated with true bone.
The cells were also stained with an alkaline phosphatase (ALP) staining solution (Sigma Diagnostics, Inc., St. Louis, MO) after 7 days induction to examine the osteogenic differentiation.
Western blot analysis
Western blot analysis was performed as previously described [15]. Antibodies against acetyl-histone H3K9, trimethyl-histone H3K9, trimethyl-histone H3K4, and histone H3 were purchased from Upstate Biotechnology (Lake Placid, NY). Bound primary antibodies were detected with immunoglobulins conjugated with horseradish peroxidase (HRP; Upstate) and visualized by enhanced chemiluminescence (ECL). ECL signals were quantified using Image J software and normalized against the H3 intensity level.
Enzyme-linked immunoabsorbent assay
The in vitro ALP production was quantified using a commercial ALP enzyme-linked immunoabsorbent assay (ELISA) kit (Hermes Criterion Biotechnology, Vancouver, BC, Canada) according to the manufacturer's instructions. Briefly, the cells seeded in a 6-well plate at 1×106 cells per well (3 wells for each time point) were induced in the osteogenic differentiation medium. On days 0, 3, 7, and 14, the cells were harvested and dissociated in 0.1 M Tris (pH 7.4) containing 1% Triton X-100 and 5 mM MgCl2 by sonication. The ALP concentration was measured and the data were normalized against the protein concentration and expressed as nanogram of ALP per milligram of total protein. The protein concentration was measured with a BCA protein assay kit (Pierce Biotechnology, Inc., Rockford, IL) using bovine serum albumin as a standard.
Flow cytometry analysis for cell apoptosis
Cells were cultured in a 6-well plate at 1×106 cells per well with complete DMEM in the presence or absence of a HDAC inhibitor TSA (Sigma) that was dissolved in dimethylsulfoxide at indicated concentrations. After 72 h of culture, cells were harvested and washed 3 times in phosphate-buffered saline. Cells from each sample were processed for Annexin V fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis detection (Becton Dickinson, Franklin Lakes, NJ) according to the manufacturer's instructions. All experiments were carried out in triplicates and repeated 3 times.
Cytotoxicity assays
Cells were cultured in a 96-well plate at 1×103 cells per well with 100 μL complete DMEM in the presence or absence of a HDAC inhibitor TSA for 72 h. Cytotoxicity was measured by performing WST-8 assay using a CCK-8 cell proliferation kit (Dojindo Laboratories, Kamimashiki Gun, Kumamoto, Japan), according to the manufacturer's instructions. The 450 nm absorbance was measured with a microplate reader (Bio-Rad, La Jolla, CA). All experiments were carried out in triplicate and 3 independent experiments were performed.
Real-time reverse transcription-polymerase chain reaction analysis
Cells were seeded into a 6-well plate at 1×105 cells per well and harvested at assigned time points postinduction. Total RNA extraction was performed using the Trizol Reagent (Invitrogen, Carlsbad, CA) based on the manufacturer's instructions. Amplifications were performed with the ABI 7300 Real-Time PCR System (Carlsbad, CA) with different primers. The primers used were as follows: Runx2 forward TCCAGACCAGCAGCACTCC and reverse TCAGC GTCAACACCATCATTC; osteopontin (OPN) forward AAT GAAGGGCCCTGAGC and reverse GCCAGTTCTGCAAGG AAGC; osteocalcin (OCN) forward AACGGTGGTGCCAT AGATGC and reverse AGGACCCTCTCTCTGCTCAC; BMP2 forward GGAAAACTTCCCGACGCTTCT and reverse CCTGCATTTGTTCCCGAAAA; BMP4 forward TTATGAGG TTATGAAGCCCCCA and GCTCACATCGAAAGTTTCC CAC; 18S rRNA forward GTAACCCGTTGAACCCCATT and reverse CCATCCAATCGGTAGTAGCG. All annealing temperatures were set at 60°C. Transcription levels were normalized to 18S rRNA level. Each value represents the average of at least 3 independent experiments.
