
Abstract
Umbilical cord blood (UCB) transplantation is associated with prolonged periods of cytopenia. Ex vivo expansion of hematopoietic stem and progenitor cells (HSPCs) is currently investigated as a mean to accelerate hematological recovery. Contrary to neutrophils, platelet recovery remains problematic. For this reason, we have developed a culture protocol promoting the expansion of megakaryocyte (Mk) progenitors. The objective of this work was to determine whether the expanded (E) UCB HSPCs could accelerate platelet recovery in vivo using a murine HSPC transplantation model. The thrombopoietic activity of UCB and mobilized peripheral blood CD34+ cells expanded under mild hyperthermia (MH, ie, 39°C) with the optimized megakaryocyte progenitor cocktail (OMPC) diverged significantly from the nonexpanded (NE) cells of origin; E cells provided rapid platelet release, while NE cells strongly contributed to platelet production past 10 days of transplantation. Consequently, the complementary of both cell sources was investigated. Cotransplantation of NE with E UCB cells significantly improved the recovery of human platelets (hPLTs) in vivo due to their complementary and synergistic thrombopoietic activities. Moreover, short-term human bone marrow (BM) reconstitution was also improved. Finally, we show that early hPLT release is dependent on Mk-primed cells and that E cells do not act as accessory cells, but have a more active role. In conclusion, hPLT recovery and short-term BM engraftment can be efficiently improved by the cotransplantation of Mk-primed UCB cells with NE HSPCs in a murine transplantation model.
Introduction
U
Cellular therapies based on the growth and differentiation potential of hematopoietic stem and progenitor cells (HSPCs) have been investigated as a mean to enhance graft success and accelerate hematological recovery (reviewed in Refs. [1,2]). The principle behind this strategy is that ex vivo expansion of HSPCs should accelerate hematopoietic recovery based on the infusion of a greater quantity of progenitors that have already initiated their differentiation program, thus shortening the time required for mature cells to be released into the circulation. Hence, tremendous resources are being devoted to develop culture protocols that would support the expansion of HSPCs ex vivo. The most common approaches include optimization of cytokine cocktails, coculture with mesenchymal stromal cells, extracellular matrix proteins coating, and gene transfer of pro-self-renewal genes (recently reviewed in Refs. [1,2]).
Results from the first human trials [6 –11] essentially set the stage for more elaborate schemes. Indeed, new emerging strategies based on improved culture procedures have now clearly demonstrated that neutrophil recovery can be accelerated by infusion of ex vivo expanded adult [12 –15] and UCB [16] HSPCs. Interestingly, Delaney and colleagues combined double UCB transplantation with ex vivo expansion [16]. In their first trial, CD34+ cells from the smallest unit were first expanded with stem cell factor (SCF), thrombopoietin (TPO), Flt-3 ligand (FL), interleukin (IL)-3, and IL-6 in the presence of fibronectin fragments and the fusion protein Delta1-IgG (directed at the notch ligand). On the day of the transplantation, a second unmanipulated unit was infused 4 h before the expanded (E) cells. The results of this trial demonstrated that the E cells strongly contributed to the recovery of neutrophils in the first month, while the nonexpanded (NE) cells provided the bulk of the long-term engraftment. Unfortunately, the impact of this strategy on platelet recovery was not discussed. Conversely, De Lima et al. presented their ongoing trial also based on the co-infusion of 2 independent units: one expanded, but without any prior hematopoietic progenitor cell selection procedure and in the presence of SCF, FL, TPO, granulocyte colony-stimulating factor (G-CSF), and mesenchymal stem cells. Their preliminary results indicated a faster recovery of neutrophils and perhaps of platelets [17]. Hence, these 2 examples demonstrate that ex vivo expansion of HSPCs can be of significant benefit for hematopoietic recovery.
Typically, cytokine cocktails used so far in such trials were developed for the expansion of HSPCs. These cocktails are composed of multiple cytokines often used at very high concentrations to stimulate cell cycle entry and expansion of primitive hematopoietic cells. The drawback of these cocktails is that support to the expansion of megakaryocyte (Mk) progenitors and/or Mks remains quite limited [18 –20]. For instance, we showed that while high concentrations of SCF strongly favor cell expansion, it has a modest influence on the expansion of Mk progenitors and Mks [20,21]. Interestingly, culture conditions favoring Mk expansion have been found to lead to increased platelet levels in transplanted mice [19,22 –24].
For several years, we have been optimizing culture conditions to enhance the expansion of Mk progenitors [20,21,25,26]. Our working hypothesis being that optimal expansion of Mk progenitors is a key component to provide rapid platelet release in vivo.
Toward this goal, we reported that incubation of UCB CD34+ cell cultures under mild hyperthermia (ie, 39°C) improved the expansion of Mk progenitors and promoted Mk differentiation ex vivo [26]. Recently, we observed with the optimized megakaryocyte progenitor cocktail (OMPC), an increase in UCB Mk progenitors expansion by 30% compared to a cytokine cocktail previously used in clinical trials (TSG; TPO, SCF, and G-CSF) [20]. OMPC also achieved the same level of CD34+ cell and myeloid progenitor expansions, while using 85% less cytokines. UCB cells expanded with OMPC were also shown to have a greater thrombopoietic potential 14 days post-transplant also [20].
