
Abstract
Although transplantation of neural stem/progenitor cells (NSPC) encourages regeneration and repair after spinal cord injury (SCI), the survival of transplanted NSPC is limited. Ephrin-B3 has been shown to reduce the death of endogenous NSPC in the subventricular zone of the mouse brain without inducing uncontrolled proliferation. Due to similarities in the environment of the brain and spinal cord, we hypothesized that ephrin-B3 might reduce the death of both transplanted and endogenous spinal cord-derived NSPC. Both normal and injured (26 g clip compression) spinal cords were examined. Ephrin-B3-Fc was tested, and Fc fragments and phosphate-buffered saline (PBS) were used as controls. We found that EphA4 receptors were expressed by spinal cord-derived NSPC and expressed in the normal and injured rat spinal cord (higher expression in the latter). In vitro, ephrin-B3-Fc did not significantly reduce the survival of NSPC except at 1 μg/mL (P<0.05), but Fc fragments alone reduced NSPC survival at all doses in a dose-dependent fashion. In vivo, intrathecal infusion of ephrin-B3-Fc increased the proliferation of endogenous ependymal cells and the proportion of proliferating cells that expressed the glial fibrillary acidic protein astrocytic marker in the injured spinal cord compared with the infusion of PBS (P<0.05). However, in the injured spinal cord, the infusion of either ephrin-B3-Fc or Fc fragments alone caused a 20-fold reduction in the survival of transplanted NSPC (P<0.001). Thus, after SCI, ephrin-B3-Fc and Fc fragments are toxic to transplanted NSPC.
Introduction
T
Ephrin ligands and Eph receptors represent the largest sub-family of tyrosine receptor kinases in the nervous system [27,28]. Ephrin-Eph signaling pathways are important during the development of the central nervous system for axon path-finding and branching, cell migration, and maintenance of regional patterning [28 –30]. Recently, ephrins have been shown to regulate the NSPC population in the subventricular zone (SVZ) of the adult mouse brain [25,31,32]. In particular, ephrin-B3 was shown to have anti-apoptotic properties [26,33]. The ephrins-Ephs are subdivided into families A and B; however, binding within and between families can be very promiscuous [27,28]. For example, ephrin-B3 has been found to bind to EphA4, EphB1, EphB2, and EphB3 [28], and its anti-apoptotic effects have been observed in interactions with EphA4 receptors in cell culture and in the mouse SVZ [25,26]. EphA4 receptors are expressed by NSPC from the mouse brain [26,31,34] and are up-regulated in the injured spinal cord [35 –39]. However, there are no previous reports on the effect of ephrin-B3 on the survival of endogenous or transplanted spinal cord-derived NSPC.
Based on the similarities between the environment of the brain and the spinal cord [40], we investigated the effect of ephrin-B3 on adult rat spinal cord-derived NSPC in culture and in vivo. We assessed whether ephrin-B3 improved the survival of spinal cord-derived NSPC in vitro, endogenous ependymal cells in vivo, and transplanted spinal cord derived NSPC in the injured spinal cord. We administered ephrin-B3 locally by intrathecal (IT) infusion to adult rats with SCI produced by clip compression, which simulates clinical SCI [41]. We found that ephrin-B3 slightly altered the survival of spinal cord-derived NSPC in vitro and slightly increased the proliferation of endogenous ependymal cells in the injured rat spinal cord. Notably, we found that ephrin-B3 was toxic to adult rat spinal cord-derived NSPC transplanted into the injured rat spinal cord.
Methods
Animals
Adult female Wistar rats (250–300 g) were used in all experiments (n=61 total). Transgenic rats expressing green fluorescent protein (GFP) were used for harvesting NSPC (YS Institute, Inc.) [42], and wild-type adult female Wistar rats (6–8 weeks old; Charles River) were used for all other experiments. All animal procedures were approved by the Animal Care Committee of the Research Institute of the University Health Network in accordance with the Guide to the Care and Use of Experimental Animals (Canadian Council on Animal Care). Table 1 shows the treatment groups and the respective number of animals used.
PBS, phosphate-buffered saline; NSPC, neural stem/progenitor cells; IT, intrathecal; Fc, crystallizable fragment of immunoglobulin G. N/A, not applicable.
Neural stem/progenitor cells
NSPC were derived from the central canal region of the spinal cord and from the periventricular region of the lateral ventricles of the brain of adult GFP-positive (GFP+) transgenic rats. The procedures for the harvesting and passaging of NSPC in our laboratory have been previously described by Mothe et al. [19] and Guo et al. [24]. Briefly, the spinal cord and brain were excised under sterile conditions. The spinal cord was dissected so that only the central canal region (including the ependymal, subependymal and some gray matter surrounding the central canal) remained, and the brain was dissected so that only the periventricular tissue remained. The excised tissues were divided into small pieces and dissociated with 0.01% papain and 0.01% DNAse I at 37°C. The cells obtained were resuspended in serum free media with 20 ng/mL of mouse epidermal growth factor (EGF), 20 ng/mL of human basic fibroblast growth factor (FGF2), and 2 μg/mL of heparin (all from Sigma-Aldrich). The cells were cultured as neurospheres and passaged every 7 days.
Spinal cord injury
Adult rats were anesthetized by inhalation of 4% isofluorane and a mixture of 1:2 nitrous oxide and oxygen, and then maintained at 2% isofluorane during surgery. The spinal cord was exposed through a midline incision, and a laminectomy was performed at levels T7 and T8 (Fig. 1). The spinal cord was then compressed at T7/8 by a 26 g clip for 1 min [43]. Animals used for assessing endogenous ependymal cells with IT infusion in the normal spinal cord received only laminectomy (no clip compression). Two additional animals used for assessing endogenous ependymal cells without IT infusion in the normal spinal cord did not receive any manipulations (no laminectomy or clip compression).