Chromatin immunoprecipitation assay
Formaldehyde cross-linked chromatin preparation and chromatin immunoprecipitation (ChIP) were carried out using the ChIP assay kit (Upstate). Briefly, chromatin was cross linked with 1% formaldehyde for 10 min at room temperature. Cells were washed twice in ice-cold PBS, resuspended in the SDS-lysis buffer, and sonicated until the cross-linked chromatin was sheared to an average DNA fragment length of 100–800 bp. Normal rabbit IgG was used as a negative control, and antibodies against acetyl-histone H3K9, trimethyl-histone H3K9, and trimethyl-histone H3K4 were used for each immunoprecipitation. The immunoprecipitated DNAs were then analyzed using real-time polymerase chain reaction (PCR). For the rat Runx2 promoter region, the forward primer was 5′-GGACCGCCTCCT TCCAACT-3′, while the reverse primer was 5′-TCACTCGC CTCCGTCTACC-3′. The annealing temperature was 60°C. The amount of immunoprecipitated DNA was normalized to the input DNA. Each value represents the average of at least 3 independent experiments.
Statistical analysis
The results are expressed as means±standard deviation. Comparisons between groups were analyzed using the Student's t-test or ANOVA, and the Student-Newman-Kleuss method was used to estimate the level of significance. Differences were considered to be statistically significant for P<0.05.
Results
BMSCs possess higher osteogenic differentiation capacity than ADSCs
We first investigated the osteogenic differentiation potential of ADSCs and BMSCs. ALP and alizarin red staining showed that these 2 types of MSCs all had an osteogenic differentiation capacity after induction (Fig. 1A), but BMSCs showed much higher increases in ALP and alizarin red staining compared with ADSCs. The real-time RT-PCR results showed that the expression of the osteogenesis specific gene Runx2 was peaked at day 3 in BMSCs, but the maximum expression of Runx2 was seen at day 7 in ADSCs (Fig. 1B). We also detected the expression of 2 osteogenic-related genes, OCN and OPN. As shown in Fig. 1C and D, the expression of OCN and OPN were upregulated after osteogenic induction. The expression of OCN peaked at day 7 in BMSCs and at day 14 in ADSCs (Fig. 1C), while the expression of OPN (Fig. 1D) and the ALP concentration were much higher in BMSCs than ADSCs (Fig. 1E) throughout the induction process. These results demonstrate that BMSCs possess a higher osteogenic differentiation capacity than ADSCs.

Comparative analysis of the osteogenesis capacity of ADSCs and BMSCs.
The epigenetic changes on the Runx2 promoter are more obvious in BMSCs than in ADSCs
Epigenetic modulation plays a critical role in stem cells differentiations [16,17]. To determine whether epigenetic mechanisms were involved in ADSCs and BMSCs osteogenesis, we examined the epigenetic characteristics of ADSCs and BMSCs during their osteogenic differentiation process. As shown in Fig. 2A, the degree of transcriptionally permissive histone modifications of H3K9 acetylation and H3K4 trimethylation in both ADSCs and BMSCs were increased after osteogenic induction, while the level of transcriptionally repressive histone modification of H3K9 trimethylation was simultaneously decreased. Moreover, the epigenetic changes in BMSCs were more obvious than in ADSCs.