In this study, we sought to establish the kinetics of platelet release between NE CD34+ cells and those expanded with OMPC under optimized condition. We demonstrate that the thrombopoietic potentials of both cell types are complementary, and that cotransplantation leads to more rapid human platelet (hPLT) reconstitution. Finally, we demonstrate that committed Mks are responsible for the early platelet engraftment, and that the E cells exert their effect in a cell autonomous fashion.
Materials and Methods
CD34+-enriched cell cultures and progenitor assays
Human UCB and mobilized peripheral blood (mPB) cells were collected after obtaining the institutional review board approval and written informed consent. The UCB CD34+ cell enrichment procedure was carried out as previously described in [20]. mPB cells were obtained from adult healthy volunteers and collected by apheresis after donor exposure to 10 μg G-CSF/kg/day subcutaneously (Neupogen, Amgen) for 4–5 days. CD34+ cells were isolated using CD34-immunomagnetic positive selection (Miltenyi Biotec, Inc., Auburn, CA) according to the manufacturer's protocol. The serum-free medium used consisted of the Iscove-modified Dulbecco's medium (Invitrogen, Burlington, Ontario, Canada), 20% serum substitute (bovine serum albumin, insulin, and transferrin; StemCell Technologies, Vancouver, Canada), 40 μg/mL of low-density lipoproteins (Sigma, St.-Louis, MO), and 5×10−5 M 2-mercaptoethanol (Sigma). CD34+-enriched cells (purity ≥75%) were placed in culture [21,27] with the cytokine cocktail OMPC (SCF 10 ng/mL, TPO 35 ng/mL, and FL 11 ng/mL) [20] at 4×104 cells/mL with equal volume medium refreshment on day 4. All cytokines were purchased at Feldan Bio (Montréal, PQ, Canada), except for FL, which was purchased at Peprotech (Rocky Hill, NJ). Cultures were incubated in a humidified atmosphere (10% CO2) at 39°C or 37°C when indicated. In some experiments, CD41+ cells enriched by positive magnetic selection using a CD41a fluorescein isothiocyanate (FITC) monoclonal antibody (Immunotech, Marseille, France) and MACS separation columns according to the manufacturer's instructions (Miltenyi Biotec, Inc.). Viable nucleated cells were enumerated using trypan blue exclusion (Invitrogen). Myeloid clonogenic progenitor cells (colony-forming cells [CFCs]) were measured using MethoCult H4436 and colony-forming-unit megakaryocytes (CFU-MK) using MegaCult-C, according to manufacturer's instructions (both StemCell Technologies).
Flow cytometry analysis
Cell staining and flow cytometry analysis by flow cytometry (FACS-Cy Flow ML; Partec, Görlitz, Germany) were performed as previously described [21]. A minimum of 20,000 and 400,000 cell and platelet events were acquired per sample, respectively. Monoclonal antibodies used consisted of anti-CD41a (GPIIb)- allophycocyanine (APC), anti-CD34-FITC, anti-CD45-APC, anti-CD235-phycoerythrin (PE), anti-CD33-APC, rat anti-mouse CD41 FITC, rat anti-mouse CD41 FITC, rat anti-mouse CD45-PE, and corresponding control isotype-matched antibodies. All antibodies were purchased from Becton Dickinson (BD) Pharmingen (Mississauga, ON, Canada), except for anti-CD34 (Immunotech, Beckman Coulter, Marseille, France).
Mouse strains and transplantation of human cells
Eight-week-old NOD.Cg-Prkdcscid Il2rgtm1Wjl /SzJ (NSG) mice were purchased at The Jackson Laboratory (Bar Harbor, ME). Sublethally irradiated (325 cGy, 137Cs) mice were transplanted intravenously with 1.25×105 UCB CD34+-enriched cells (NE mouse group) or the total progeny of 1.25×105 UCB (or 1.5×105 mPB) CD34+-enriched cells expanded (E mouse group) for 6 days (or 7 days for mPB cells). Cotransplanted mice (NE+E) received 1.25×105 UCB CD34+-enriched cells mixed with the total progeny of 1.25×105 UCB CD34+-E cells. Each cohort contained a total of 5–11 mice per experiment. Independent experiments were carried out with different UCB and mPB cell preparations. In each independent experiment, all cohorts compared were injected with cells derived from the same CD34+ cell preparation. Mice injected with saline solution served as negative control for hPLT and bone marrow (BM) engraftment measurements.