Schematic diagram describing the relative positions of the clip compression injury, the injection sites for cell transplantation, and the placement of the intrathecal catheter. The distances are not to scale. The arrowheads show the location of the neural stem/progenitor cells (NSPC) injection sites rostal and caudal to the injury site. The suture immobilizes the catheter preventing movement. (Modified from Parr et al. [23])
Transplantation
Seven days after the initial SCI, animals were anesthetized again by inhalation of 4% isofluorane and a mixture of 1:2 nitrous oxide and oxygen and then maintained at 2% isofluorane in order to re-expose the injury site. GFP+ neurospheres were injected approximately 1 mm rostral and 1 mm caudal to the injury site (Fig. 1). One rostral and one caudal injection were made in the midline at a depth of 1–1.5 mm into the spinal cord with a 32-gauge needle (Hamilton 7803–04; 32 gauge with 20° bevel) attached to a Hamilton syringe (Hamilton). At each injection site, 2 μL containing 200000 cells were injected at a rate of 2.5 μL/min with a motorized pump (Model 780310; Stoelting) [19]. Starting from the day of transplantation, animals were immunosuppressed daily with 15 mg/kg of cyclosporine (Sandimmune, Novartis) until they were sacrificed. Animals used for assessing endogenous ependymal cells in the normal and injured spinal cord did not receive NSPC transplantation. These animals were immunosuppressed daily with 15 mg/kg of cyclosporine starting from the date of IT infusion until they were sacrificed. Two additional animals used for assessing endogenous ependymal cells without IT infusion in the injured spinal cord did not receive NSPC transplant. These 2 animals were sacrificed 7 days postinjury.
Preclustering
Ephrin-B3 was purchased in the form of ephrin-B3-Fc (R&D Systems). The human immunoglobulin G Fc fragments (IgG Fc) (Jackson ImmunoResearch) were used as a ligand control for the ephrin-B3-Fc. The soluble forms of ephrin-B3-Fc and Fc fragments have a very minimal effect on receptor activation [44], and, therefore, these compounds were preclustered using anti-Fc antibodies (Fitzgerald Industries) at room temperature for 3 h at a ratio of 1:10 (ligand: anti-Fc), as previously described [25,45].
Intrathecal infusion
Immediately after the transplantation of NSPC, preclustered ephrin-B3-Fc or Fc fragments were delivered intrathecally through a polyurethane catheter (Alzet Model No. 0007741; Alzet Osmotic Pumps) attached to a mini-osmotic pump (Alzet Model No. 1007D) (Fig. 1). 100 μL of preclustered ephrin-B3-Fc or Fc fragments (100 μg/mL) were injected into the mini-osmotic pump. An equal volume of phosphate-buffered saline (PBS) was used as a control in animals not receiving preclustered ephrin-B3-Fc or Fc fragments. The pump was primed in sterile saline at 37°C overnight. A small midline durotomy was made at T9 through which the catheter was inserted into the IT space [23]. The tip of the catheter was directed rostrally to the T7 level, which was approximately 1 mm rostral to the site of the rostral injection of NSPC. The dura was then sealed around the catheter by fibrin glue (Beriplast, Beriplast, CSL Behring), and the catheter and pump were sutured to the subcutaneous tissues. Preclustered ephrin-B3-Fc, preclustered Fc fragments, or PBS were then intrathecally delivered continuously for 3 or 7 days (0.5 μL/h) via the catheter and pump system.
Histology, immunohistochemistry, and image analysis
Animals were sacrificed with a lethal intraperitoneal injection of sodium pentobarbital, and then received perfusion with 4% paraformaldehyde for fixation. The spinal cord was excised and maintained in 30% sucrose at 4°C. A 1 cm segment of spinal cord was mounted, cryosectioned parasagittally into 20 μm sections, and collected on Superfrost slides (Fisher Scientific). The following antibodies were used for immunofluorescent staining: anti-EphA4 (1:100; Santa Cruz Biotechnology, Inc.) for EphA4 receptors, anti-Ki67 (1:100, Novacastra) for proliferating cells, anti- glial fibrillary acidic protein (GFAP) (1:500; Chemicon) for astrocytes, and anti-CC1/APC (1:1000; Calbiochem) for oligodendrocytes. During immunostaining, sections were rehydrated with 0.1 M PBS and blocked with normal goat serum in 0.1 M PBS for 1 h. The sections were incubated with primary antibody at 4°C overnight, washed with 0.1 M PBS for 30 min, and then incubated with secondary antibody conjugated to Alexa Fluor 568 (1:500) or Alexa Fluor 488 (1:500) (all from Millipore) for 1 h. Negative controls were obtained by omission of the primary antibody. Sections were counterstained with the 4′,6-diamidino-2-phenyl-indole (DAPI) nuclear counterstain (Vector Laboratories).
For quantitative cell survival analysis in tissue sections, GFP+ cells were examined in every 8th section [19]. Images of each section were captured using a Zeiss LSM 510 confocal microscope, and GFP+ cells were counted in a total of 15 sections (every 8th section, 140 μm apart, was selected to avoid double counting of cells appearing on more than one section). The total cell count was estimated using the Abercrombie method [46] for the entire cord thickness. For quantitating the Ki67-labeling index in the ependymal region, a total of 15 sections were selected in a similar fashion to the cell survival analysis. Images were captured on sections containing the ependymal region. The Ki67-positive (Ki67+) and DAPI-stained cells in the ependymal region were counted, and the Ki67+ cells were expressed as a percentage of the DAPI-stained cells. To quantitate the number of Ki67+ cells that express either GFAP or CC1 markers, a total of 15 sections were selected in a similar fashion to the cell survival analysis. Four regions (368×368 μm rectangular fields) of each section were selected in the parasagittal tissue sections encompassing the areas adjacent to the site of injury and extending rostrally and caudally approximately 1.05 mm. Images of each region were captured using a Zeiss LSM 510 confocal microscope. Ki67+ and double-labeled Ki67+/GFAP+ or Ki67+/CC1+ cells were counted, and the double-labeled cells were expressed as a percentage of the Ki67+ cells.
The ImageJ software developed by the National Institutes of Health was used to analyze the intensity of the GFAP immunostaining in tissue sections of animals that received transplanted NSPC and 7 days of IT delivery of PBS, Fc fragments, or ephrin-B3-Fc. 15 sections were selected in a similar fashion to the cell survival analysis, and 4 regions were selected similar to the quantitation of Ki67+ cells that express either GFAP or CC1 markers. Images of each region were captured using a Zeiss LSM 510 confocal microscope; these images were thresholded using the binary (Black/White) option in the ImageJ software, and the integrated density was calculated by the software. These densities were then averaged and analyzed.