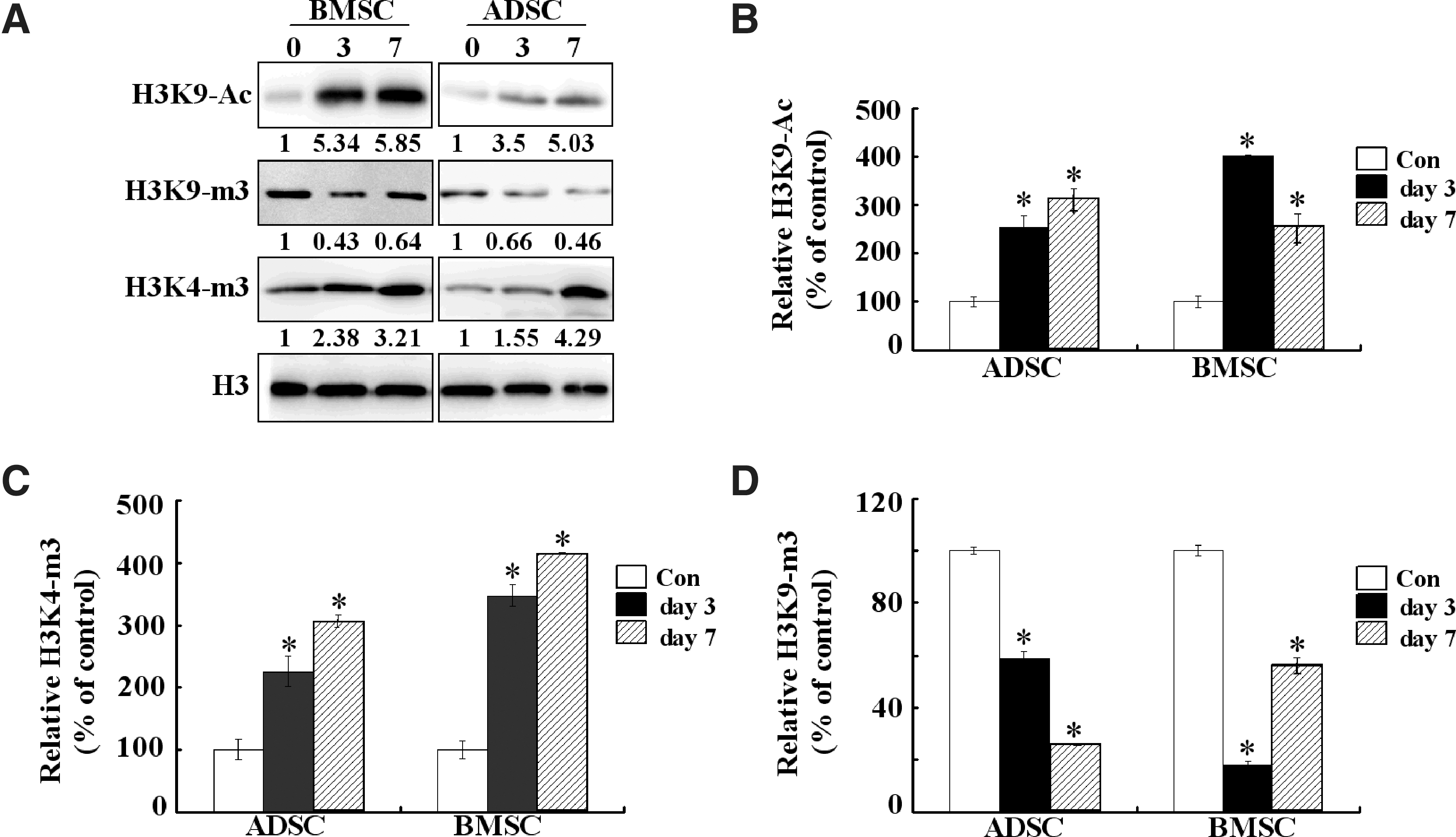
Epigenetic modifications on histones during osteogenesis of ADSCs and BMSCs.
Runx2 is essential for osteoblast differentiation and bone formation. To determine whether epigenetic changes of histone H3 occurred on the Runx2 promoter during osteogenesis, we performed ChIP assays. As shown in Fig. 2B and C, the recruitments of acetylated H3K9 and trimethylated H3K4 on the Runx2 promoter were increased after 3 days osteogenic induction both in ADSCs and BMSCs. In contrast, the recruitment of trimethylated H3K9 was decreased (Fig. 2D). Additionally, in comparison with ADSCs, BMSCs underwent more obvious epigenetic changes on the Runx2 promoter after 3 days osteogenic induction. After 7 days osteogenic induction, the recruitment of trimethylated H3K4 on the Runx2 promoter was also increased in both ADSCs and BMSCs (Fig. 2C). However, the recruitment of acetylated H3K9 and trimethylated H3K9 on the Runx2 promoter was different between ADSCs and BMSCs. The recruitment of acetylated H3K9 was also increased in ADSCs at day 7, while it was downregulated in BMSCs. Meanwhile, the recruitment of trimethylated H3K9 was also decreased in ADSCs at day 7, but it was upregulated in BMSCs. These epigenetic modification changes on the Runx2 promoter were in accordance with the expression profile of Runx2 in Fig. 1B, that is, the expression of Runx2 was further upregulated in ADSCs, but downregulated in BMSCs.