Analysis of platelets in transplanted mice
Retro-orbital venous plexus blood was collected from anesthetized mouse with EDTA-coated capillaries (Drummond Scientific, Broomall, PA). Platelets analyses were done in 2 steps. First, the concentration of murine platelets was measured in whole blood by staining murine platelets with a rat anti-mouse CD41 FITC Ab. Samples were then brought to a final dilution 1/30,000, and the concentration of CD41+ murine platelets was measured by cytometry (Partec). Second, the proportion of human and murine platelets was determined in platelet-rich plasma (PRP). PRP was prepared by diluting whole blood (1/2) with PBS and microcentrifuging for 30 s at 146 g. Ten microliters of PRP (upper phase) was stained with mouse anti-human CD41a-APC and rat anti-mouse CD41 FITC species-specific antibodies (BD Pharmingen). At least 400,000 platelet events were then acquired for each sample by cytometry. The specificity of each antibody was confirmed in vitro on human and murine PRPs (data not shown) as well as into transplanted recipients. The background level for hPLT detection in control mice (1 hPLT×103/mL blood) was similar to that of our previous study [28].
Analysis of human BM engraftment in mice
For BM analysis, red cells were lyzed from flushed BM cell solutions with Becton Dickinson Lysing Solution following the manufacturer's instructions. Nucleated marrow cells were then analyzed by cytometry using the following monoclonal antibodies: anti-hCD45, anti-mCD45, anti-hCD33, anti-hCD41, anti-mCD41, anti-hCD34, anti-hCD11b, anti-hCD235, and anti-hCD19 (all from Pharmingen BD, except x-CD34, from Immunotech).
Statistical analysis
GraphPad InStat version 3.00 (La Jolla, CA) was used for statistical analyses. Significant differences between mouse groups were determined using the Mann–Whitney test and analysis of variance, for dual- and multiple-group comparison, respectively. Paired Student's t-test was used to compare the impact of mild hyperthermia on the ex vivo expansion of UCB CD34+ cells. P-values smaller than 0.05 were considered significant.
Results
Expansion of UCB CD34+ cells at 39°C accelerates the release of platelets in vivo
Recently, we demonstrated that UCB cells expanded at 39°C maintained short- and long-term engraftment in nonobese diabetic/severe combined immunodeficient (NOD-SCID) IL-2 receptor-gamma null (NSG) mice [20]. However, the impact of mild hyperthermia on the in vivo properties of hematopoietic cells has not yet been properly addressed. Hence, we first compared the thrombopoietic and engraftment potentials of UCB CD34+-enriched cells expanded at 39°C to those grown at 37°C.
The total progeny of 1.25×105 UCB CD34+ cells expanded for 6 days at 37°C or 39°C with the cytokine cocktail OMPC (SCF 10 ng/mL, TPO 30 ng/mL, and FL 11 ng/mL) was transplanted into sublethally irradiated NSG mice. Each mouse group (n=19) received similar doses of total nucleated cells (TNC, mean of 2.5 vs. 2.8×106, P>0.05) and CD34+ cells (1.2 and 0.8×106 at 37°C and 39°C, respectively). However, a greater number of Mk-committed cells (CD41a+) were injected in the 39°C mouse group (9.4×105 vs. 4.8×105, P<0.02), since mild hyperthermia increased the expansion of Mks over the 37°C controls, achieving a level of ex vivo expansion in line with that previously reported [26,28].
hPLTs, as measured by cytometry in PRP prepared from peripheral blood using species-specific antibodies against GPIIIa (CD41a), were found to be increased in mice transplanted with E UCB cells over saline controls (Fig. 1A). A very small proportion (<0.1%) of platelet events was positive for both human and murine antibodies during the PRP acquisitions (Fig. 1A). These events were attributed to human murine platelet dimers based on 2 characteristics: (1) compared to individual platelets, these were larger in size (forward scatter) and showed a slight increase in the granular content (side scatter), and (2) the level of murine CD41 and human CD41 cell surface expression was similar to that of corresponding single platelets of either origin (Fig. 1A).