Cell culture, survival, and proliferation assays
Cultured GFP+ NSPC were dissociated and seeded in Matrigel™ (Growth Factor Reduced BD Matrigel Matrix and diluted by a factor of 25; BD Biosciences) coated wells in 24- and 96-well plates with serum-free media containing 20 ng/mL of EGF, 20 ng/mL of FGF, and 2 μg/mL of heparin for 24 h [19]. Thereafter, the media was changed to serum-free media with ephrin-B3-Fc or Fc fragments (0.1 μg/mL, 1 μg/mL, 2 μg/mL, 10 μg/mL, and 100 μg/mL) for 48 h (Table 2). The control wells contained serum-free media with an equal volume of PBS in place of the preclustered ephrin-B3-Fc or Fc fragments (labeled as the 0 μg/mL condition). The MTS tetrazolium salt [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium)] colorimetric assay was used to assess cell survival (CellTiter 96® AQueous Nonradioactive cell proliferation assay; Promega). Briefly, after 48 h of incubation with preclustered ephrin-B3-Fc or Fc fragments or PBS in serum-free media, 20 μL of the MTS/PMS solution was added per 100 μL of NSPC in media (resulting in a final concentration of 333 μg/mL MTS and 25 μM of PMS). The resulting mixture was incubated at 37°C with 5% CO2 for 1 h, and the absorbance was recorded at 490 nm using an UV plate reader. Each treatment was repeated in 4 wells, and the entire assay was repeated thrice.
MTS, 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium); EGF, epidermal growth factor; FGF2, basic fibroblast growth factor.
Cell proliferation was further analyzed through Ki67 immunocytochemical staining (Table 2), as previously described by Mothe et al. [19]. Briefly, after 48 h of incubation with preclustered ephrin-B3-Fc or Fc fragments or PBS in serum-free media, cultures were fixed in 4% paraformaldehyde for 20 min at room temperature. The cultures were washed with 0.1 M PBS, blocked with 10% normal goat serum for 1 h at room temperature, and incubated with the primary antibody anti-Ki67 (1:100) overnight at 4°C. Cultures were washed with 0.1 M PBS and incubated with fluorescent Alexa 568 secondary antibody (Millipore) for 1 h. Thereafter, cultures were washed with 0.1 M PBS and counterstained with Hoechst (Invitrogen) to identify the nuclei. Each treatment was repeated in 4 wells, and the entire assay was repeated thrice. In a quantitative analysis of the Ki67-labeling in cell cultures [18], 5 fields at 20× magnification (710×530 μm rectangular fields) were selected from each well; immunofluorescent cells were examined using a Nikon Eclipse TE 300 microscope, and images were captured with a CCD camera and Bioquant Software (R&M Biometrics, Inc.). Ki67+ and Hoechst labeled cells were counted, and the Ki67+ cells were expressed as a percentage of the Hoechst-labeled cells.
Statistical analysis
All data were presented in the form of mean±standard error (m±se) and were analyzed using SigmaStat 3.1 (Systat). Statistical differences between groups were examined with one-way ANOVA and Bonferroni-adjusted pairwise multiple comparisons. In cases where only 2 experimental conditions were examined, statistical differences were evaluated with the Student's t-test. In all statistical tests, P<0.05 was set as the significance level, unless otherwise indicated.
Results
Cultured NSPC and normal and injured spinal cord tissue express the EphA4 receptor
Immunofluorescent staining showed that cultured adult rat brain (Fig. 2A–C) and spinal cord-derived NSPC (Fig. 2D–F) express the EphA4 receptor. We did not find a significant difference between brain and spinal cord-derived NSPC in culture (80.95%±2.42% of brain NSPC showed EphA4 expression and 76.84%±3.85% of spinal cord NSPC showed EphA4 expression). After transplanting into an injured spinal cord, many GFP+ spinal cord NSPC showed EphA4 expression at 3 and 14 days post-transplant (data not show). Thus, cultured adult spinal cord NSPC express the receptor that is capable of responding to the ephrin-B3 ligand. We found that EphA4 receptors were also expressed in vivo in the normal (Fig. 2H–L) and injured spinal cord (Fig. 2M–Q). The injured spinal cord showed a higher level of expression than the normal spinal cord, suggesting that the EphA4 receptor is up-regulated after SCI. Double immunofluorescence staining showed co-localization of EphA4 with GFAP or CC1 in both the normal and injured spinal cord (Fig. 2K, L, P, Q).