TSA enhances the osteogenic differentiation capacity of ADSCs
Our results indicate that the osteogenic differentiation capacity of BMSCs seems higher compared with ADSCs. We also observed epigenetic modification differences between BMSCs and ADSCs. To determine whether epigenetic changes could affect osteogenic differentiation of ADSCs, we treated the cells with a potent HDAC inhibitor, TSA. We first examined TSA's effect on ADSCs' survival. The cells were treated with TSA (0–500 nM) for 72 h. Then, we performed flow cytometry to quantify the apoptosis rate of ADSCs with Annexin V FITC/PI staining. As shown in Fig. 3A, TSA had no effect on the percentage of Annexin V-positive ADSCs at concentrations of 20, 75, and 150 nM. However, the percentage of Annexin V-positive cells increased along with 300 and 500 nM TSA treatment, indicating an enhanced apoptosis. The cytotoxicity of TSA was determined by CCK-8 analysis. As shown in Fig. 3B, ADSCs viability was decreased in a dose-dependent manner, while 20–75 nM TSA induced minimal cytotoxicity to ADSCs.
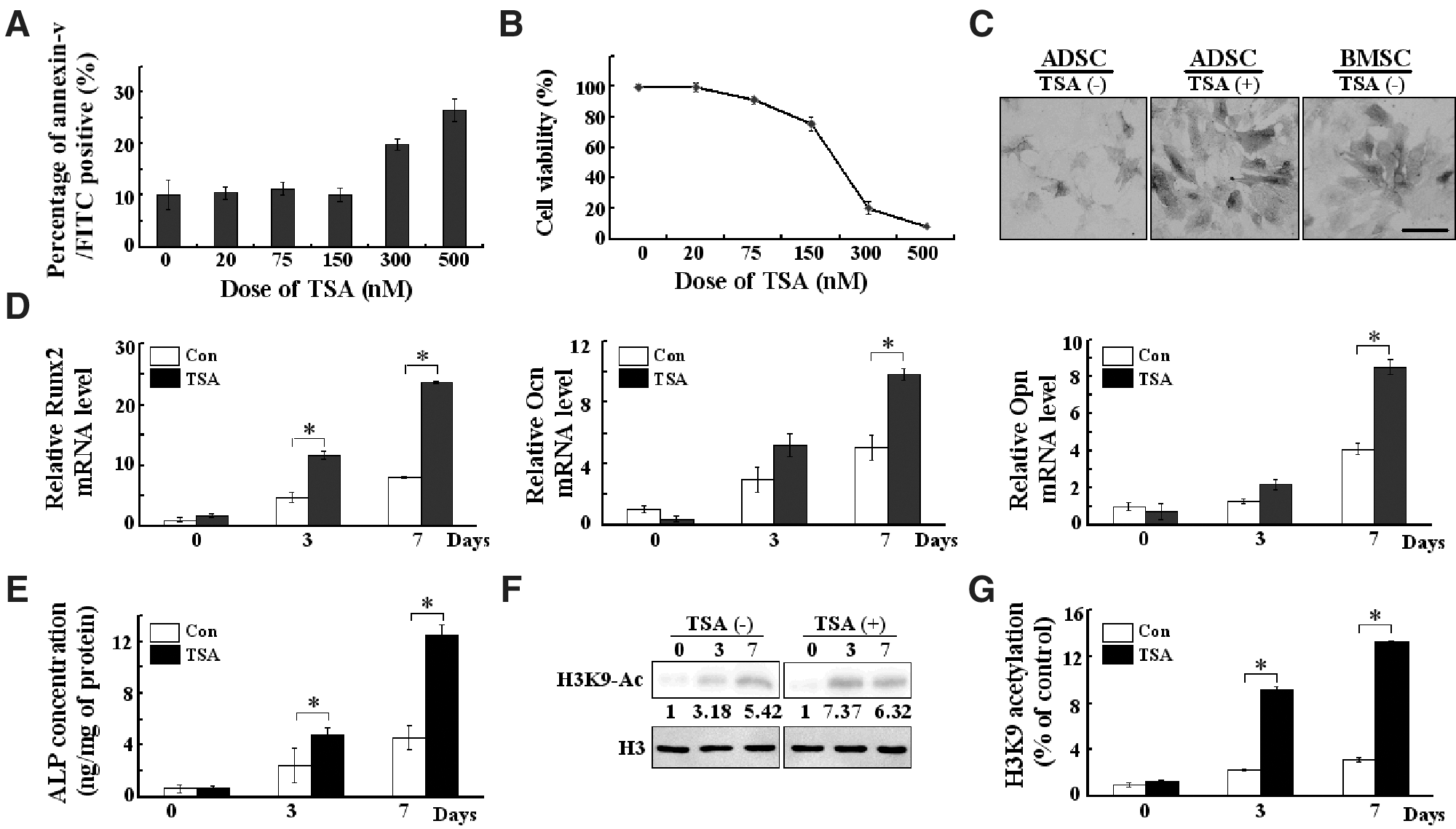
The influence of histone deacetylase inhibitor on the osteogenic differentiation of ADSCs.