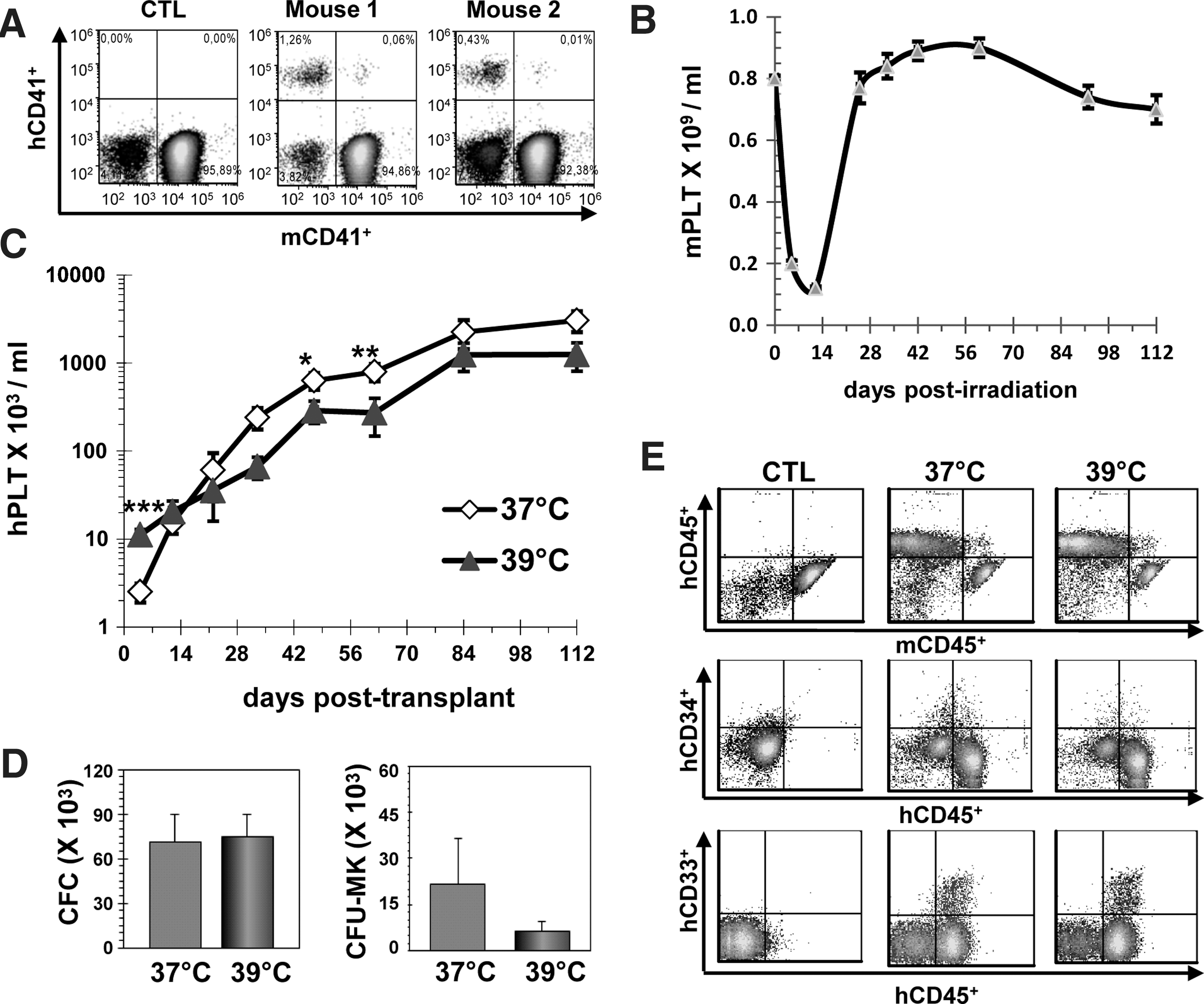
Mild hyperthermia accelerates the release of platelets in vivo.
Thrombocytopenia (<0.5×109 platelets/mL) after sublethal irradiation (325cGy) of NSG mice was clearly evident 5 and 12 days postirradiation (Fig. 1B). Murine platelet counts subsequently returned to near normal levels by day 25 (Fig. 1B). hPLTs were below the detection level on the first day post-transplant in both mouse cohorts, but were readily detected 4 days post-transplant in all recipients injected with UCB grown at 39°C (mean of 11±1.7×103/mL vs. 0.4±0.2×103/mL for control, P<0.001). Conversely, the mean level of hPLTs in mice transplanted with UCB cells expanded at 37°C (2.5±0.6×103/mL) was not statistically different to that seen in control mice 4 days post-transplant. Hence, hPLT levels in the 39°C recipients were 27.8-fold and 4.4-fold greater than those in the saline controls and the 37°C ex vivo expansion group (P<0.001), respectively. After the third week of transplantation, the opposite scenario was observed with hPLT levels usually greater in mice transplanted with the UCB cells expanded at 37°C (Fig. 1C).
Next, we investigated the impact of mild hyperthermia on short-term marrow reconstitution. A total of 6 mice per group were sacrificed 12 days post-transplant. The proportions of human CD45+ [6%±3% vs. 7%±3% with 39°C and 37°C expansion groups, respectively (mean±standard error of the mean, SEM)], CD41+ (1%±0.3% vs. 1%±0.6%), CD33+ (7%±3% vs. 7%±2%), and CD34+ (1%±0.2% vs. 1%±0.2%) cells were undistinguishable in both cohorts. Likewise, there was no statistical difference in the net cellular content (total BM cells and human subpopulations) of the marrow (data not shown) as well as the total amounts of human myeloid clonogenic (CFC) progenitors (Fig. 1D). The net content of Mk progenitors (CFU-MK) was also similar (P=0.84) in both cohorts though 2 outliers (a high and a low in the 37°C and a 39°C groups, respectively) skewed the average in favor of the 37°C cohort (Fig. 1D). Thus, ex vivo expansion at 39°C had no deleterious effect on other progenitor cell populations.