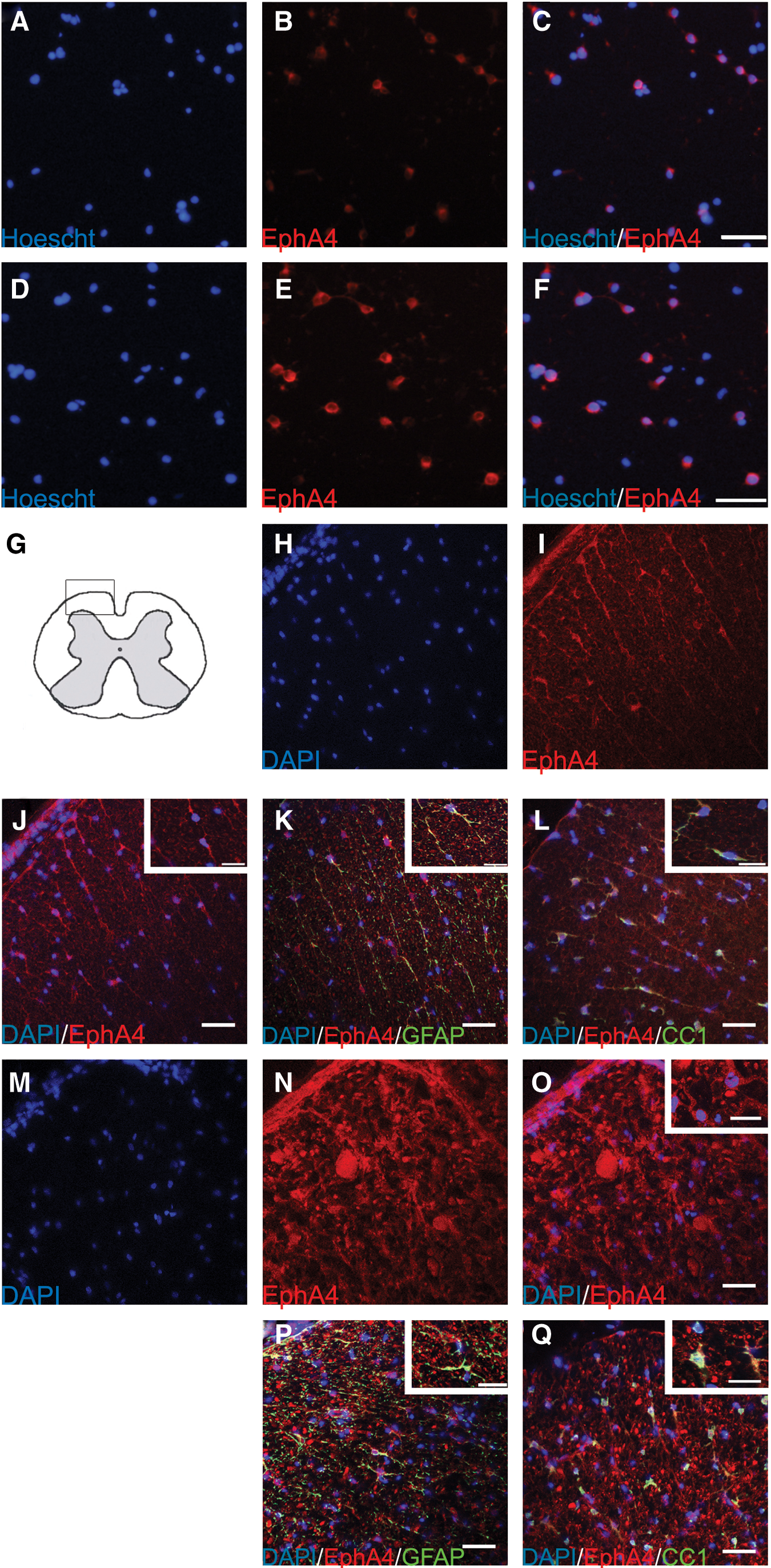
Immunostaining of the EphA4 receptor in cultured brain and spinal cord-derived NSPC
Ephrin-B3 did not significantly alter NSPC survival in vitro
To investigate the effect of ephrin-B3 in vitro, ephrin-B3-Fc was added to the NSPC in culture, with Fc fragments and PBS serving as controls (Table 2). Based on the MTS assay, most concentrations of ephrin-B3-Fc had no effect on NSPC survival, although there was a small increase in survival at 1 μg/mL (P<0.05) (Fig. 3B). In contrast, Fc fragments decreased NSPC survival starting at 2 μg/mL with progressively greater toxicity in increasing concentrations (P<0.05) (Fig. 3A).

Survival and proliferation of dissociated spinal cord-derived NSPC in culture with ephrin-B3-Fc or crystallizable fragment of immunoglobulin G (Fc) fragments for 3 days. Survival of NSPC is assessed by the MTS assay (m±se) with the addition of Fc fragments
Since the MTS assay did not differentiate between the effects of the compounds on NSPC survival or those on proliferation, further testing was performed using Ki67 labeling. Figure 3C and D shows that ephrin-B3-Fc or Fc fragments did not alter proliferation compared with the PBS control except at the highest concentration (100 μg/mL), where proliferation was significantly decreased (P<0.001). These results suggest that ephrin-B3-Fc had a minimal effect on NSPC survival in vitro and no effect on NSPC proliferation (except toxicity at a very large concentration) in vitro, and that Fc fragments decreased NSPC survival but had no effect on NSPC proliferation (except at a very large concentration) in vitro.
Ephrin-B3 slightly increased the proliferation of endogenous periventricular ependymal cells in the injured adult spinal cord
We examined the effect of ephrin-B3 on periventricular cells in the normal and injured adult spinal cord. These periventricular cells are known to contain stem/progenitor cells [3,6,18]. Ephrin-B3-Fc (100 μg/mL), Fc fragments, (100 μg/mL) or PBS as a negative control were infused intrathecally for 7 days (n=5/group). In the normal spinal cord (Fig. 4A), no significant difference was observed among the 3 groups in the percentage of Ki67+ cells. In the injured spinal cord (Fig. 4B), a small significant increase was observed in the ephrin-B3-Fc group in comparison to the PBS group (P<0.05). However, the difference between ephrin-B3-Fc and Fc fragments groups did not reach significance, and no significant difference was observed between the Fc fragments and PBS groups. The Ki67 labeling in the injured spinal cord (Fig. 4B) was re-analyzed based on distances rostral and caudal to the injury site (Fig. 4C–E). The Ki67 labeling between the same lengths of the cord, which were of equal distance rostral or caudal from the injury site (e.g., 1.2 mm vs. −1.2 mm), were compared. In the PBS and Fc fragments groups, Ki67 labeling in the range of 1.8 mm to 3.5 mm rostral to the injury site was significantly higher than the labeling at the respective caudal distances (P<0.001). In the ephrin-B3-Fc group, Ki67 labeling in the range of 1.2 mm to 4.1 mm rostral to the injury site was significantly higher than the labeling at respective caudal distances (P<0.001). Moreover, for all 3 groups, Ki67 labeling was concentrated at approximately 1.8 mm to 2.9 mm with the highest labeling at approximately 2.3 mm rostral to the site of injury, which corresponds with the approximate location of the tip of the IT catheter. At approximately 2.3 mm rostral to the injury site, Ki67 labeling was significantly higher in the ephrin-B3-Fc and Fc fragments groups than in the PBS group (P<0.001). Furthermore, we observed a higher number of proliferating cells in all the animals with IT catheter implantation than in those without catheters for both the normal and injured spinal cord (Fig. 5A–D).

Effect of intrathecal delivery of ephrin-B3-Fc or Fc fragments on the endogenous spinal cord ependymal cells. Ephrin-B3-Fc or Fc fragments were delivered for 7 days to normal and spinal cord injury (SCI) rats, and Ki67 labeling was used to measure proliferation of the endogenous spinal cord ependymal cells. Data are reported as m±se, and asterisk denotes a significant difference.