To examine the osteogenic effect of TSA, ADSCs were treated with 75 nM TSA during the first 3 days of differentiation. As shown in Fig. 3C and E, TSA could significantly enhance the ALP staining and ALP production of ADSCs, even stronger compared with nonstimulated BMSCs, suggesting that the treatment of TSA significantly increased the osteogenic differentiation capacity of ADSCs. Furthermore, we found that histone H3K9 acetylation (Fig. 3F) as well as the expression of Runx2, OCN, and OPN (Fig. 3D) increased after TSA treatment. This indicates that the suppression of HDAC enzymatic activity enhanced osteogenic differentiation concomitantly with the stimulation of osteogenesis-specific genes expression. Meanwhile, ChIP assay demonstrated that TSA significantly increased acetylated histone H3K9 recruitment onto the Runx2 promoter (Fig. 3G). These results indicate that the upregulation of Runx2 during osteogenesis is tightly associated with the selective induction of histone hyperacetylation at its promoter regions.
BMP signaling is required for TSA-induced osteogenic differentiation of ADSCs
BMPs are potent osteogenic agents that have strong abilities to induce ectopic bone formation [18]. It has been proposed that Runx2 is the principal downstream target of BMPs, and hence that BMP activity is required for induction of osteoblast differentiation mediated by Runx2 [19]. This knowledge led us to exam whether BMP signaling pathway is involved in TSAs osteogenic effects. Indeed, 75 nM TSA could enhance the expression of BMP2 and BMP4 in ADSCs after osteogenic induction (Fig. 4A, B). Considering that the expression of Runx2 mRNA was highly upregulated after TSA treatment in ADSCs (Fig. 3D), we hypothesized the involvement of BMPs and Runx2 signaling in the osteogenic effect of TSA.