The contribution of the E cells to the long-term hematopoiesis was determined 16 weeks post-transplant (8 mice/cohort). Representative flow cytometry analyses of BM cells from sacrificed recipients are presented in Fig. 1E. The mean proportions of human CD45+ (60%±2.0% vs. 62%±7.0%) and CD34+ cells (4%±1% vs. 2%±1%) were again similar in the 37°C and 39°C mouse recipients (P>0.05). Moreover, no significant differences were observed in the frequency and net number of myeloid cells (CD33+, CD11b+, and CD235+), progenitors, and CD19+-B cells (data not shown). Therefore, mild hyperthermia accelerated the release of hPLTs in vivo without causing any harmful effect on both the short- and long-term reconstitution activity of the E UCB cells.
Ex vivo culture of UCB and mPB CD34+ cells significantly affects the hPLT production kinetics in vivo
We then determined whether ex vivo expansion under mild hyperthermia exerted a favorable thrombopoietic effect over fresh NE UCB CD34+ cells. Importantly, hPLTs were readily detected during the first week after transplant in mice injected with E UCB cells, but not in those transplanted with NE CD34+ cells (Fig. 2A, day 4 and 7). In addition, hPLT levels were higher between 11 and 42 days post-transplant in the NE group (Fig. 2A). However, hPLT levels in the mild hyperthermia group reached that in the NE group after 6 weeks post-transplant. The same procedure was performed using adult-derived mPB CD34+ cells rather than UCB cells. Because the growth response and progenitor content of mPB CD34+ cells are both lower to that of their UCB counterpart [1], we raised the starting cell dose (+20%) and the length of expansion (7 days) for mPB CD34+ cells. The net dose of cells injected in NE and E cohorts was of 1.5 and 32×105 TNC per mouse, respectively. As shown in Fig. 2B, the kinetic of hPLT recovery between E and NE mPB cells was similar to that seen previously with UCB cells (Fig. 2A). Platelet engraftment 5 days post-transplantation was superior (P<0.01) in the E group, while we found a trend for greater hPLT levels after day 14 in the NE mPB group (Fig. 2B).

hPLT recovery is significantly affected by culture.
Cotransplantation of NE with E UCB cells improves the recovery of hPLTs
To confirm these results and to directly test whether the simultaneous administration of both cell sources could result in improved hPLT recovery, we compared the hPLT kinetics in cohorts injected with either NE, E or, NE+E UCB CD34+cells (cotransplantation). The doses of TNC, CD34+ cells, CD41+ cells, CFC, and CFU-MK injected into each cohort are presented in Table 1.
Net number of cells and progenitors present in each dose (mean±SEM of 4 independent experiments) and produced from the expansion of 1.25×105 UCB CD34+ cells in culture for 6 days with the OMPC at 39°C.
Data presented for reference only (cells were not placed in culture).
ND, not determined; UCB, umbilical cord blood; Mk, megakaryocyte; NE, nonexpanded; OMPC, optimized megakaryocyte progenitor cocktail; CFC, colony-forming cell; CFU, colony-forming-unit; TNC, total nucleated cells; SEM, standard error of the mean.
The advantages of both cell sources were essentially maintained in the cotransplantation setting. Hence, hPLT levels at day 5 (Fig. 3A) were significantly greater (P<0.001) in mice cotransplanted with NE+E cells (16±3.1 hPLT×103/mL, mean±SEM) or E cells (18±4.1 hPLT×103/mL) than in mice injected with NE cells (0.5±0.4 hPLT×103/mL). Cotransplantation offered no advantage over the E group alone at 5 days after transplantation. The improved platelet recovery at 5 days seen in mice injected with E cells is consistent with the considerable increase in the total number of Mk-committed cells and Mk progenitors injected in E and NE+E cohorts compared to NE mice (Table 1).

Cotransplantation of NE with E cells improves the recovery of hPLTs in immunodeficient mice.
Similarly, the increase in hPLTs previously observed with NE cells past 10 days was recapitulated in cotransplanted mice, with higher hPLT levels at day 12 in mice that received NE+E cells or NE over E cells only (Fig. 3B, P<0.001). Interestingly, hPLT levels demonstrated a trend in favor of cotransplanted mice (1585±512.0×103/mL) with near 2-fold greater numbers at day 12 than those measured in the NE cohort (895±247×103/mL, Fig. 3B, P=0.0699). This level is 170% greater than the sum of each individual cell source, suggesting that both contributors synergized together to promote hPLT recovery.
The long-term hPLT kinetics in the 3 test groups are shown in Fig. 3C. In general, the highest levels of hPLTs were observed in the NE+E or NE cohorts, whereas the lowest levels were seen in mice transplanted exclusively with E cells. Taken together, these results demonstrate that cotransplantation of NE cells with E cells is advantageous for platelet recovery due to their complementary and synergistic thrombopoietic activities.