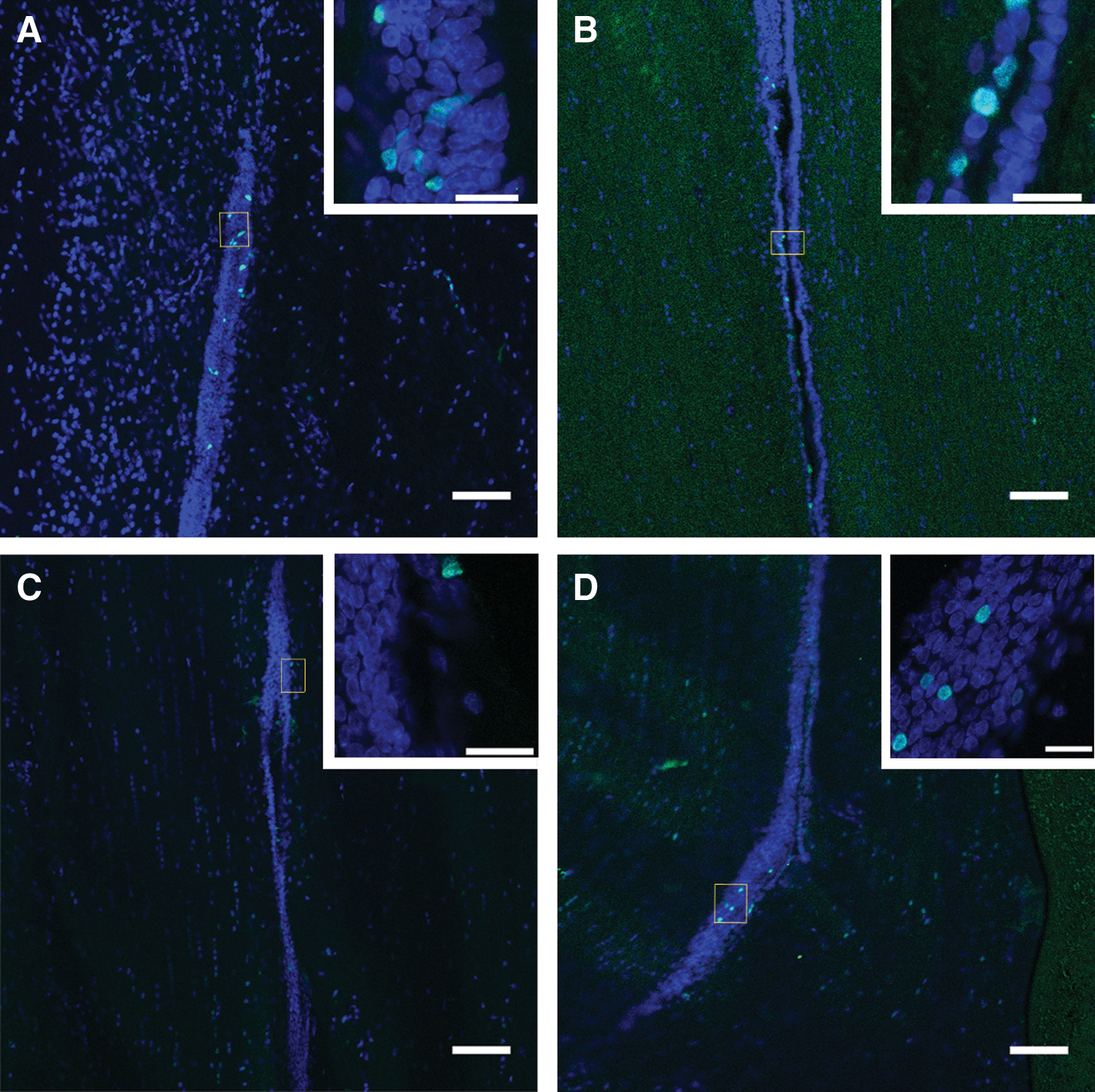
Sample images showing cells labeled for Ki67 confined to the ependymal region of the spinal cord. Merged confocal images showing Ki67+ cells counterstained with 4′,6-diamidino-2-phenyl-indole (DAPI).
Ephrin-B3-Fc was toxic to transplanted adult spinal cord-derived NSPC
Ephrin-B3-Fc (100 μg/mL), Fc fragments (100 μg/mL), or PBS as a negative control were infused intrathecally for 3 (n=3/group) or 7 (n=6/group) days. In all the 3-day IT infusion groups, many GFP+ NSPC were observed (Fig. 6A–C). There was no statistical difference in the number of surviving GFP+ NSPC between the 3 groups, although there was a trend toward slightly lower GFP+ NSPC survival in the ephrin-B3-Fc and Fc fragment groups (Fig. 6G; PBS group had a survival of 4.07%±1.50%, Fc fragments alone group had a survival of 2.97%±0.88%, and ephrin-B3-Fc group had a survival of 3.25%±0.93%). Interestingly, in the ephrin-B-Fc and Fc fragments groups, the transplanted NSPC were more localized to the sites of injection in comparison to the PBS group, in which the cells were more widely dispersed (Fig. 6A–C). In contrast, in the 7-day IT infusion groups, only a small number of surviving transplanted NSPC were observed in the ephrin-B3-Fc and Fc fragment groups (Fig. 6D–F); large clusters of degraded GFP+ cell debris were observed in localized collections at the sites of injection, and some GFP+ remnants were observed in macrophages (Fig. 6E–F). Compared with the PBS group, we observed a 20-fold decrease in the percentage of surviving NSPC in the ephrin-B3-Fc and Fc fragment groups (Fig. 6H). The PBS group had a survival of 2.19%±0.47% with all 6 animals showing live transplanted NSPC. In contrast, the ephrin-B3-Fc group had a survival of 0.15%±0.10% with only 3 out of 6 animals showing any surviving transplanted NSPC, and the Fc fragments group (n=5; one animal died during the transplantation surgery) had a survival of 0.11%±0.10% with only 2 out of 5 animals showing any surviving transplanted NSPC.

Effect of intrathecal delivery of PBS
Ephrin-B3 increases the percentage of proliferating cells that express the GFAP astrocytic marker in the spinal cord parenchyma
Through immunofluorescent staining, we showed that EphA4 receptors are also expressed by endogenous astrocytes and endogenous oligodendrocytes. To further analyze the effect of ephrin-B-Fc on these cells that endogenously express the EphA4 receptor, we assessed the number of proliferating cells (as labeled by Ki67) that express the GFAP astrocytic (Ki67+/GFAP+; Fig. 7A) or CC1 oligodendrocyte marker (Ki67+/CC1+; Fig. 7B). We analyzed these markers in ependyma and parenchyma, including the perimeter, of the normal and injured spinal cord. The Ki67+ cells in the normal and injured spinal cord ependyma did not show any GFAP or CC1 staining (data not shown), indicating that the cells in the ependyma region are in an undifferentiated state in agreement with previous literature [4,6,10]. In the parenchyma of the normal spinal cord, we found a significant increase in the percentage of Ki67+cells that expressed the GFAP marker with the delivery of ephrin-B3-Fc compared with PBS or Fc fragments (P<0.05). In the parenchyma of the injured spinal cord, we also found a significant increase in the proportion of Ki67+ cells that expressed the GFAP marker (expressed as % Ki67+/GFAP+cells) with the delivery of ephrin-B3-Fc compared with the delivery of PBS (P<0.05), but not compared with Fc fragments. No significant difference was observed between the delivery of PBS or Fc fragments in both normal and spinal cord. Moreover, there was no significant difference in the percentage of Ki67+ cells that expressed the GFAP marker before and after injury, although a trend toward increased expression of GFAP after injury was observed. With regard to the proportion of Ki67+cells that expressed the CC1 marker (expressed as % Ki67+/CC1+cells), we did not observe a significant difference among the 3 IT delivery groups before or after injury. This result suggests that it is possible that the delivery of ephrin-B3-Fc mainly affected cells of an astrocytic lineage or drove undifferentiated cells into an astrocytic rather than an oligodendrocyte lineage.