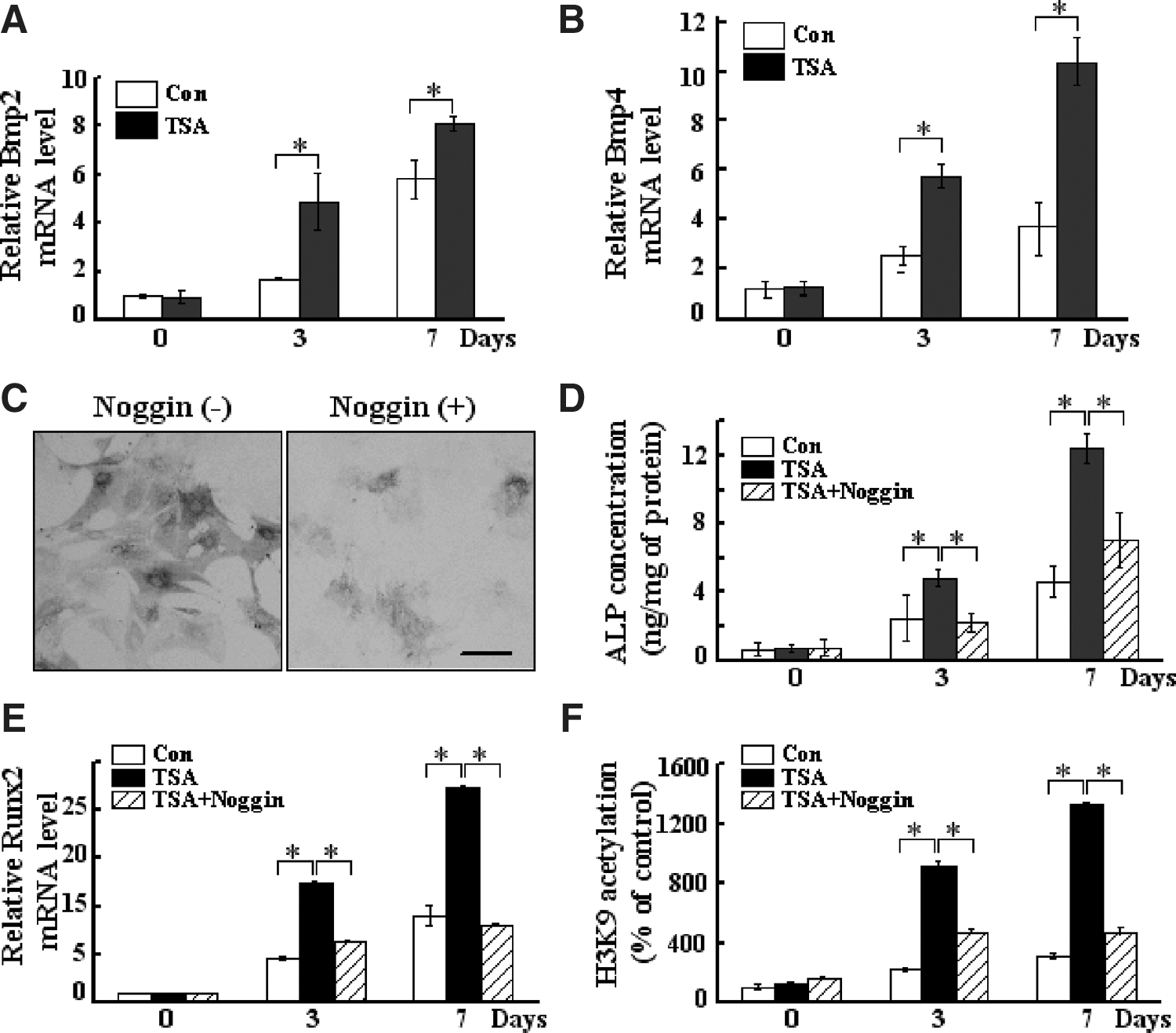
The involvement of BMP signaling in TSA effect on the osteogenesis of ADSCs.
To further test this hypothesis, we examined whether the osteogenic effect of TSA could be affected by inhibiting BMP signaling. ADSCs were treated with Noggin (Sigma), a BMP antagonist, at a concentration of 1,000 ng/mL and cultured for 24 h. Addition of Noggin into the culture medium decreased the TSA-induced ALP staining (Fig. 4C) and ALP concentration (Fig. 4D), supporting the possibility that extracellular BMP is involved in the TSA effect. The expression level of Runx2 mRNA was also decreased after the addition of Noggin, suggesting an interaction between BMP and Runx2 signaling (Fig. 4E). In addition, Noggin decreased the TSA-induced acetylated histone H3K9 recruitment to the Runx2 promoter (Fig. 4F). These results further indicate that BMP signaling participates in the TSA-induced epigenetic changes of Runx2 during osteogenesis.
Discussion
Stem cell-based regenerative medicine holds great promise for clinical applications in the repair of diseased tissues. Although MSCs have been isolated from most adult tissues, ADSCs and BMSCs are the most thoroughly investigated populations of adult stem cells for clinical applications. Compared with BMSCs, ADSCs can be easily and abundantly available from the adipose tissue and may be a promising candidate cell source for tissue engineering. Previous studies indicate the differences on the osteogenic differentiation capacity between BMSCs and ADSCs. Our present study aimed to explore its mechanism and improve the osteogenic potential of ADSCs for further tissue engineering.