Impact of expansion and cotransplantation on short- and long-term BM engraftment
Next, we sought to investigate whether engraftment properties were different between E and NE UCB cells, also if cotransplantation had a beneficial impact on BM engraftment characteristics. In all, 10 to 12 recipients were sacrificed 12 days post-transplantation to measure the level of human BM engraftment and progenitor content. The proportions of hCD45+ cells at 12 days were similar in all 3 cohorts (Table 2). However, the total number of nucleated cells and human CD45+ cells tended to be lower in E mice compared to the NE or NE+E cohorts (Table 3), despite a 20-fold and 14-fold increase in the dose of human TNC and CFC injected in the E recipients versus NE transplants (Table 1).
Cells positive for CD235 or hCD45.
Number of mice analyzed from 3 independent experiments.
P<0.05 versus E.
P<0.01 versus E.
m; murine, h; human; E, expanded; NE, nonexpanded.
Cells positive for CD235 or hCD45.
Number of mice analyzed from 3 independent experiments.
P<0.05 versus E.
Moreover, there was a significant loss of erythroid differentiation potential associated with the E cells, since the proportion and net numbers of human CD235a+ (glycophorin A) cells were significantly reduced in the E cohort compared to the other groups (Tables 2 and 3). Because erythroid cells do not express the pan-hematopoietic antigen CD45, when human BM engraftment was considered to be the sum of both CD235a+ and CD45+ cells, then short-term engraftment was significantly greater (P<0.05) in mice transplanted with NE+E and NE cells (Table 3). Interestingly, the number of hCD11b+ cells (expressed on neutrophils and monocytes) increased by almost 20% in cotransplanted mice versus NE mice (Table 3), though the difference failed to be statistically significant. This finding is consistent with the accelerated neutrophil recovery reported in several clinical trials [12,14,16]. No significant difference in the proportion or the number of human CD33+, CD41+, and CD34+ was observed (Tables 2 and 3).
When the NE and NE+E cohorts were compared one to another, a clear tendency of increased cellularity (TNC, hCD45+, hCD34+, hCD11b+, hCD33+, and hCD235+) was evident in the cotransplanted cohort (Table 3). In addition, cotransplantation resulted in greater numbers of human hematopoietic progenitors in the BM, with Mk progenitor numbers increased by 2.4-fold and 13.2-fold in NE+E versus NE, and E recipients, respectively (Fig. 4A). In addition, CFC progenitors in cotransplanted mice were also greater than those seen in E (9.5-fold, P<0.05) and NE (1.6-fold, P>0.05) mice (Fig. 4A). Together, these results suggest that cotransplantation of NE with E cells improves short-term BM reconstitution after nonlethal whole-body irradiation. This improvement appears to be the combined results of the increased cell dose and of the coinjection of 2 functionally distinct HSPC sources with overlapping and complementary short-term reconstitution potential [29].

Impact of cotransplantation on human BM engraftment. Total number of myeloid and Mk progenitors detected 12 days
The frequency and net number of human and murine subpopulations analyzed 4 months post-transplant in the 3 cohorts are presented in Table 4 and Fig. 4B, respectively. Human BM engraftment (Fig. 4B) and progenitor content (Fig. 4C) were generally comparable between the different groups. The frequency of hCD45+ cells in cotransplanted mice was reduced compared to NE mice (Table 4), which was mostly attributable to a reduction in the human CD19+ B-cell compartment in these mice. Indeed, the total number of CD19+ cells was significantly lower in NE+E (19±9.0×106) versus NE (30±7.0×106, P<0.05) cohorts. A reduction in CD19+ cells was also observed in the E group (23±7.0×106). It should be stressed that the reduction in CD19+ cells in cotransplanted mice was contributed mostly by 1 of the 3 independent cohorts analyzed. When the cohorts from this experiment were excluded, then the mean numbers of CD19+ cells in the NE+E and NE mice were similar [26±4.0×106 (n=16) versus 32±3.0×106 (n=12), respectively (P=0.31)], whereas that of the E group remained lower [18±3.0×106 (n=11), P<0.05 vs. NE].
Cells positive for CD235 or hCD45.
Number of mice analyzed from 3 independent experiments.
P<0.01 versus E.
P<0.05 versus E.
P<0,05 versus NE.
P<0.001 versus E.
E cells committed to the Mk lineage are responsible for the improved short-term platelet recovery
Next, we sought to determine the cellular origin of the early hPLT engraftment from E UCB cells. Since the cytokine cocktail OMPC used herein was optimized for Mk progenitor expansion [20] and based on previous studies done with TPO culture [19,24], Mk progenitors and Mks present in the graft after expansion were most likely responsible for the release of hPLTs at or before day 12 post-transplant. To test this hypothesis, expanded, cells were fractionated by immunomagnetic selection of CD41+ cells into 2 subpopulations: the CD41+-enriched fraction (mean of 92% CD41+, n=2) was administered to the first group (mean dose of 0.8×106 total cells/mouse) and the CD41−-impoverished fraction (<10% CD41+) to the second group (mean dose of 1.7×106 total cells/mouse). Despite the administration of a lower cell number in the CD41+ group, hPLT levels were significantly superior in mice transplanted with the CD41-enriched cells 5 days post-transplant, whereas hPLT levels in mice injected with CD41-depleted cells failed to be superior to the background level (Fig. 5A). Conversely, hPLT levels became and remained significantly superior (P≤0.05) by day 12 and thereafter in the CD41−-depleted cohort (Fig. 5B, E). Thus, E cells committed to the Mk lineage provide the first wave of hPLT production, whereas noncommitted CD41− E cells provide the second thrombopoietic wave.