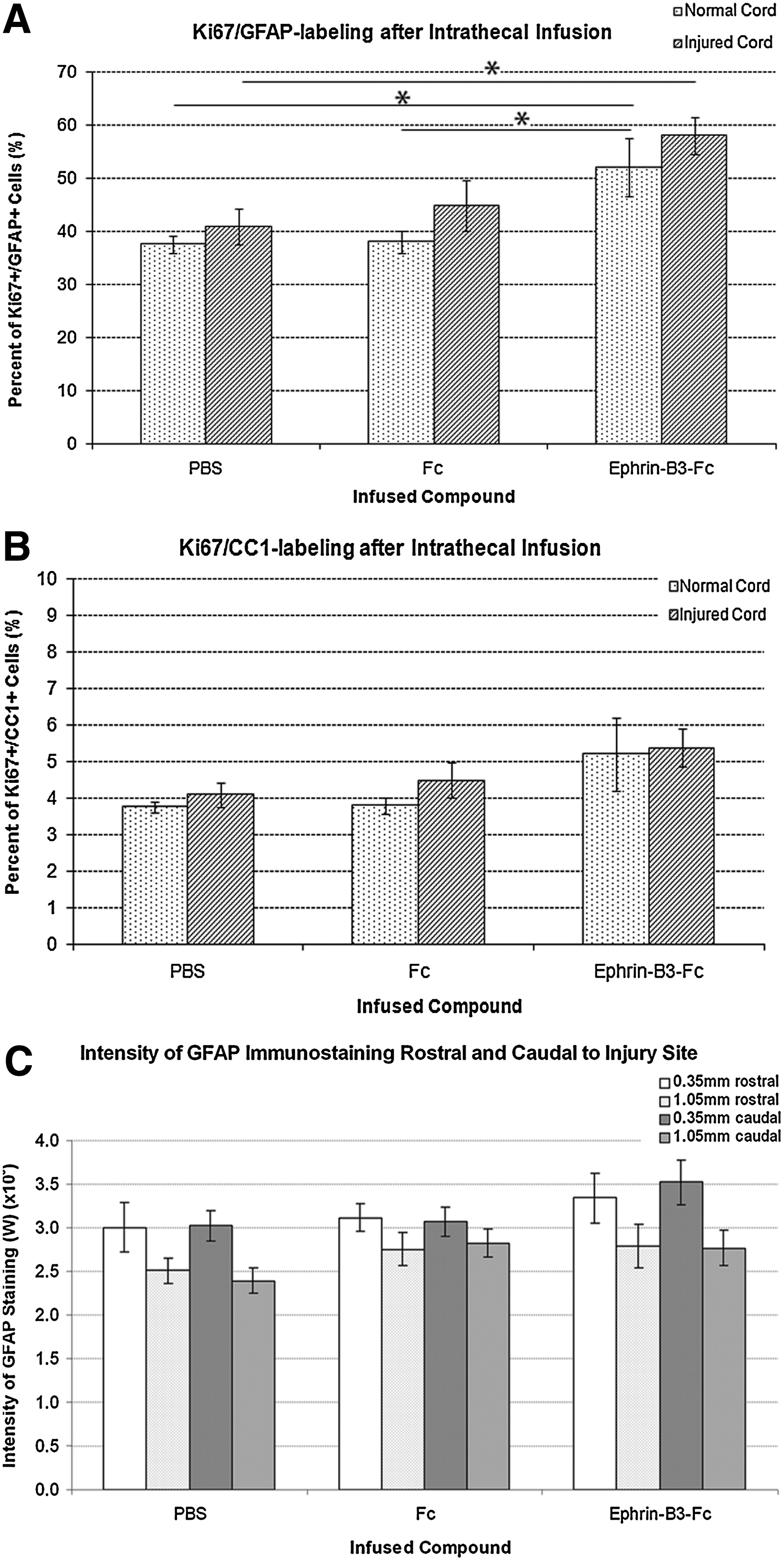
Effect of IT delivery of ephrin-B3-Fc or Fc fragments on the percentage of proliferating cells that express the glial fibrillary acidic protein (GFAP) astrocyte
Since ephrin-B3-Fc increased the percentage of Ki67+cells that expressed the GFAP marker, we assessed the possibility that IT delivery of ephrin-B3-Fc could have contributed to astrogliosis, which could render the environment hostile for cell survival [2,13]. We measured the intensity of the GFAP immunostaining in tissue sections of animals that received transplanted NSPC. However, although a trend toward higher GFAP staining was observed with the delivery of ephrin-B3-Fc, no significant difference was observed across the 3 IT delivery groups (Fig. 7C).
Discussion
Adult spinal cord-derived NSPC are multipotent and have regenerative potential in culture and after transplantation into animals with SCI [13,14,18 –20]. However, in these studies, the percentage of surviving transplanted NSPC has been small; for example, Parr et al. [20] reported that only 1.2% of transplanted NSPC survived at 12 weeks post-transplant. To augment the survival of transplanted NSPC, the infusion of growth factors has led to promising results. For instance, Karimi-Abdolrezaee et al. [13] increased the survival of transplanted NSPC to 37% at 8 weeks post-transplant through IT administration of EGF, FGF2, and platelet-derived growth factor for 7 days. Due to the potential side effects of growth factors, such as uncontrolled proliferation [22 –24], we sought alternative measures to enhance transplant survival and selected ephrin-B3, because it improved the survival of brain-derived NSPC [25,26,33].