Increasing evidence indicates that the multilineage differentiation ability of stem cells is defined by developmentally regulated transcription factors and of lineage specification genes [20]. Runx2 is an osteogenic specific transcription factor and essential for osteoblast differentiation and bone formation through stimulating bone marker genes transcription in osteoblasts. Overexpression of Runx2 can induce and upregulate expression of multiple osteoblast-specific genes in nonosteogenic cells. Previous studies have shown that BMSCs had a significantly high expression of Runx2 during osteogenesis compared with ADSCs, suggesting that the Runx2 expression level is highly correlated with the osteogenic differentiation ability of MSCs.
Gene expression can be regulated at the epigenetic level that includes different modifications of chromatin and such epigenetic regulation could alter the genes' accessibility to transcription factors and other regulators [21]. DNA methylation and histone modifications are the major epigenetic modifications, which are tightly linked to mammalian gene regulation and some cellular processes. Recent studies have indicated that the lineage-specific differentiation of adult stem cells could be influenced by different epigenetic states. Our results demonstrated that the degree of H3K9 acetylation and H3K4 trimethylation in ADSCs and BMSCs both increased, while the level of H3K9 trimethylation decreased (Fig. 2A), but the epigenetic changes in BMSCs were more obvious than in ADSCs. Meanwhile, the epigenetic modifications exhibited a dynamic pattern that is well correlated to the expression profile of Runx2 after osteogenic induction, especially H3K9 acetylation and trimethylation. From Fig. 1B, we can see that the expression of Runx2 was increased until day 7 in ADSCs, but was already decreased in BMSCs at this time point. Consistently, the recruitment of acetylated H3K9 on the Runx2 promoter was reduced, while the recruitment of trimethylated H3K9 was increased in BMSCs. These results indicate that histone modification may play an important role in the osteogenesis process. The difference of osteogenic differentiation capacity between ADSCs and BMSCs could be mostly attributed to the changes of epigenetic regulation on the Runx2 promoter after osteogenic induction.
Indeed, the manipulation of epigenetic modifications on the genes associated with lineage-specific differentiation will enhance the osteogenic differentiation of ADSCs. TSA, a potent HDAC inhibitor, can increase the gene transcription commonly through promoting the acetylation level of histones and chromatin relaxation, and the degree of this process is depending on the cell type, the drug dose and the treatment interval [22].
In this study, the effects of TSA on the osteogenic differentiation of ADSCs are clearly demonstrated. The treatment of TSA significantly increased the osteogenic differentiation of ADSCs. The increased level of histone H3K9 acetylation and acetylated histone H3K9 binding to the Runx2 promoter region further indicate that the upregulation of Runx2 during osteogenesis is tightly associated with the selective induction of histone hyperacetylation at the promoter region of this gene. More importantly, the osteogenic potential of TSA-stimulated ADSCs is higher than nonstimulated BMSCs as indicated by ALP staining in Fig. 3C, and the expression level of Runx2 in Figs. 1B and 3C, suggesting that histone modification of these stem cells is a major event for osteogenic differentiation on ADSCs.
BMPs are potent osteogenic agents and required for induction of osteoblast differentiation by Runx2 [19]. TSA could enhance the BMP2 expression and BMP4 activity in ADSCs at day 3 after induction. The blocking of BMP pathways by noggin could inhibit TSA-induced osteogenic differentiation, suggesting that the effect of TSA on osteogenic differentiation is, at least in part, mediated by the BMP signaling.
The findings in this study indicate that the epigenetic states are responsible for the different osteogenic differentiation capacity of ADSCs and BMSCs, and pretreatment with TSA could enhance the osteogenic potential of ADSCs in vitro, which provides useful information for further clinical application in ADSC-based bone tissue regeneration.
Footnotes
Acknowledgments
This work was supported by Specialized Research Fund for the Doctoral Program of Higher Education (20110001130001), the National Natural Sciences Foundation of China (90919022, 81101350, 81070112, 81071675), the 111 Project of China (B07001), China Postdoctoral Science Foundation (20110490249).
Author Disclosure Statement
No competing financial interests exist.