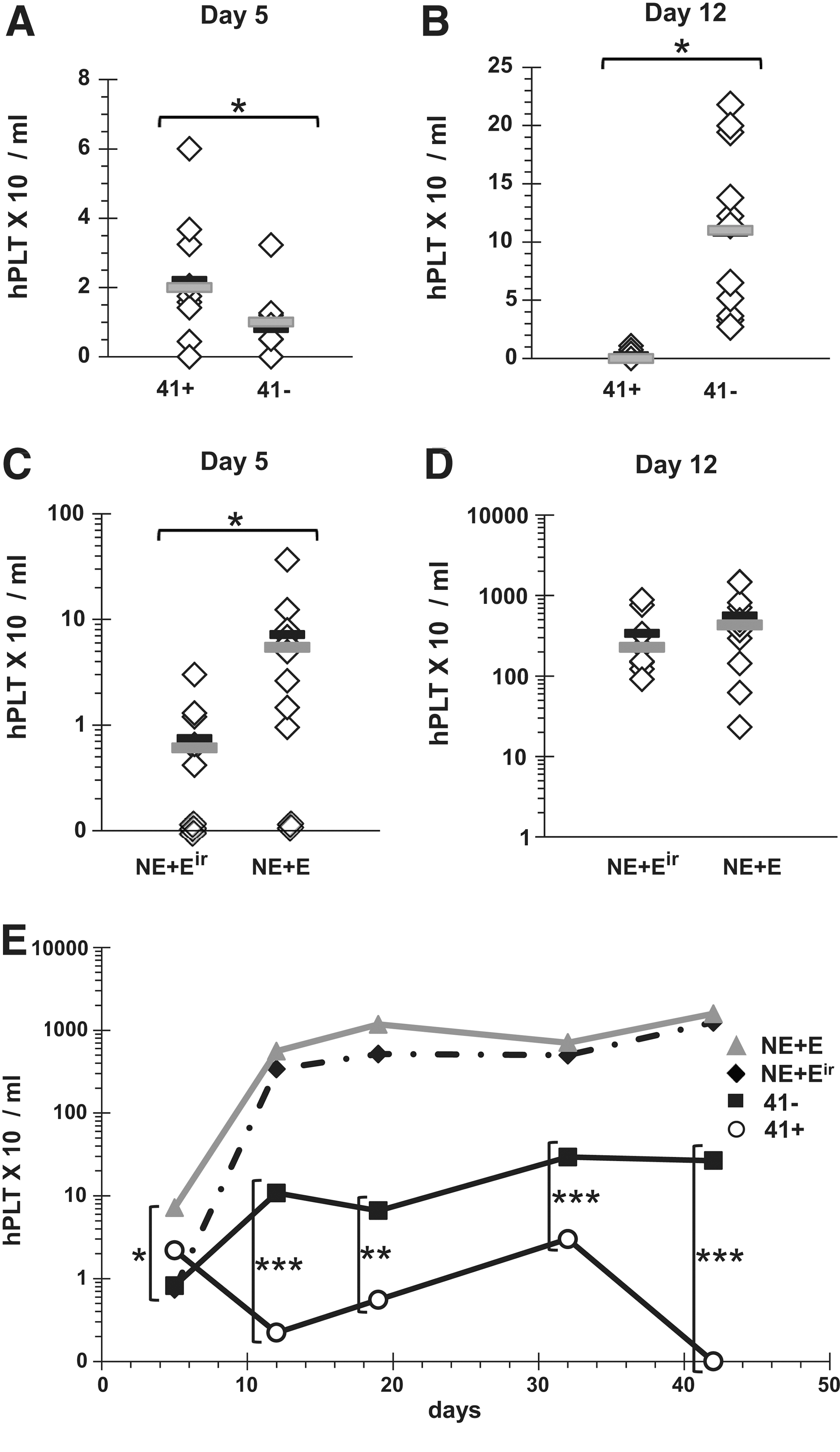
UCB cells committed to the Mk lineage are responsible for the improved short-term platelet recovery.
The results showing improved short-term hPLT recovery in cotransplanted mice (Fig. 3) may have resulted from the expanded fraction, facilitating the engraftment of NE cells (so-called accessory cell effect [30]). To determine whether the E cells played such a role, E cells were irradiated and administered concomitantly with NE cells (NE+Eir), since accessory cells maintain their activity after irradiation, and compared to NE+E cells. Interestingly, hPLT recovery 5 days post-transplant was abolished with irradiation, indicating that the improved hPLT recovery associated with cotransplantation relied on E cell engraftment (Fig. 5C, P<0.05). No significant differences were observed at day 12 (Fig. 5D) or thereafter (Fig. 5E).