First, we demonstrated that adult spinal cord-derived NSPC express EphA4 receptors which are capable of binding ephrin-B3 ligands, similar to its expression in brain-derived NSPC [26,31,47] (Fig. 2). We found no statistical difference between brain and spinal cord-derived NSPC in the percentage of cells expressing EphA4, suggesting that the brain and spinal cord NSPC are equally likely to respond to ephrin-B3.
In culture, we showed that ephrin-B3-Fc exerted small effects on spinal cord NSPC survival, but Fc fragments reduced NSPC survival in a dose-dependent manner (Fig. 3A, B). The observation that Fc fragments had toxic effects on NSPC was surprising, because Fc fragments have been used as a control for various -Fc recombinant fusion proteins in studies examining cells of neural origin and, to our knowledge, no toxic effects have been reported [26,32,33,45]. One possible explanation is related to the fact that the Fc fragment is a crystallizable region of the immunoglobulin (we used the Fc fragments of immunoglobulin G [IgG]) [48,49]. Notably, the application of IgG in cell cultures, including rat and human neuroblastoma cell lines, reduced cell survival and proliferation in a dose-dependent manner [50]. Moreover, Ekberg et al. [51] reported that, in peripheral blood mononuclear cells, the effect of IgG on cell apoptosis changes over time. Ekberg et observed an increase in cell death in the presence of IgG between 24 h and 72 h in culture; thereafter, the presence of IgG significantly decreased cell death. In our experiments, we assessed cell survival at only 72 h (after the addition of Fc fragments), which coincides with the time of increased apoptosis reported by Ekberg et al. [51]. In addition, in contrast to previous studies with brain NSPC [26,32,33], we removed the growth factors from the culture media before the addition of the Fc fragments. The absence of growth factors probably produced an environment that was less conducive to cell growth and survival, and could have rendered the NSPC more susceptible to the apoptotic effects of the Fc fragments. Thus, our experimental conditions (culture media in the absence of serum and growth factors) could have favored the inherent apoptotic effect of the Fc fragments. To our knowledge, ephrin-B3-Fc is currently the only commercially available form of ephrin-B3. Since we tested ephrin-B3-Fc, we were not able to differentiate the possible confounding effects of the -Fc fragments in the ephrin-B3-Fc. Different from Fc fragments, we showed that ephrin-B3-Fc did not have toxic effects on the survival of spinal cord NSPC in vitro except at the highest concentration tested. We think that either the clustering of ephrin-B3-Fc might have caused the -Fc component to be less readily available for binding with other receptors or the binding of ephrin-B3 with EphA4 might have counteracted the apoptotic effect of the -Fc component.
We also examined the effect of ephrin-B3-Fc and Fc fragments on the endogenous stem/progenitor cells in the ependymal region from which we derived the spinal cord NSPC (Fig. 4) (We performed Western blotting on ephrin-B3-Fc and Fc fragments after incubation at 37°C for 7 days and did not find degradation or denaturing of the proteins [data not shown], indicating that the proteins are likely to be functional). In the normal spinal cord, IT infusion of ephrin-B3-Fc or Fc fragments for 1 week did not result in any significant difference in the percentage of proliferating ependymal cells in comparison to the infusion of PBS. Interestingly, we observed a higher percentage (approximately 7%) of proliferating cells in the ependymal region of normal rats in all 3 treatment groups (in comparison to our previous report of approximately 2% proliferating cells) [6]. This increased proliferation (Fig. 5A–D) is probably related to the trauma from the insertion of the catheter for IT delivery (including the effects of laminectomy and the pressure of the catheter on the adjacent spinal cord), and other possible effects on the adjacent tissue due to the continued presence of the catheter [10,23,52 –54]. For example, our laboratory has reported proliferative mass lesions arising from the meninges associated with the use of IT catheters [23,24].
In the injured rat spinal cord, we found that the infusion of ephrin-B3-Fc for 1 week significantly increased the percentage of proliferating cells in the ependymal region in comparison to the infusion of PBS (Fig. 4B). Figure 4C–E shows significantly higher Ki67 labeling rostral to the injury site compared with respective caudal distances. This observation further illustrates the trauma resulting from the presence of the IT catheter. In addition, Ki67 labeling was mainly concentrated rostrally with the maximum at approximately 2.3 mm rostral to the site of injury, which coincides with the location of the tip of the IT catheter. In comparison to the PBS group, the ephrin-B3-Fc and Fc fragments groups showed a higher percentage of Ki67 labeling in the same region, most likely reflecting an additive effect of the catheter tip and presence of the infused compounds. There was a lower percentage of Ki67+ cells in the injured spinal cord compared with the normal cord (Fig. 4A, B), perhaps due to the relatively short rostral-caudal length of injured spinal cord tissue that was examined. In SCI, central regions of the spinal cord containing the ependyma are severely damaged, and some damaged areas may have been dislodged and lost during processing. Hence, only limited areas of tissue rostral or caudal to site of SCI could be examined, whereas in the normal spinal cord, the entire length of the ependymal region was intact. Thus, it is possible that if longer rostral-caudal segments of injured tissue were analyzed, we would have seen more areas of the ependyma and a higher percentage of Ki67+ cells.
With regard to the proliferation of NSPC, our results in rat spinal cord differ from those previously reported in the SVZ of the mouse brain. For example, in the brain SVZ of genetically modified mice [25,33], enhanced proliferation was observed in mice with absent ephrin-B3 created through gene-targeted deletion. However, we observed the opposite effect, because the infusion of ephrin-B3-Fc increased the percentage of proliferating cells in the spinal cord ependyma (Fig. 4B: 1.5% vs. 0.5%). Nevertheless, it should be noted that there were many differences between the present study and previous reports that could explain the disparity in the findings. First, more recent studies have reported differences in response between brain and spinal cord-derived NSPC to a wide range of molecules [55 –57], and ephrin-B3, especially considering its roles in development, could be another factor that elicits differing responses in brain versus spinal cord. Second, species-specific responses to injury have been reported in mice and rat, which could account for differences in treatment outcomes [58,59]. Third, traumatic injury in genetically modified animal models may produce different effects than in normal animal models [60]. Exploration of these potential reasons for the effects observed is beyond the scope of this study. In essence, we showed that endogenous ependymal cells can be influenced in vivo through IT delivery of ephrin-B3 in the injured spinal cord.
Contrary to our aim of enhancing transplanted NSPC survival, the intrathecal delivery of ephrin-B3-Fc or Fc fragments had toxic effects on transplanted NSPC (Fig. 6H). The presence of endogenous EphA4 receptors, which we demonstrated in Fig. 2, may provide an explanation for the marked reduction in NSPC survival. There may have been nonspecific binding of the intrathecally delivered compounds to endogenous receptors in the injured spinal cord. Although we did not trace the location of ephrin-B3-Fc or Fc fragments after IT infusion, we presume that the infused compounds could have interacted with the EphA4 expressing cells in the spinal cord, which included astrocytes and oligodendrocytes (Fig. 2H–Q). If ephrin-B3 and EphA4 interaction reduces cell death, this binding of the infused ephrin-B3-Fc with EphA4 expressed by astrocytes in the injured spinal cord could have caused enhanced survival of reactive astrocytes, thereby possibly intensifying astrocytic gliosis. This increased astrogliosis could worsen the hostile injury environment for transplanted NSPC as well as block NSPC migration [2,21,35,39,61]. We assessed this possibility and found that the delivery of ephrin-B3-Fc significantly increased the percentage of proliferating cells which express the GFAP astrocyte marker (Fig. 7A). This observation suggests the possibility that ephrin-B3-Fc might have stimulated either the differentiation of ependymal cells into astrocytes or the proliferation of astrocytes. However, we did not observe a significant difference in the intensity of the GFAP immunostaining across the 3 delivery groups (Fig. 7C) perhaps due to the 1 week post-transplant sampling time. It is possible that significant differences in astrogliosis are not apparent at 1 week post-transplant.
With regard to the toxicity of the IT-delivered Fc fragments, we suspect that, similar to our in vitro results, the Fc fragments acted as a component of IgG. Lapointe et al. [62] reported that delivery of intravenous immunoglobulin (containing mostly IgG) in mice suffering from ischemia-reperfusion stroke significantly increased platelet and leukocyte accumulation in the brain. Moreover, Lapointe et al. [62] suggested that, since platelets increase Fc receptor (FcR) expression poststroke, intravenous immunoglobulin might have acted on the FcR of platelets, triggering further platelet activation and leukocyte recruitment (the latter through platelet-leukocyte interactions). Thus, we could have triggered a similar effect as that reported by Lapointe et al. [62]. Indeed, the clip compression involves brief ischemia-reperfusion trauma and triggers considerable inflammatory responses, and, thus, the IT delivery of Fc fragments could have exacerbated platelet and leukocyte recruitment initiated by the compression injury through binding with FcR. After the initiation of inflammatory responses by SCI, transplanted NSPC are targeted by macrophages as a result of their allogeneic nature [14]. Exacerbated platelet and leukocyte recruitment from the delivery of Fc fragments through Fc-FcR interactions might have contributed to the degradation of transplanted NSPC, which could account for our observation of the large collections of GFP+ remnants and cellular debris (Fig. 6B, C, E, F), resulting in a significant reduction of cell survival (Fig. 6H). Since we delivered ephrin-B3 in the -Fc form, it is possible that the -Fc component of the ephrin-B3-Fc also intensified platelet and leukocyte recruitment. Hence, the infusion of ephrin-B3-Fc may have reduced NSPC survival after transplant from the combination of aggravated astrogliosis and inflammation. However, the effect of ephrin-B3-Fc or Fc fragments on inflammation post-SCI is beyond the scope of the present study and could be examined in future studies.
In summary, we demonstrated that the survival of spinal cord-derived NSPC is affected by the presence of ephrin-B3-Fc or Fc fragments in culture and after transplantation into the injured spinal cord. With regard to ephrin-B3, we suspect that its binding with the EphA4 receptor reduces cell apoptosis (as shown in our small but positive effect in culture), but this effect indirectly contributed to astrogliosis in the injured spinal cord, thereby resulting in a poor survival of transplanted NSPC. With regard to Fc fragments, we suspect that this crytallizable component of IgG acted on FcR [48], which promoted the apoptosis of NSPC in culture and after transplantation. In the case of Fc fragements, 2 mechanisms may have reduced cell survival. First, the binding with FcR may have led to an activation of the Fas-dependent apoptotic pathway [51,63,64]. Prasad et al. [63] showed that the delivery of Fas inhibited IgG-induced apoptosis, and Yu et al. [65] showed that Fas significantly increased apoptosis of spinal cord cultures and reduced perseveration of spinal cord tissue after injury. Second, Fc fragments may have indirectly exacerbated the inflammatory response after SCI (through increasing platelet and leukocyte recruitment), thereby forming a more hostile environment for the survival of transplanted NSPC [14,62]. We feel that ameliorating the hostile environment and providing a more localized delivery of ligands are necessary to enhance the survival of transplanted NSPC. Different from the endogenous cells of the spinal cord ependymal region, transplanted NSPC are challenged by additional factors, such as the lack of scaffolds and degradation by leukocytes [14,66]. These additional factors could render them more susceptible to apoptotic factors. Furthermore, IT delivery using catheters causes pressure on the spinal cord and results in nonspecific binding as mentioned earlier [23]. These problems of IT delivery using catheters are especially problematic after SCI, because additional pressure could exacerbate the injury. In addition, the expression of receptors could be altered in the injured spinal cord (such as an up-regulation of EphA4 by astrocytes), which could intensify the effect of nonspecific ligand-receptor binding. Future studies could focus on pretreating cultured NSPC before transplant or providing a better transplant environment for cultured NSPC in order to enhance their survival after transplantation. For example, Kim et al. [67] have recently reported improved survival of transplanted NSPC in a complete spinal cord transection model by pretreating the cells with cyclic adenosine monophosphate [cAMP] and encapsulating cells in chitosan channels. Exploring the effects on transplanted NSPC has strong clinical relevance, as cell transplantation is currently one of the main interests for patients with SCI.
Footnotes
Acknowledgments
The authors thank Linda Lee and Rita van Bendegem for their technical assistance. They thank Dr. Cindi Morshead and Dr. Michael Fehlings for advice. They are grateful to Dr. David G. Wilkinson (National Institute for Medical Research, Mill Hill, London, UK) for his anti-EphA4 antibody and to CSL Behring for their fibrin glue. They thank Dr. Howard Kim for assistance with the MTS assay and Sydney Dennis-Birbaum for data collection. This project was supported by the Canadian Institute of Health Research (CIHR), the James F. Crothers Family Fellowships, the Unilever/Lipton Neuroscience Fellowship (OSOTF), and the Canadian Paraplegic Association (Ontario Branch).
Author Disclosure Statement
The authors declare that there is no conflict of interest in this article.