Discussion
Transplantation of UCB HSPCs results in a longer median time to platelet (29 vs. 60 days) and neutrophil (20 and 27 days) engraftment than adult derived HSPCs [31]. This may be due to the combination of lower cell numbers [32] and different intrinsic molecular and cellular properties related to their developmental stage in ontogeny [33]. Advances in UCB transplantation practice such as double UCB transplantation and in culture protocols for the ex vivo expansions of HSPCs have already improved engraftment characteristics [1,16]. Nevertheless, platelet reconstitution remains a significant issue for these patients. Cell dose is paramount for successful engraftment, and the number of cells available per UCB unit is limited; it is not surprising that only mitigated results could be achieved when a single UCB unit was fractionated, expanded, and cotransplanted with the second unmanipulated fraction in the first series of clinical trials [10]. However, the odds that cellular therapy could overcome delayed hematopoietic recovery received a much-needed lift when studies such as that by Weinreb and Barker demonstrated the safety of transplanting multiple UCB units [34 –36]. As stated by Barker and colleagues, this provided the opportunity to completely expand one unit while maintaining the second unit intact. Recent clinical results from 2 independent groups certainly support the efficacy of such a strategy [16,17].
Building on these principles, we have focused our research efforts on improving platelet recovery, which is a significant challenge given that very few studies have reported any benefit on this important biological response. Herein, we demonstrate that UCB cells primed along the Mk lineage by a 6-day expansion procedure are capable and responsible for early hPLT release in vivo. Moreover, we demonstrate that NE and E UCB cells have complementary and at times synergistic thrombopoietic activities. The results obtained in this work confirm and extend those previously reported by Tijssen et al. [23]. In their work, they showed that expansion of mPB CD34+ cells with TPO and IL-1β for 7 days reduced the time for platelet release in vivo over that of unexpanded cells. Thus, these 2 studies demonstrate that cotransplantation of NE with cells expanded in a condition supporting Mk expansion can improve hPLT recovery. It is also important to stress that the thrombopoietic activities of the E and NE cells reported herein were highly underestimated, since the half-life of hPLTs in immunodeficient mice is considerably less than that seen in humans [37,38].
The culture protocol used herein was composed principally of 2 key elements: first, the use of the cytokine cocktail OMPC optimized by statistical design of experiments for the expansion of Mk progenitors while maintaining the expansion of CD34+ cells and myeloid progenitors [20], and second, the incubation of the UCB CD34+ cultures under mild hyperthermia. The latter was selected based on 3 considerations: (1) its proexpansion [26] and (2) prodifferentiation effects on Mks (increased differentiation and accelerated maturation [39]) and (3) acceleration of platelet release from the E cells. However, incubation under mild hyperthermia did reduce the thrombopoietic potential of the E cells during the second month post-transplant. Nonetheless, mild hyperthermia conditions were retained, because the gain in early hPLT recovery exceeded the lost seen later on, given that the contribution of the NE cells during that period exceeded that of the E fraction (37°C or 39°C).
The benefit of our proposed approach remains untested in human settings. It is possible that the growth condition proposed herein such as mild hyperthermia and low concentration of cytokines may undermine the expansion of both short- and long-term multilineage, reconstituting cells, though our results suggest otherwise except for a reduction in the frequency and the number of B cells at 4 months. While this reduction was largely attributable to a single-animal cohort, the reduction in CD19+ cells was also observed in the E group as well as in our previous study [20]. Hence, it may originate from the cytokine cocktail (OMPC) used to expand the UCB cells. However, it is unlikely to represent an issue in humans in the context of double UCB transplantation given that long-term chimerism is rarely observed and that uncultured units commonly take over [16,40]. Indeed, the unmanipulated unit has a significant engraftment advantage, given that the culture affects homing and other intrinsic properties of HSCs [41]. Moreover, immunological-mediated clearance of the E cells typically tips the balance in favor of the NE unit [40]. In any events, the principal objective of the E cells in the context of double UCB transplantation is to provide short-term platelet and neutrophil support and not long-term hematopoiesis. The increased number of hPLTs, human BM cells, and progenitors during the first 2 weeks post-transplant in cotransplanted mice certainly supports this strategy.
In conclusion, this study demonstrates for the first time in the context of UCB transplantation that hPLT recovery and short-term BM engraftment in nonlethal whole-body irradiated mice can be improved by the cotransplantation of E and NE cells. It will be most important to test the strategy proposed herein in the human setting to determine its full clinical impact.
Footnotes
Acknowledgments
We wish to thank the staff at the Hôpital St-Francois d'Assise and at the Laval University animal facility for their precious help in collecting UCB units and for their technical contributions, respectively. The authors would also like to thank Nellie Dumont for her contribution to this work. This work was supported by a National Blood Foundation grant and by the TheCell Network. D.C.R. is supported by the FRQS.
Author Disclosure Statement
The authors have no conflict of interest to disclose.